基质及多菌灵对水分胁迫下芳樟扦插苗根系活力的影响
2018-06-25张国防黄秋良毕学琴谢亚兵陈瑞炎陈志平
林 铃 张国防 黄秋良 毕学琴 谢亚兵 陈瑞炎 陈志平
(1. 福建农林大学林学院,福建 福州 350000;2. 永安市林业局,福建 三明 366000)
芳樟(Cinnamomumcamphoravar.linaloolifera)是樟树(C.camphora)的生化变种[1],因其富含芳樟醇而极受国际市场欢迎。张国防等[2-4]对它的遗传、变异和精油成分等领域展开了许多研究,但目前尚未有基质、多菌灵、水分胁迫对芳樟复合影响的研究。芳樟苗木规模化生产易受气候环境的影响,而长江中下游地区为亚热带季风气候,气温高且暴雨多发,容易对芳樟苗产生水分胁迫影响。根系是受水分胁迫影响最直接的器官,同时根系也是苗木从基质中吸收矿质元素的重要器官,长时间的水分胁迫最终会影响芳樟苗的正常生长甚至死亡,导致苗木产量和质量下降。
本试验对芳樟苗水分胁迫以及育苗相关技术进行研究,比较芳樟苗根系矿质元素吸收能力,分析不同基质配比和多菌灵对水分胁迫下芳樟根系活力的影响,以期为芳樟优质苗木的培育提供理论基础。
1 材料和方法
1.1 试验材料
容器苗试验的材料为芳樟1年生扦插苗,生长均匀,高度相近,苗木运抵大棚后缓苗1个月。容器育苗袋材质为可降解的无纺布 (6 cm × 9 cm)。泥炭土生产自福建省三明市尤溪县,有机质体积分数 ≥ 65%,腐殖酸体积分数为20%,pH值为4.0~5.5。保水透气性良好的粘性黄心土取自福建农林大学后山B层,放在阳光下充分消毒、晒干,并磨碎过1 mm筛。珍珠岩规格大小为3~6 mm,且颗粒饱满。土壤消毒剂采用多菌灵可湿性粉剂 (有效质量含量:50%多菌灵 + 3%中生菌素)。
1.2 试验方法
1.2.1试验设计
试验地位于福建省福州市福建农林大学南门妙峰山苗圃地施育苗大棚 (118°08′~120°31′ E,25°15′~26°29′ N),采用三元二次回归正交旋转设计 (见表1),总共设置23个处理 (T1~T23),每个处理30株,重复3次。育苗基质设计根据泥炭土体积分数设置5种配比方案 (见表2)。多菌灵用水溶解并均匀的喷洒在育苗基质上,使其充分混合在基质之中。水分胁迫0 cm组用塑料薄膜铺于地面与土壤隔离,将苗木置于其上,每周浇1次水;2.0 cm组、5 cm组、8.0 cm组、10 cm组分别置于有塑料薄膜覆盖的土沟中,每天及时补充各组水分,保证试验所需水分高度。
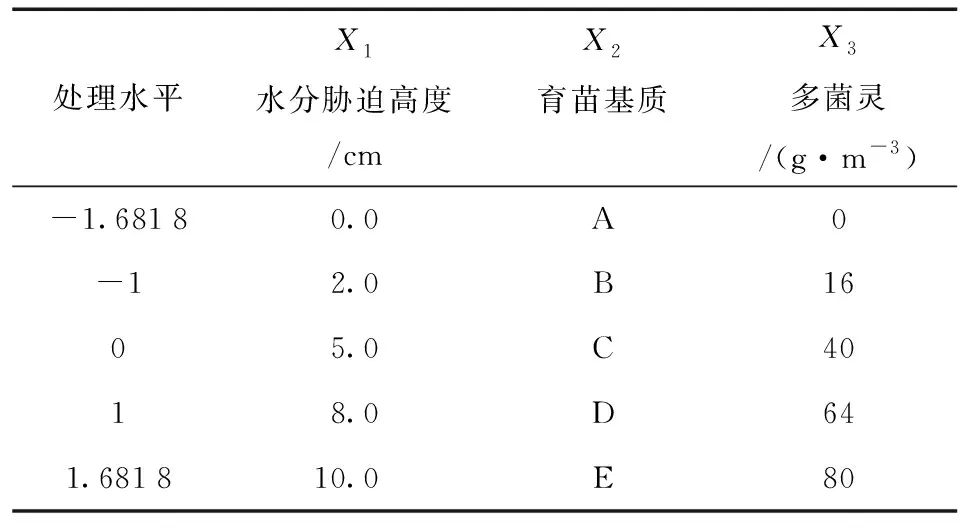
表1 不同因素各水平实施方案Table 1 Implementary plan of different factors and levels
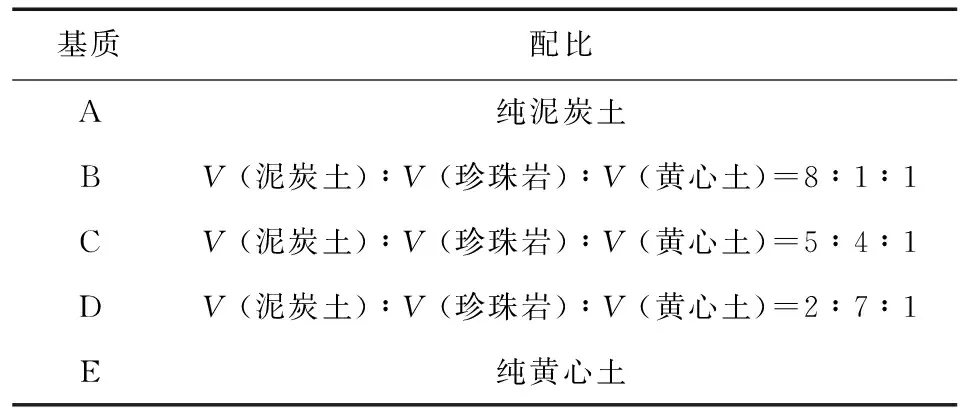
表2 基质配比方案Table 2 Substrates ratio scheme
1.2.2测定方法
试验于2017年4月1号开始,于6月1号对各处理每个重复的每1株芳樟苗木根系样品进行采集,根系的矿物质质量分数采用火焰原子吸收光谱法[5]测定,根系活力采用TTC法[6]测定。
1.3 分析方法
通过DPS软件对芳樟根系活力数据进行分析得到数学模拟方程,如公式 (1)。
Y=a+bX1+cX2+dX3+eX12+fX22+
gX32+hX1X2+iX1X3+jX2X3
(1)
式中:a、b、c、d、e、f、g、h、i、j均为常数。
在公式 (1) 的基础上,将X2、X3的值设为0,得到X1单因素与芳樟根系活力方程,如公式 (2)。以此类推得到X2单因素与芳樟根系活力方程以及X3单因素与芳樟根系活力方程,如公式 (3)~(4)。
Y=a+bX1+eX12
(2)
Y=a+cX2+fX22
(3)
Y=a+dX3+gX32
(4)
在公式 (1) 的基础上,如果X2与X3存在显著的互作效应,则将X1的值设为0,得到X2X3互作效应与芳樟根系活力方程,如公式 (5)。并以此类推得到X1X3互作效应与芳樟根系活力方程以及X1X2互作效应与芳樟根系活力方程,如公式 (6)~(7)。
Y=a+cX2+dX3+fX22+gX32+jX2X3
(5)
Y=a+bX1+dX3+eX12+gX32+iX1X3
(6)
Y=a+bX1+cX2+eX12+fX22+hX1X2
(7)
试验数据采用Excel 2003和DPS 7.05进行分析。
2 结果与分析
2.1 不同处理对芳樟根系矿物质质量分数的影响
由表3可知,芳樟根系矿质元素K、Ca、Na、Mg、Fe、Zn、Cu质量分数最高的处理组分别为T5、T6、T5、T21、T21、T5、T5,分别比平均值高26.99%、25.64%、26.38%、8.14%、43.05%、92.52%、48.20%。芳樟根系矿质元素K、Ca、Na、Mg、Fe、Zn、Cu质量分数最低的处理组分别为T4、T3、T12、T3、T8、T12、T3,分别比平均值低28.29%、40.21%、38.45%、11.94%、64.26%、75.74%、62.00%。可见,不同处理对芳樟矿质元素质量分数影响较大的是Fe、Zn、Cu,其次是矿质元素K、Ca、Na,影响最小的是矿质元素Mg。
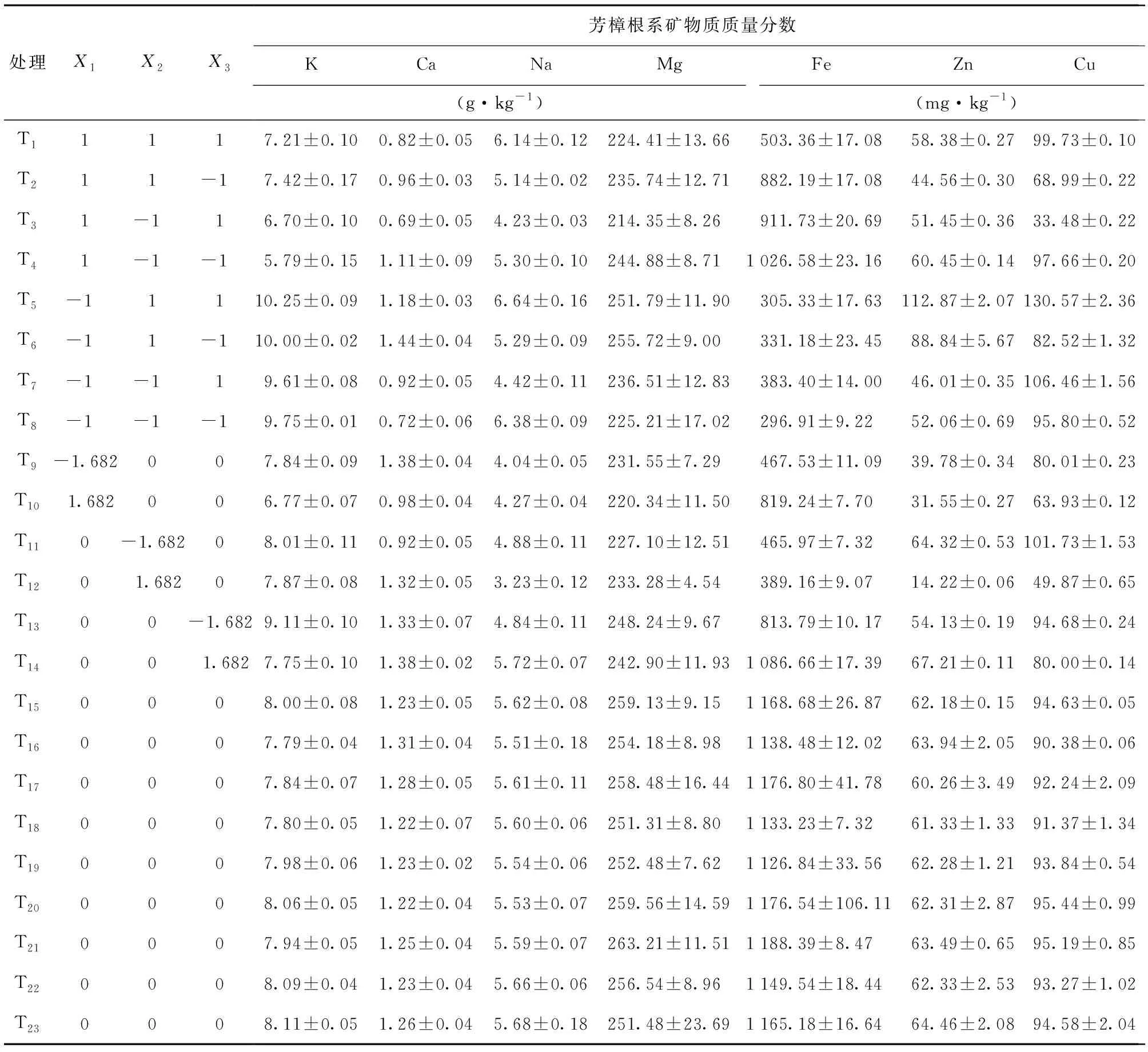
表3 芳樟根系矿质元素质量分数Table 3 Mass fraction of mineral elements in roots of C.camphora var. linaloolifera
由表4可知,各矿质元素质量分数的处理间均方都远大于处理内均方,说明芳樟根系各矿质元素质量分数不同处理间数据差异较大;处理内差异较小,反映了处理内个体样本之间数据的稳定性。不同水平基质、多菌灵、水分胁迫的复合处理对芳樟根系K、Ca、Na、Mg、Fe、Zn、Cu质量分数都产生了极显著影响 (P< 0.01)。不同处理对芳樟根系各矿质元素质量分数差异影响的效果为Cu > K > Fe > Zn > Na > Ca > Mg。
2.2 不同处理对芳樟根系活力的影响
由图1可知,不同的处理对芳樟根系活力产生比较大的影响。其中T6、T7、T10的芳樟根系活力处于较低的水平;T3、T4、T8、T11、T12、T13、T14均处于较高的水平;其余处理组相较而言处于中等水平。芳樟处理组整体根系活力均值为20.52 mg/(g·h);T4根系活力最大,为30.72 mg/(g·h);高于均值49.70%;T10号处理组根系活力最小,为10.55 mg/(g·h);低于均值48.58%。说明不同组合的处理对芳樟根系活力产生了较大的差异,水分胁迫制约了芳樟根系活力的提升,而基质与多菌灵有利于芳樟抵抗水分胁迫。
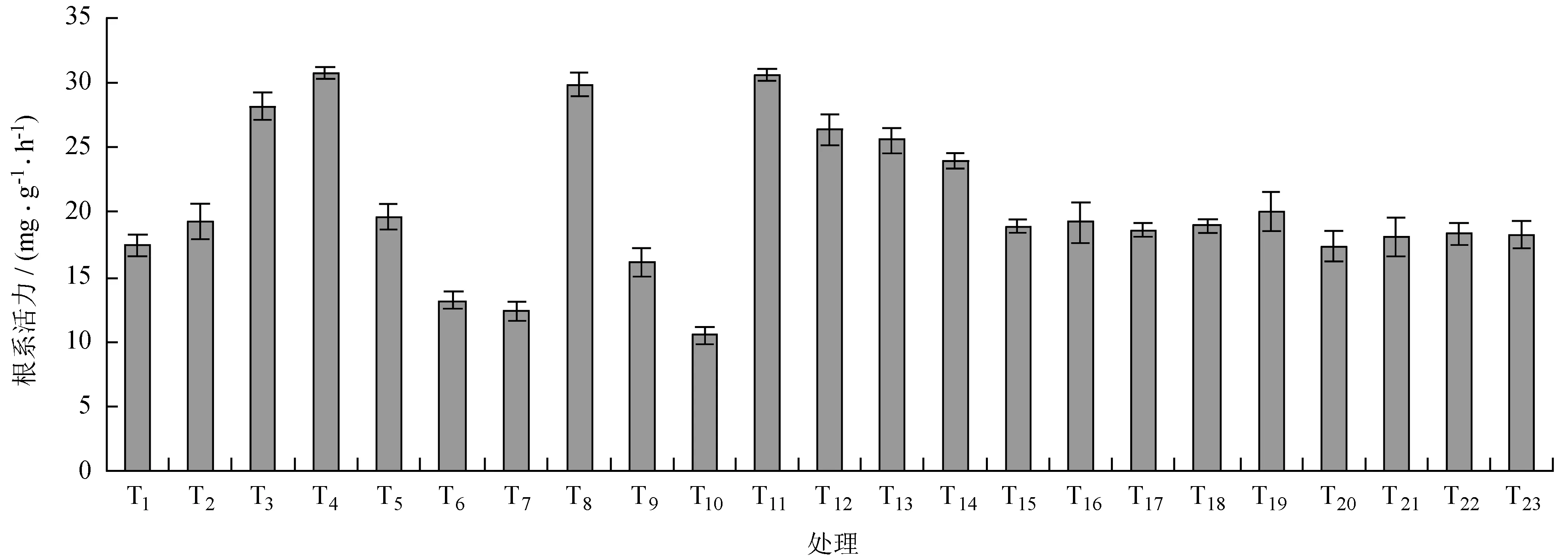
图1不同处理对芳樟根系活力的影响
Fig.1 Effects of different treatments on root activity ofC.camphoravar.linaloolifera
2.3 芳樟根系活力数学方程模型的建立及回归显著性分析
将根系活力的数据导入DPS软件得到如下芳樟根系活力数学方程模型。
Y=18.664+0.812X1-2.831X2-1.322X3-2.090X12+3.260X22+1.960X32-1.584X1X2+0.830X1X3+3.090X2X3
(8)
对芳樟根系活力回归方程进行方差分析 (表5) 可知,其F回归=4.721 >F0.01(9,13)=4.19,说明芳樟根系活力对于X1、X2、X3的回归方程建立成立。基质中泥炭土体积分数、水分胁迫高度的平方水平、基质中泥炭土体积分数的平方水平、单位体积基质中多菌灵使用量的平方水平以及基质中泥炭土体积分数和单位体积基质中多菌灵施用量的相互作用对芳樟的根系活力均表现出显著水平 (P< 0.05)。
2.4 单因素分析
3个单因素芳樟根系活力方程如下:
Y=18.664+0.812X1-2.090X12
(9)
Y=18.664-2.831X2+3.260X22
(10)
Y=18.664-1.322X3+1.960X32
(11)
从图2可知,水分胁迫高度对芳樟的根系活力产生抑制作用,而且干旱胁迫的抑制程度明显要高于水涝胁迫。育苗基质中泥炭土含量对芳樟根系活力有促进作用;当泥炭土体积分数低于50%时,对芳樟根系活力促进作用就会减弱。多菌灵也对芳樟根系活力有促进作用,但在本试验中不使用多菌灵的情况下对芳樟根系活力最有利。X3与芳樟根系活力效应曲线是开口向上的抛物线,增大单位体积基质中的多菌灵使用量是否能放大其促进作用,有待进一步的研究。
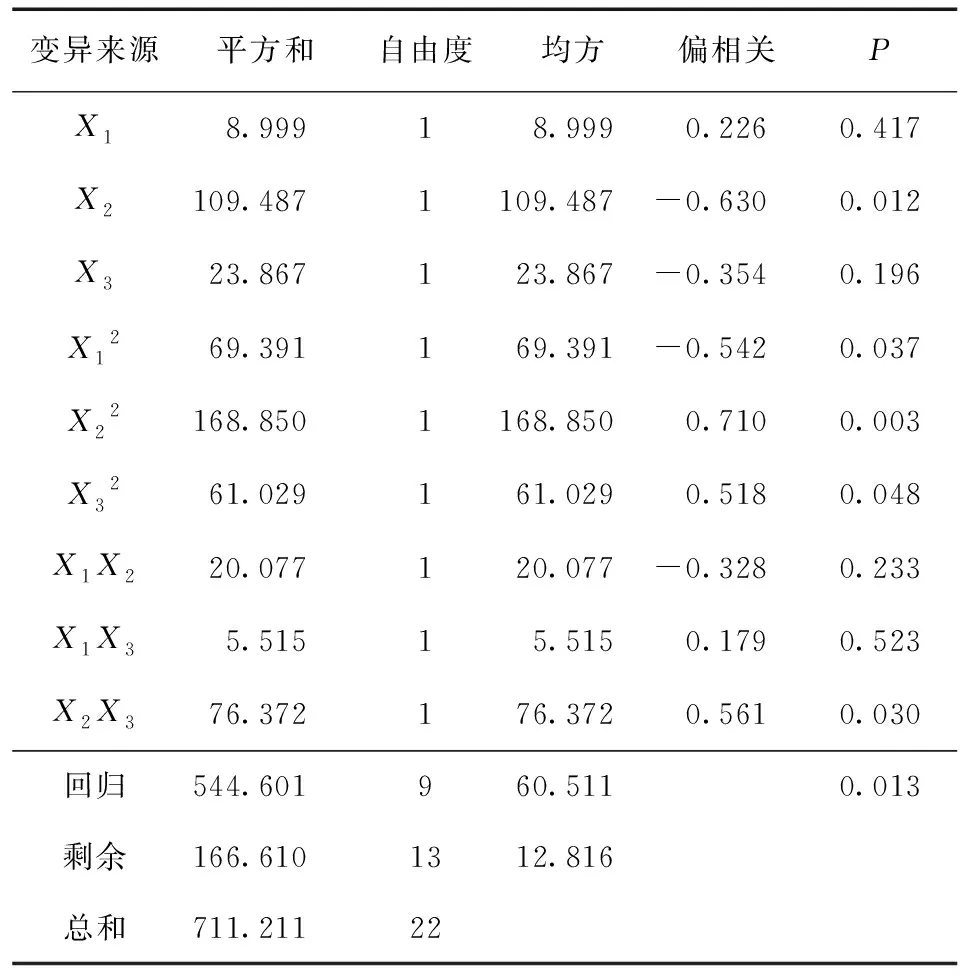
表5 水分胁迫高度、基质中泥炭土体积分数、多菌灵与芳樟根系活力回归方程的方差分析Table 5 Variance analysis results for effects of different factors of X1, X2 and X3 on root activity of C.camphora var. linaloolifera
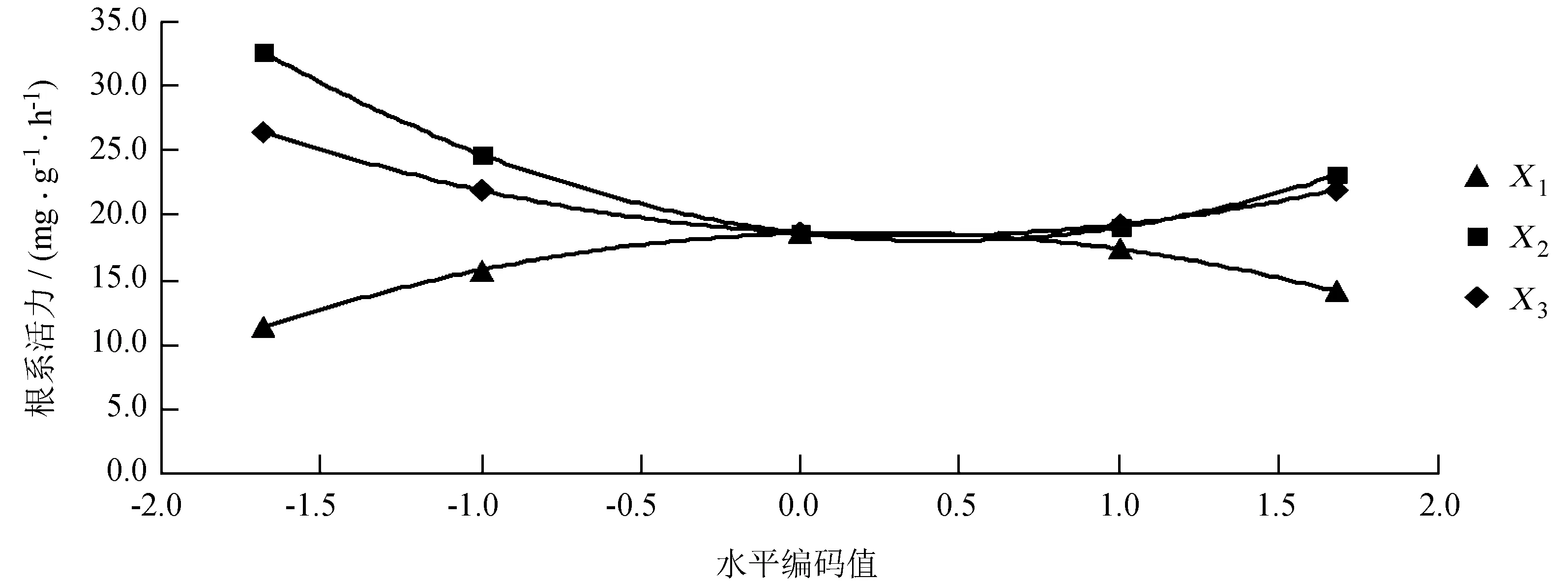
图2三个单因素与芳樟根系活力的效应曲线
Fig.2 Root activity effect curve of 3 single factors ofC.camphoravar.linaloolifera
在本试验范围内,X1、X2和X3的水平编码值分别为0、-1.682、-1.682,即X1=5 cm、X2=100%、X3=0 g/m3时,芳樟根系活力达到最大,为49.16 mg/(g·h)。
2.5 互作效应分析
由表5可知,只有X2与X3存在显著的互作效应。故X2、X3互作效应芳樟根系活力方程如下:
Y=18.664-2.831X2-1.322X3+3.260X22+
1.960X32+3.090X2X3
(12)
从图3可知,当育苗基质中泥炭土体积分数大于80%时,增加多菌灵的使用浓度会降低芳樟的根系活力;当育苗基质中泥炭土体积分数为50%左右时,增加多菌灵的使用浓度不会芳樟的根系活力产生太大影响;育苗基质中泥炭土体积分数低于20%时,增加多菌灵的使用浓度会促进芳樟的根系活力。所以X1、X2的水平编码值分别为1.682、-1.682,即X2=100%、X3=0 g/m3时,芳樟根系活力达到最大,符合单因素的分析结果。
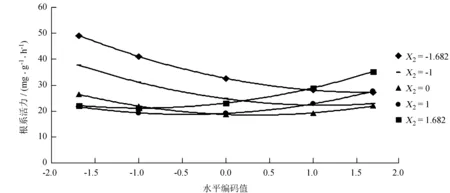
图3不同基质中泥炭土含量(X2)下多菌灵使用量(X3)与芳樟根系活力的关系
Fig.3 Relationship amongX2,X3and root activity ofC.camphoravar.linaloolifera
3 结论与讨论
无论是干旱胁迫还是水淹胁迫都会抑制芳樟苗根系活力,而且干旱胁迫的抑制程度会强于水淹胁迫。许国伟等[7]对水稻 (Oryzasativa) 在水分胁迫下全生育期的根系活力进行了研究,结果发现轻度的水分胁迫可以明显提高水稻的根系活力。牛素贞等[8]对1年生野生茶树 (Camelliasinensis) 幼苗展开了干旱胁迫,幼苗的根系活力随着干旱胁迫的加强而下降。秦嗣军等[9]对东北山樱 (Cerasussachalinensis) 幼苗进行了干旱、水淹胁迫,两种水分胁迫最终都会导致幼苗根系活力下降,但是水淹胁迫下短期内会提高幼苗根系活力。杨传杰等[10]研究也发现,水分过多或过少都会降低棉花 (Gossypiumherbaceum) 的根系活力。
育苗基质中泥炭含量介于20%~50%,芳樟幼苗根系活力表现最差;纯泥炭土基质和纯黄心土都有利于芳樟苗的根系活力,但是纯泥炭土基质明显要优于纯黄心土。符真珠等[11]研究了多年生金叶复叶槭 (Acernegundo) 嫩枝扦插基质的配比,对于金叶复叶槭幼苗根系活力影响中,m(珍珠岩)∶m(泥炭土)=1∶1时在2个月的试验期内表现最好,明显优于纯河沙和纯珍珠岩。熊燕等[12]高山杜鹃 (Alpinerhododendro) 幼苗根系活力进行了相关研究,当m(松针)∶m(草炭)∶m(沼渣)=2∶1∶1时最有利于幼苗的根系活力提高。
使用多菌灵可以提高芳樟苗根系活力,而且其使用量因基质的不同也有所不同。当基质中泥炭土体积分数 > 80%时,增大单位体积基质中多菌灵的使用量会抑制芳樟苗根系活力;当基质中的泥炭土体积分数处于50%左右时,不同的多菌灵使用量并不会对芳樟苗根系活力大小产生明显差异;但是增加多菌灵在单位体积黄心土基质中的使用量会明显提高芳樟苗根系活力。王丽丽等[13]研究了多种杀菌剂对马拉巴粟 (Pachiramacrocarpa) 残根使用效果,发现多菌灵提高根系活力的效果最好。赵国玲等[14]研究了3种杀菌剂对平邑甜茶 (Minushupehensis) 幼苗根系活力的影响,发现多菌灵混土后提高幼苗根系活力最好。多菌灵对芳樟苗根系活力的效应曲线为开口向上的抛物线,介于本试验单位体积基质中多菌灵使用量设置范围有限,但增大其使用量能否继续放大这种促进作用,有待进一步研究。
根系是植物吸收基质土壤中矿质元素的重要器官,本试验芳樟幼苗对于K、Ca、Na、Mg、Fe、Zn、Cu的吸收也产生了极显著差异 (P< 0.01),而且这种吸收差异与根系活力大小存在较强的关联性。刘媛等[15]在镉胁迫对于秋华柳 (Sulixvariegata) 扦插苗根系活力以及根系Ca、Mg、Mn、Zn、Fe含量进行了研究,发现幼苗根系活力随着镉胁迫加强而降低,而且对根系5种矿质元素含量产生了显著抑制作用 (P< 0.05)。而且植物在矿质元素缺少的情况下,会反馈给根系,导致植物根系活力下降。肖家欣等[16]对枳 (Poncirustrifoliata) 在Zn、Fe缺乏下的根系活力进行研究,发现Zn、Fe缺乏下会导致枳根系活力显著下降。
综上所述,使用水分胁迫、基质配比以及多菌灵会对芳樟幼苗根系活力产生明显差异,影响大小关系为X2>X1>X3,其中水分胁迫制约了芳樟根系活力提升,而基质与多菌灵有利于芳樟抵抗水分胁迫。芳樟苗根系对于矿质元素K、Ca、Na、Mg、Fe、Zn、Cu的吸收也会随着根系活力的影响而显现出极显著差异 (P< 0.01)。而基质和多菌灵的合理配比施用能提高芳樟苗根系活力,促进根系对于矿质元素吸收。基质中泥炭土含量对芳樟苗根系活力影响较大,而多菌灵的施用量也要随着基质不同而作出相应调整。
[1] Fujita Y.CinnamomumcamphoraSieb. and its allied species. their interrelations considered from the view-points of species characteristics, chemical constituents, geographical distributions and evolution[J]. Shokubutsugaku Zasshi, 1952, 65(771): 245-250.
[2] 张国防, 陈存及. 福建樟树叶油的化学成分及其含量分析[J]. 植物资源与环境学报, 2006, 15(4): 69-70.
[3] 张国防, 陈存及, 赵刚. 樟树叶油地理变异的研究[J]. 植物资源与环境学报, 2006, 15(1): 22-25.
[4] 张国防, 陈存及, 邢建宏, 等. 芳樟工业原料林营建中的若干问题[J]. 林业科技开发, 2004, 18(3): 7-10.
[5] 工广印, 荆瑞俊, 张伟娟, 等. 火焰原子吸收光谱法测定大葱中的10种金属元素[J]. 光谱实验室, 2009, 26(2): 246-250.
[6] Knievel D P. Procedure for estimating ratio of live to dead root dry matter in root core samples[J]. Crop Sci, 1973, 13: 124-126.
[7] 徐国伟, 孙会忠, 陆大克, 等. 不同水氮条件下水稻根系超微结构及根系活力差异[J]. 植物营养与肥料学报, 2017, 23(3): 811-820.
[8] 牛素贞, 宋勤飞, 樊卫国, 等. 干旱胁迫对喀斯特地区野生茶树幼苗生理特性及根系生长的影响[J]. 生态学报, 2017, 37(21): 7333-7341.
[9] 秦嗣军, 吕德国, 李志霞, 等. 水分胁迫对东北山樱幼苗呼吸等生理代谢的影响[J]. 中国农业科学, 2011, 44(1): 201-209.
[10] 杨传杰, 罗毅, 孙林, 等. 水分胁迫对覆膜滴灌棉花根系活力和叶片生理的影响[J]. 干旱区研究, 2012, 29(5): 802-810.
[11] 符真珠, 杜君, 孟月娥, 等. 不同基质和设施条件对金叶复叶槭嫩枝扦插繁殖的影响[J]. 中国农学通报, 2015, 31(31): 25-29.
[12] 熊燕, 曲彦, 唐焕伟, 等. 低温胁迫及不同基质对高山杜鹃生理和根系活力的影响[J]. 东北农业大学学报, 2013, 44(10): 117-121.
[13] 王丽丽, 文婧, 黄燕雪, 等. 不同杀菌剂与杀菌时间对发财树残根生长的影响[J]. 华南农业大学学报, 2014, 35(5): 69-75.
[14] 赵国玲, 姜兴印, 孙燕, 等. 三种杀菌剂对平邑甜茶生理指标的影响[J]. 果树学报, 2013, 30(5): 823-828.
[15] 刘媛, 马文超, 张雯, 等. 镉胁迫对秋华柳根系活力及其Ca、Mg、Mn、Zn、Fe积累的影响[J]. 应用生态学报, 2016, 27(4): 1109-1115.
[16] 肖家欣, 齐笑笑, 张绍铃. 锌和铁缺乏对枳生理指标、矿物质含量及叶片超微结构的影响[J]. 应用生态学报, 2010, 21(8): 1974-1980.
