代谢组学法研究转录因子SlNAC4对番茄果实代谢产物的影响
2019-06-11郑海英张冬青赵晓丹
郑海英,张冬青,赵晓丹*
(北京工商大学食品学院,北京 100048)
番茄作为消费者喜爱且营养丰富的果蔬,是肉质果实研究的模式材料。转录因子NAC(NAM、ATAF1/2、CUC2)的变化影响番茄果实色素和硬度等品质指标[1]。运用代谢组学研究转录因子NAC对番茄代谢产物的影响,可以丰富番茄相关研究的理论内容,也对改善番茄品质具有重要意义。
在植物中,转录因子NAC显著特点是有一个高度保守的N端NAC结构域,有5 个保守的子域(A~E),还有一个不同的转录调节C端能启动和抑制多种靶基因的转录[2-3]。研究结果显示番茄中一共有104 个NAC基因[4],分布在12 条染色体上,其中NAC4位于第I条染色体上,编码蛋白长413 aa,分子质量46.93 kDa[5]。Riahi等[6]测序50 份野生葡萄和73 个葡萄品种发现NAC4转录基因编码区缬氨酸变成异亮氨酸,功能性候选基因在物种进化过程中为适应环境而发生基因变异,说明转录因子在植物发育和胁迫调控等生命活动中发挥着关键作用。据研究,硬皮豆在不同盐度、寒冷、干旱和脱水胁迫条件下,NAC4基因表达均有一定的调节,尤其是盐胁迫导致NAC4转录水平增加了6 倍,NAC4基因通过载体克隆表达,在大肠杆菌的细胞转化后,NAC4蛋白增强了大肠杆菌细胞耐盐胁迫能力,表明NAC4基因可适合作为提高植物对盐胁迫和其他非生物胁迫耐受性的遗传候选基因[7]。落花生中Mu转录因子NAC4能调控植株抗旱功能,MuNAC4基因的过表达,在干旱环境下能减少对细胞膜的损害、增强膜的渗透性和抗氧化酶的活性[8]。植株抗胁迫能力的强弱势必会影响果实成熟过程中的代谢水平,转录因子NAC4是影响果实成熟的重要因素。沉默NAC4基因,果实软化相关基因EXP/LOX/PG/PL的表达下调,显著抑制果实软化酶活性,延缓果实软化进程,导致果实呼吸速率降低,乙烯含量下降,延缓叶绿素降解和类胡萝卜素积累的速度,果实无法正常变红[9]。Shan Wei等[10]研究转录因子NAC在香蕉成熟过程中的表达情况,发现果皮和果肉中NAC4转录水平降低,表明NAC4可能通过与乙烯信号物质的相互作用参与香蕉果实的成熟。有研究显示NAC4调控植物对氮的运输功能,氮是蛋白质、叶绿素、核酸、酶、生物激素等重要成分,转录因子NAC4主要是通过作用中柱鞘细胞中AFB3基因,影响应答硝酸盐的侧根基因密度[11],并增强植物的耐盐能力[12]。植株生理上的变化势必影响果实的成熟基质,相关研究也表明转录因子NAC4确实在植株生长和果实成熟过程中具有影响基体代谢水平的重要作用。
本实验以正常番茄和SlNAC4沉默的番茄为研究对象,采用代谢组学法全面分析两组番茄在破色期的代谢产物分布情况。代谢组学法是对植物中的化合物分布进行全面探索的一种有效方法,能够找出差异代谢产物[13],进而挖掘可能发生变化的代谢通路[14],掌握植物生长发育中的多种生理信息[15],对果实的品质[16]和营养改良提供初步的理论支撑[17]。能阐明生物合成的自然多样性,有助于对化合物进行注释和分类,发现新的生物合成分支[18]。番茄含有丰富的抗氧化和酚类物质,比如VC、酚类化合物(芦丁和绿原酸)和类胡萝卜素等[19-20]。运用代谢组学阐明基因对番茄成熟过程中代谢体系的调控作用,对番茄营养、品质和风味改良具有重要意义[21-22]。
1 材料与方法
1.1 材料与试剂
野生型和基因沉默番茄(Solanum lycopersicum)由中国农业大学食品生物技术实验室提供,野生型品种为AC(Ailsa Craig)。
甲醇、乙醇(均为分析纯) 北京化工厂;乙腈(色谱纯) 美国Fisher Chemical公司;甲酸、氨水(色谱纯) 美国ACS公司;所有分离用有机溶剂均为国产分析纯。
1.2 仪器与设备
Triple TOFTM5600高效液相色谱-质谱(high performance liquid chromatography-high resolution mass spectrometer,HPLC-HRMS)联用仪(配有DBS和MMDF Trigger IDA数据采集系统及MarkerView、PeakView、MetabolitePilot数据分析软件) 美国AB Sciex公司;超低温冷冻储存箱 青岛海尔特种电器有限公司;Hamburg离心机 德国艾本德公司;电子天平德国Sartorius公司。
1.3 方法
1.3.1 色谱条件
Waters An ACQUITY UPLC BEH Amide(100 mm×2.1 mm,1.7 μm)色谱柱;柱温35 ℃;流动相:A为25 mmol/L醋酸铵+25 mmol/L氨水,B为异丙醇-乙腈(9∶1,V/V)+0.1%甲酸;进样量4 μL;流速0.4 mL/min;洗脱梯度:0~0.5 min,95% B;0.5~9.5 min,95%~65% B;9.5~10.5 min,65%~40% B;10.5~12 min,40% B;12~12.2 min,40%~95% B;12.2~15 min,95% B。
1.3.2 质谱条件
采用HPLC-HRMS联用仪检测柱上洗脱的代谢产物。正负两种模式,喷雾电压为5 000 V和-4 500 V;气帘气压力为30 psi,雾化气压力(Gas 1)为60 psi,辅助气压力(Gas 2)为60 psi;离子源温度为650 ℃;质谱数据采集模式为IDA模式,TOF质量数范围为60~1 200 Da。采集时间为150 ms/spectrum,采集速率为100 counts/s,总循环时间为0.56 s;40 GHz四极杆/质量分析器多通道TDC检测,脉冲为11 kHz;动态排除设置为4 s;在采集过程中,每20 个样品对质量精度进行校准。此外,为评价HPLC-HRMS在整个采集过程中的稳定性,在样品前中后分别插入质量控制(quality control,QC)样品以作实验技术重复评估。
1.3.3 样品制备
野生型(6 个平行样品)和沉默组番茄(5 个平行样品)都在破色期后3 d取样。果实采摘后,取番茄果肉部位,液氮冻存后,-80 ℃保存待用。提取时取出样品在冰上解冻,用120 μL预冷的50%甲醇溶液提取20 mg研磨成粉的番茄样品,旋转1 min,室温放置10 min,提取液在-20 ℃过夜,4 000×g离心20 min后,上清液转入96 孔板。样品在-80 ℃保存,然后进行HPLC-HRMS分析。此外,取等量的11 份样品提取液混合而成QC样品,在样品前中后分别插入以检验实验的重复性。
1.4 数据处理
1.4.1 代谢产物鉴定
利用生物信息分析进行质谱数据解读,主要利用XCMS软件进行物质检测,利用metaX软件对检测到的物质进行定量、差异物质筛选。利用代谢物一级m/z和二级碎片离子m/z与数据库里标准品匹配进行代谢物鉴定。首先利用开源软件metaX通过物质的一级m/z分别与PlantCyc、KEGG、HMDB数据库里的代谢物进行匹配,得到一级鉴定结果;同时利用in-house图谱库对物质二级质谱图进行代谢物鉴定。最后利用公共PlantCyc数据库和KEGG数据库对物质进行功能注释。
1.4.2 代谢产物定量分析
代谢物的定量信息来自于物质一级色谱峰面积。本研究利用XCMS软件对每个物质在每个样品的强度信息提取,随后对提取出的数据使用metaX软件进行质控,首先去低质量峰(将QC样本中缺失超过50%,或者实际样本中缺失超过80%的离子去除),随后利用KNN(K-Nearest Neighbors)方法进行缺失值填充,接着采用PQN(Probabilistic Quotient Normalization)和Q C-RSC(QC-robust spline batch correction)方法校正。对校正后的数据进行过滤,即将所有QC样品中变异系数(coefficient of variation,CV)大于30%的离子过滤(CV大于30%的离子在实验过程中波动较大,不进行差异定量分析)。
1.4.3 差异代谢产物的筛选
利用开源软件metaX对代谢组学结果分析,进行单变量和多变量分析,从而获得组间差异代谢物。其方法包括参数检验和非参数检验、差异表达倍数分析、主成分分析(principal component analysis,PCA)、偏最小二乘法判别分析等。同时满足3 个条件即认定为差异化合物:1)物质倍数比值ratio≥2或者ratio≤1/2;2)wicoxon统计检验BH校正值q-value≤0.05;3)偏最小二乘法模型的变量投影重要度值VIP≥1。
1.4.4 代谢通路图的绘制
找出差异代谢产物的KEGG号,进入KEGG首页(https://www.kegg.jp/),进入分析工具KEGG Mapper,进入代谢通路图工具Search Pathway,将搜索选为番茄属sly,将差异代谢化合物KEGG号输入,数据库就会出现与化合物相关的代谢通路图,结合各条通图路绘制出代谢网络。
2 结果与分析
2.1 代谢产物的代谢组学分析结果
在所有检测到的代谢产物中,既能与数据库物质的一级m/z匹配到,也能与数据库的碎片离子(二级)m/z匹配到的物质数目正模式有1 092 个,负模式有510 个。与正常番茄相比,SlNAC4基因沉默番茄中含量增加的差异化合物有640 种,含量减少的化合物有880 种。在这些差异物质中,只有67 种其一级和二级质谱均能够在数据库中找到匹配物质,从而得到鉴定,其余大部分尚无法得到最终鉴定,因此后面重点研究的差异代谢产物主要集中在这67 种得到鉴定的物质上。
2.2 SlNAC4沉默组番茄与普通番茄的差异代谢产物分析
基于HPLC-HRMS数据对沉默组番茄和对照组番茄进行PCA,发现两组番茄呈现显著差异[23-24],见图1。两主成分分数体现了77.41%总变异,对照图1中两组番茄的相对位置发现,两者在PC1轴上相距甚远,且PC1对该差异的贡献率为71.61%,说明两种番茄的代谢化合物差异很大。

图1 基于HPLC-HRMS一级峰面积数据破色期的代谢差异PCAFig. 1 PCA plot for metabolite differences of CRC4 compared to wild-type tomato at the breaking stage based on HPLC-HRMS peak area data
对最终鉴定得到的物质进行统计学分析后,两组番茄中可以鉴定的具有显著差异的化合物一共有67 种,分布情况如表1所示。

表1 1 SlNAC4lNAC4沉默组番茄与对照组番茄的差异代谢化合物分布Table 1 Differential metabolites in CRC4 compared to wild-type tomato

续表1
差异较大的氨基酸衍生物及二肽类19 种,其中脯氨酰-络氨酸、异亮氨酰-天冬氨酸相对含量增加极显著(q-value<0.001),N-乙酰-L-苯丙氨酸、络氨酰-亮氨酸、脯氨酰-苏氨酸、N-乙酰-DL-亮氨酸、L-焦谷氨酸相对含量下降极显著(q-value<0.001)。氨基酸和多肽的变化是由于器官特异性活动,将叶子的氨基酸转移到果实中[25],相比于对照组,基因沉默组果实的氨基酸发生了变化,说明转录因子SlNAC4可能对控制从其他器官转移氨基酸至果实的生物过程起着重要的调控作用。
差异较大的维生素类7 种,其中VB1、4-吡哆酸、D-泛酸钙相对含量增加显著(q-value<0.01),VL1、VC相对含量降低显著。
差异较大的糖类及其衍生物4 种,其中D-木酮糖增加极显著。
差异较大的核酸及其衍生物11 种,其中6-苄氨基嘌呤、1-甲基鸟苷相对含量增加极显著(q-value<0.001),胞嘧啶、二磷酸腺苷、3’-磷酸腺苷-5’-磷酸硫酸酯下降极显著(q-value<0.001)。
差异较大的生物碱4 种,其中罂粟碱相对含量增加极显著(q-value<0.001),乙酰胆碱、苯甲酰芽子碱相对含量下降极显著(q-value<0.001)。
差异较大的香精香料4 种,其中羟基丙酮相对含量增加极显著(q-value<0.001),3-甲硫基丙醇相对含量增加显著(q-value<0.01),香豆素相对含量降低显著(q-value<0.01)。
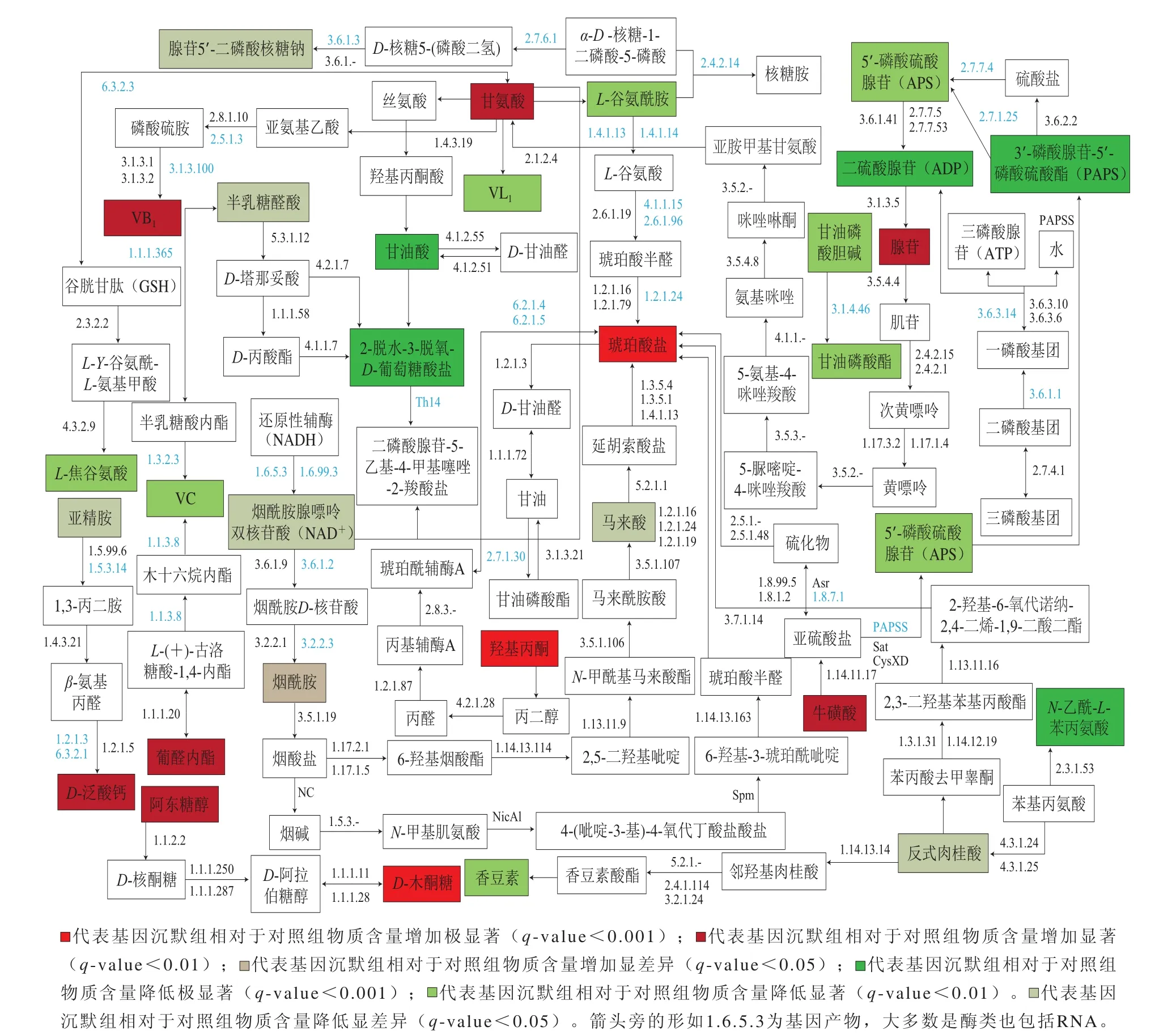
图2 2 SlNAC4lNAC4调控破色期番茄初级代谢通路图Fig. 2 Modulation of primary metabolism by SlNAC4lNAC4 in breaking tomato fruit
差异较大的有机酸9 种,其中3-丁炔酸相对含量增加极显著(q-value<0.001),邻羟基苯乙酸、丙炔酸相对含量增加显著(q-value<0.01),间羟基苯甲酸、水杨酸、甘油酸相对含量下降极显著(q-value<0.001),2-苯基丁酸相对含量降低显著(q-value<0.01)。果实由未成熟到成熟阶段有机酸、糖类、氨基酸及多肽等代谢强盛,也是果实从涩到甜的生理过程,期间呈味物质发生变化,沉默组相对于对照组有机酸、糖类、氨基酸及多肽含量的变化差异很大,说明转录因子SlNAC4影响番茄的呈味[26-27]。
2.3 差异代谢产物在代谢通路中的分布分析
为阐明SlNAC4沉默番茄受到影响的代谢途径,使用KEGG进行详细的代谢途径查找与分析[28],在大量的番茄途径中,25 条重要的代谢途经中只有31 种化合物被识别出来,基于此研究和总结,综合作出总代谢通路见图2。
代谢通路网络图的绘制,可以更清楚地找到各代谢化合物的合成及分解去向,为解析转录因子对番茄化合物的代谢影响提供一条便利途径。从图2可以看出,嘌呤代谢途径中,腺苷5’-二磷酸核糖钠、L-谷氨酰胺含量减少,甘氨酸含量显著增加。甘氨酸在代谢途径中向L-谷氨酰胺的方向分解,L-谷氨酰胺含量减少,有利于甘氨酸积累。甘氨酸一共有4 条分解代谢和3 条合成代谢,从总体来说SlNAC4基因的沉默有利于甘氨酸的积累。3’-磷酸腺苷-5’-磷酰硫酸含量减少极显著,它可以从两条途径分解成5’-磷酸硫酸腺苷(含量减少显著),接着沿着一条通路分解为二磷酸腺苷(含量减少极显著)。苯基苯氨酸的代谢途径中,N-乙酰-L-苯丙氨酸、反式肉桂酸含量降低,琥珀酸盐则显著升高。烟酸盐和烟酰胺代谢途径中,烟酰胺和琥珀酸盐含量增大、烟酰胺腺嘌呤双核苷酸和马来酸含量则减少。硫磺代谢途径中,牛磺酸、琥珀酸盐含量升高、二磷酸腺苷和3’-磷酸腺苷-5’-磷酰硫酸含量降低。碳代谢途径中,甘氨酸含量升高,一条去向L-谷氨酰胺,一条去向甘油酸,L-谷氨酰胺和甘油酸含量都降低,原因嘌呤代谢途径已经解析过。戊糖和葡糖醛酸互换途径中,阿东糖醇和D-木酮糖含量升高很大。VB1的代谢途径中,甘氨酸和VB1含量均增加。VC代谢途径中,葡醛内酯含量降低,VC含量也降低,葡醛内酯可以向VC转化。苯丙醇的合成途径中,香豆素和反式肉桂酸含量都降低。氨基酸的合成途径中,L-谷氨酰胺和VL1含量都减少。这些代谢产物的互相影响关系,还需要后续结合番茄基因序列数据进行综合分析[29-30]。
3 结论与讨论
番茄是肉质果实相关研究的重要模式作物,在成熟过程中具有典型的生理变化过程。NAC是植物中独有的转录调节因子,因其具有一个高度保守的N端NAC结构域而被赋予极高的研究价值,NAC4属于NAC基因家族成员之一。至今转录因子NAC4对果实营养物质的影响鲜有报道,因此基于代谢组学研究转录因子SlNAC4对番茄代谢产物的影响具有巨大的意义。
番茄中很多营养物质含量发生明显变化,比如氨基酸类甘氨酸含量显著升高、L-谷氨酰胺含量显著降低,多种二肽含量发生明显变化;维生素类中VB1、D-泛酸钙显著增加,烟酰胺含量增加,VC、VL1含量显著降低;糖类物质阿东糖醇和D-木酮糖含量都增大幅增加;核酸类物质含量基本上降低,但腺苷含量显著增加;另外还有一些呈香呈味物质类,羟基丙酮等含量增加极显著,反式肉桂酸、香豆素含量降低,马来酸、甘油酸、水杨酸等有机酸含量发生大幅变化。以上结果均能说明转录因子SlNAC4确实影响了番茄的营养物质代谢水平。代谢通路图的制作概括了其中31 种化合物的代谢关系,对下一步结合基因测序分析提供了重要线索,为解析转录因子对番茄的代谢影响提供一条便利的途径。
综上所述,转录因子SlNAC4对番茄的氨基酸、维生素、糖、有机酸、核酸及核苷酸类代谢有重要影响。其具体的影响机理还有待继续深入研究。
