豫南黑猪及其杂交后代猪H-FABP基因的遗传变异研究
2016-06-22郭吉利韩雪蕾李素雅乔瑞敏任广志李新建河南农业大学牧医工程学院郑州45000河南省新大牧业有限公司郑州450000
郭吉利,韩雪蕾,李素雅,吕 刚,乔瑞敏,任广志,李新建(.河南农业大学牧医工程学院,郑州 45000;.河南省新大牧业有限公司,郑州 450000)
豫南黑猪及其杂交后代猪H-FABP基因的遗传变异研究
郭吉利1,韩雪蕾1,李素雅2,吕 刚1,乔瑞敏1,任广志1,李新建*
(1.河南农业大学牧医工程学院,郑州 450002;2.河南省新大牧业有限公司,郑州 450000)
摘 要:本研究以60头豫南黑猪、30头巴×豫猪、27头杜×豫猪、40头豫×苏猪为研究对象,采用PCR-RFLP方法检测猪心脏脂肪酸结合蛋白(H-FABP)基因的5-UTR(Hinf I)和内含子2上(HaeIII、MspI和Hinf I*)的4个SNP位点在试验群体中的分布,旨在研究H-FABP基因在豫南黑猪及其杂交后代猪中的遗传变异。结果显示,5-UTR突变位点在4个试验群体中均表现中度多态(0.25<PIC<0.5),等位基因H的频率分别为0.83,0.33,0.70和0.61;除巴豫猪群体外,其余3个试验群体的基因频率和基因型频率均处于Hardy-Weinberg平衡状态(P>0.05);内含子2上的3个突变位点处于连锁平衡状态,在4个群体中的显性等位基因频率分别为0.32、0.58、0.45和0.50,除豫南黑猪外,其余试验组群体均表现为中度多态(0.25<PIC<0.5)。本研究通过对H-FABP基因的多态位点在试验群体中的遗传变异研究,进一步揭示了H-FABP基因在不同群体中的遗传规律,为下一步通过对H-FABP基因的选择加快群体的选育提供参考依据。
关键词:豫南黑猪;杂交后代猪;H-FABP基因;PCR-RFLP法
近年来,随着人们生活水平的不断提高,猪的肉质问题已经引起了生产者和消费者的重视。肌内脂肪(Intramuscular fat,IMF)含量是影响猪肉品质的一个重要指标,与猪肉的多汁性、嫩度和风味直接相关[1-4],一般认为2%~3%的IMF含量可以产生理想的口感[5],但由于近年来过度地追求高瘦肉率,导致IMF含量降至1%~1.5%[6],严重影响了肉质风味。IMF含量作为肉质性状的一个重要指标,不能在活体准确地测量,必须在猪只达到一定体重后才能通过屠宰取样,代价巨大且周期漫长。国外育种者研究发现,IMF含量具有较高的遗传力(0.6),因此,利用分子标记辅助选择是最有效的方法[7]。猪心脏脂肪酸结合蛋白(Heart Fatty Acid-binding Protein,H-FABP)基因属于FABP家族,主要在心肌、骨骼肌和乳腺中表达,其主要作用是参与胞内脂肪酸的摄取和运输,可将脂肪酸从细胞膜上运到脂肪酸氧化和甘油三酯及磷脂的合成位置[8]。猪H-FABP基因定位于6号染色体上,前人研究发现在5’-UTR 和Intron2分别存在Hinf I和HaeIII、MspI和Hinf I*4个酶切多态性位点[9]。众多研究表明,H-FABP基因的遗传变异是影响IMF含量的主要因素之一[10-15],因此被认为是影响猪IMF的重要候选基因。
豫南黑猪是以河南省优秀地方品种淮南猪为母本,美系杜洛克为父本历时10余年培育出的一个耐粗饲的瘦肉型新品种,具有繁殖性能优良、胴体瘦肉率高、肉质鲜美等优点,IMF含量高达4.11%[16,17]。本研究为了利用豫南黑猪的优良遗传特点,在杂交测定的基础上,对豫南黑猪及其杂交后代群体中H-FABP基因的Hinf I和HaeIII、MspI、Hinf I*4个酶切遗传变异进行检测与分析,为H-FABP基因的分子标记法进一步在豫南黑猪选育过程中的应用提供理论基础和参考依据。
1 材料与方法
1.1 试验材料
试验群体包括豫南黑猪60头、巴克夏×豫南黑猪杂交后代30头、豫南黑猪×苏太猪杂交后代40头、杜洛克×豫南黑猪杂交后代27头,总计157头,全部来自河南省豫南黑猪育种中心。采集耳组织样,放入含有70%酒精的EP管,采用酚仿抽提方法提取基因组DNA,-20 ℃保存备用。
1.2 试验方法
1.2.1 H-FABP基因的PCR扩增
参照文献[9]设计特异性扩增引物,如表1所示。以试验猪基因组DNA为模板进行PCR反应。PCR扩增总体积均为15 μL,其中双蒸水5.8 μL,上下游引物各0.6 μL,TaqMix 7.5 μL,DNA模板0.5 μL,瞬时离心混匀。PCR反应程序为:94 ℃预变性5 min,94 ℃变性45 s,(61 ℃,53 ℃)退火45 s,72 ℃延伸2 min,33个循环,72 ℃延伸10 min,PCR产物用1.5%琼脂糖凝胶电泳检测。
1.2.2 酶切反应
PCR产物经过琼脂糖凝胶电泳检测之后,进行酶切。总反应体系:总体积为15 μL,其中内切酶0.5 μL (12 U/μL),10×Buffer 1 μL,PCR产物5 μL,ddH2O 8.5 μL,混匀后37 ℃水浴酶切30 min,然后用2%的琼脂糖凝胶进行电泳,根据各引物片段大小电泳30~60 min,在紫外灯下观察结果并分型,并用凝胶成像分析系统拍摄成像。随后挑选不同基因型个体送交生工生物工程(北京)有限公司,以确证分型结果。

表1 SNP扩增的引物序列
1.2.3 数据处理
试验数据经Excel进行初步处理后,采用SPSS17.0对其进行统计与分析,结果采用均值±标准误的形式表示。
2 结果与分析
2.1 H-FABP基因SNP位点PCR扩增结果
H-FABP基因的5’- UTR和Intron2中的4个SNP位点PCR扩增产物用1.5%的琼脂糖凝胶电泳进行检测(见图1和2)。PCR产物检测结果显示,扩增片段与目的片段大小一致,条带清晰,可以直接进行下一步的酶切试验。
2.2 H-FABP基因SNP位点PCRRFLP分型及测序
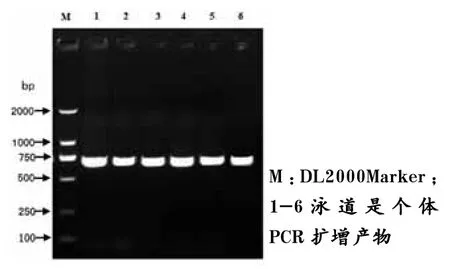
图1 H-FABP基因的693 bp扩增产物
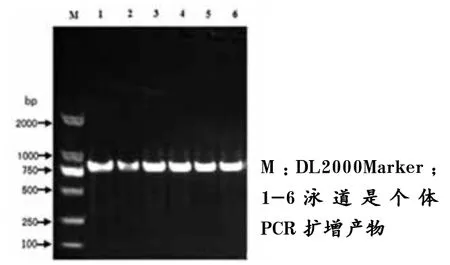
图2 H-FABP基因的816 bp扩增产物
PCR产物酶切反应后,用2%的琼脂糖凝胶进行检测,Alpha凝胶成像系统照相并进行基因分型,结果如图3~图6所示。其中Hinf I-RFLP位点扩增片段全长693 bp,存在4个Hinf I酶切位点,产生339、172、98、59和25 bp 5个片段,由于存在一个因T/C突变而产生的Hinf I酶切位点,当该位点为C时酶切位点消失,172 bp和59 bp片段合并产生231 bp的片段,从而得到3种基因型,分别定义为HH、Hh和hh基因型(见图3)。
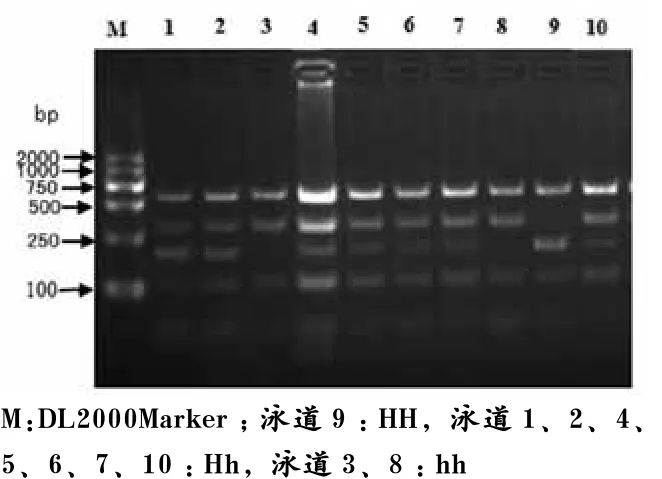
图3 猪H-FABP基因Hinf I-RFLP位点PCR产物酶切多态电泳图
如图4所示,HaeIII-RFLP位点扩增片段全长816 bp,存在3 HaeIII个酶切位点,产生405、278、117和16 bp 4个片段,其中存在一个由于G/ C突变而产生的HaeIII酶切位点,当此位点为C等位基因时,酶切位点消失,278 bp和405 bp片段合并产生683 bp的片段,从而得到3种基因型,分别定义为DD、Dd和dd基因型。
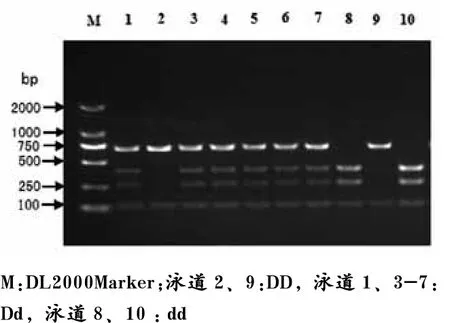
图4 猪H-FABP基因HaeIII-RFLP位点PCR产物酶切多态电泳图
如图5所示,MspI-RFLP位点扩增片段全长816 bp,在该片段750 bp处,存在一个由于T/C突变而产生的MspI酶切位点,酶切后片段大小分别为750 bp和66 bp,定义为AA基因型,当此酶切位点消失时为816 bp,定义为aa基因型。
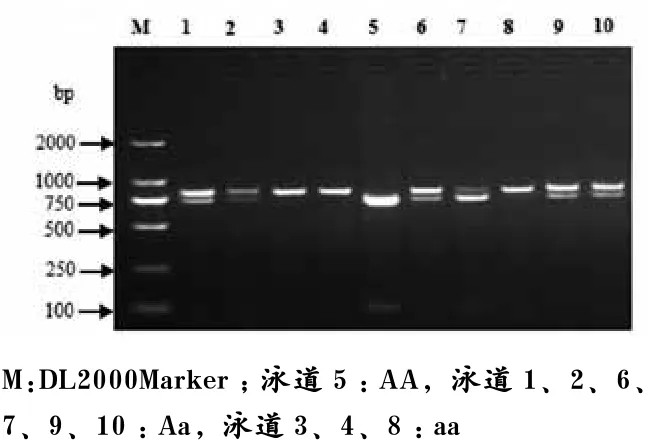
图5 猪H-FABP基因MspI-RFLP位点PCR产物酶切多态电泳图
如图6所示,Hinf I*-RFLP位点扩增片段全长816 bp,此片段中存在3个Hinf I*酶切位点,分别产生527、217、47和32 bp 4个片段,其中有一个Hinf I*酶切位点是由于在Intron2 的1 970 bp处发生C/T突变而产生,因此当此酶切位点消失时,217 bp和47 bp片段合并产生264 bp的片段,从而得到3种基因型,分别为BB型、Bb型和bb型。
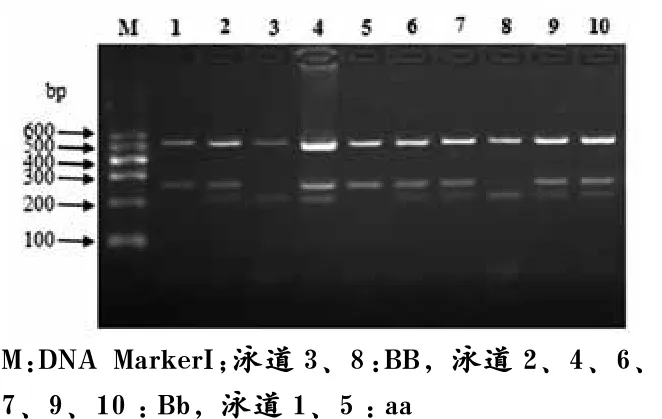
图6 猪H-FABP基因Hinf I*-RFLP位点PCR产物酶切多态电泳图
2.3 试验群体H-FABP基因SNP的遗传特征分析
对H-FABP基因的4个SNP位点在试验群体中进行基因频率和基因型频率分析,结果如表2所示, 第二内含子上的HaeIII-RFLP、MspI-RFLP和Hinf I*-RFLP3个突变位点的分型结果一致,并处于连锁平衡状态。
其中HaeIII-RFLP位点存在3种基因型分别为DD、Dd和dd,等位基因d是优势等位基因,群体中χ2值0.37(<χ=5.99),差异不显著(P>0.05);Hinf I*-RFLP位点有3种基因型HH、Hh和hh, H等位基因在群体中为优势等位基因,群体中χ2值1.33(<χ=5.99)因此差异不显著,群体处于Hardy-Weinberg平衡状态;上述2个位点的多态信息含量分别为0.35、0.37都属于中度多态(0.25<PIC<0.5),杂合度分别为1.81、1.97(表2)。

表2 H-FABP基因5’-UTR及第二内含子SNP基因型、基因频率及遗传多态指标
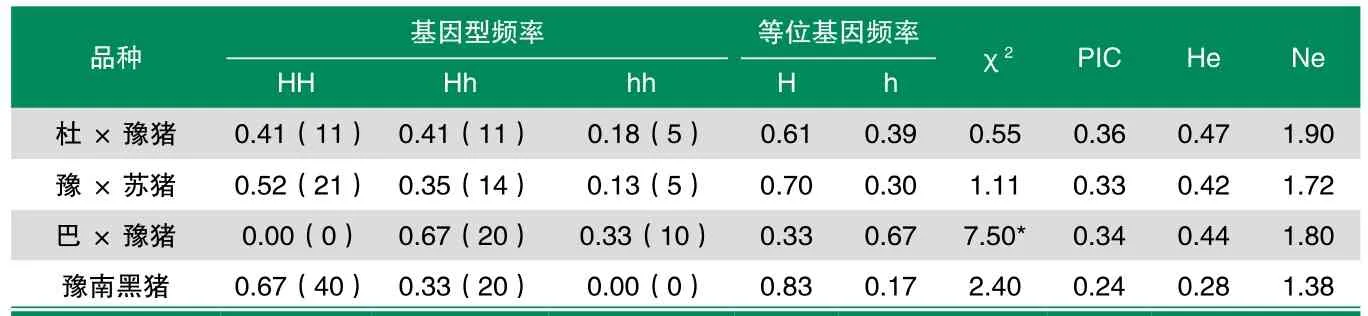
表3 Hinf I-RFLP位点在豫南黑猪及其杂交后代中基因型频率分布
如表3显示,H-FABP基因 5’-UTR上的Hinf I-RFLP位点的基因型频率在杜×豫猪、豫×苏猪和豫南黑猪中χ2值分别为0.55、1.11 和2.40(<χ=5.99),差异不显著(P>0.05),群体处于Hardy-Weinberg平衡状态;巴×豫猪χ2值为7.50(>χ=5.99),差异显著(P<0.05),群体处于Hardy-Weinberg非平衡状态;豫南黑猪群体多态信息含量为0.24(PIC<0.25),属于低度多态;杂合度为0.28。
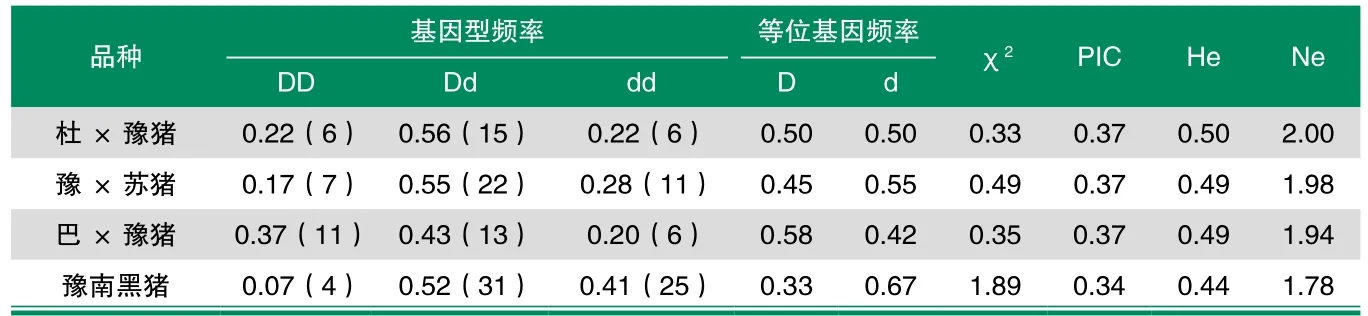
表4 HaeIII-RFLP位点在豫南黑猪及其杂交后代中基因型频率分布
表4显示,HaeIII-RFLP位点基因型频率在杜×豫猪、豫×苏猪、巴×豫猪和豫南黑猪中χ2值分别为0.33、0.49、0.35和1.89,均小于χ=5.99,因此差异不显著(P>0.05),群体处于Hardy-Weinberg平衡状态。豫南黑猪群体多态信息含量为0.34,属于中度多态(0.25<PIC<0.5),杂合度为0.44。
3 讨论
前人研究报道,位于H-FABP基因的5’-UTR的Hinf I酶切多态位点,是通过影响H-FABP基因的表达进而影响IMF的含量[11]。在本研究中,该位点豫南黑猪、豫×苏和杜×豫杂交群体中H等位基因为优势等位基因,与Gerberns(1997)等[9]研究在杜洛克群体中的研究结果相一致。Gerbens(1999)等[18]进一步分析了H-FABP基因在杜洛克群体中与肉质性状的关联,结果显示Hinf I-RFLP位点HH型个体的肌内脂肪(IMF)含量高于其他基因型,而豫南黑猪作为地方品种猪,具有肌内脂肪含量高、肉质性状好等优点[17],因此,可推断本研究结果的可靠性,在本试验群体中与肉质性状间的相关性有待于进一步的研究。
曾勇庆[19]及庞卫军等[20]一致认为aaddHH(MspI-a、HaeIII-d、Hinf I-H)具有较高的IMF含量。本研究中HaeIII酶切位点上,各组试验猪均表现多态性,豫南黑猪和豫×苏猪群体中有利等位基因d占优势,杜×豫猪和巴×豫猪群体中等位基因d的频率分别为0.50和0.42。在Hinf I*-RFLP位点,巴×豫猪的优势等位基因为D,频率为0.58,与李桢等[21]等研究结果相一致。张桂香等[22]和李桢等[23]研究发现,MspI-RFLP位点在中国地方猪种中不具有多态性,而在本研究试验群体中均表现多态,分析可能与豫南黑猪培育过程中导入了杜洛克血统有关,因此后续将利用淮南猪及豫南黑猪作为供试群体,进一步研究H-FABP基因的多态位点与生产性能的相关性。此外,本试验首次发现该豫南黑猪及其杂种猪的HaeIII、MspI和Hinf I*酶切位点处于连锁平衡状态,该研究结果进一步揭示了H-FABP基因在试验群体中的遗传规律,但至于在其他猪种中是否存在连锁,有待进一步确证。
4 结论
本研究中,豫南黑猪及其杂交后代猪的H-FABP基因在5’-UTR和intron2的4个酶切位点均存在多态性,且在intron2的HaeIII、MspI和Hinf I*酶切位点处于连锁平衡状态。本研究结果,可为将来筛选出合适有效的肉质性状相关的分子标记提供重要的理论基础。
参考文献
[1] P A T R I C I A . E a t i n g q u a l i t y of pork in Denmark [J].Pig Farming(Supplement),1985(10):56-57.
[2] DE VOL DL,MCKEITH F K,BECHTEL P L.Variation in composition and palatability traits and relationships between muscle characteristics and palatability in a random sample of pork carcasses [J]. J Anim Sci, 1988, 66(2): 385-395.
[3] H O V E N I E R R,K A N I S E,VERHOEVEN J A.Repeatability of taste panel tenderness scores and their relationships to objective pig meat quailty traits [J].J Anim Sci,1993,71(8):2018-2025.
[4] F E R N A N D E Z X,M O N I N G,TALMANT A,et al.Influence of intramuscular fat content on the quality of pig meat[J].Meat Science,1999, 53: 59-65.
[5] WOOD JD,RICHARDSON RI,NUTE G R,et al. Effects of fatty acids on meat quality:a review[J]. Meat Science, 2003, 66(1):21-32.
[6] 姜延志,刘晓妍,李芳琼,等.猪H-FABP基因多态性及其与肌内脂肪含量的相关研究[J].畜牧兽医学报, 2010, 41(7): 792-796.
[7] HOVENIER R,KANIS E,VAN ASSELDONK T,et al.Genetic parametera of pig meat quality traits in a halothane negative population[J].Livestock Production Science,1992(32):309-321.
[8] SCHAAP F G, VAN DER VUSSE G J, GLATZ J F. Fatty acid-binding proteins in the heart[J]. Mol Cell Biochem,1998, 180: 43-51.
[9] GERBENS F,RETTENBERGER G,LENSTRA J A,et a1. Characterization, chromosomal localization and genetic variation of the porcine heart fatty acid-binding protein gene[J].Mamm Genome, 1997,8(5): 328-332.
[10] DE KONING D J, JANSS L L, RATTINK A P,et al.Detection of quantitative traits loci for backfat thickness and intramuscular fat content in pigs(Sus scrofa)[J]. Genetics,1999, 152(4): 1679-1690.
[11] GERBENS F,DE KONING D J,HARDERS F L,et al.The effect adipocyte and heart fatty acid-binding protein genes on intramuscular fat and backfat content in Meishan crossbred pigs[J].J Anim Sci,2000,78(3):552-559.
[12] LEE SH,CHOI YM,CHOE JH, et al. Association between polymorphisms of the heart fatty acid binding protein gene and intramuscular fat content, fatty acid composition, and meat quality in Berkshire breed [J]. Meat Sci. 2010, 86(3):794-800.
[13] HERMESCH S,LUXFORD B G, GRASER H U.Genetic parameters for lean meat yield,meat quality, reproduction and feed efficiency traits for Australian pigs[J].Livestock Production Science, 2000,65:239-248.
[14] 林万华,黄路生,任军,等.中外十个猪种H-FABP基因遗传变异的研究[J]. 遗传学报, 2002, 29(1): 12-15.
[15] 张越,陆萌,朱卫华,等.猪心脏脂肪酸结合蛋白基因多态性检测及与肉质性状的关联分析[J].中国畜牧杂志, 2016, 52(1):7-12.
[16] 王明,李新建,冯富彦,等.豫南黑猪生理生化指标的初步分析[J].江苏农业科学,2009(1):196-198.
[17] 柳絮.国家级猪新品种“豫南黑猪”通过审定[J]. 广西农学报,2008(6): 108.
[18] GERBENS F,VAN ERP A J M, HARDERS F L, et a1.Effect of genetic variants of the heart acid bind-ing protein gene on intramuscular fat and performance traits in pigs[J].J Anim Sci, 1999, 77(4): 846-852.
[19] 曾勇庆.莱芜猪肉质特性及其分子遗传学基础的研究[D].江苏:南京农业大学, 2004.
[20] 庞卫军,孙世铎,李影,等.西部地区主要猪种和野猪H-FABP基因分子标记与IMF含量关系[J].遗传,2005, 27(3):35l-356.
[21] 呼红梅,王继英,郭建凤,等.莱芜猪和杜洛克猪肌肉H-FABP基因表达量与肌内脂肪和脂肪酸含量关联分析[J]. 华北农学报,2010, 25(4): 64-68.
[22] 张桂香,曹红鹤,王立贤,等.9个猪种H-FABP基因5’-上游区和第二内含子的遗传变异[J].畜牧兽医学报, 2002,33(4):340-343.
[23] 李桢,曹红鹤,储明星,等.中外11个猪种H-FABP基因PCRRFLP的研究[J].畜牧兽医学报,2003,34(4):313-317.
基金项目:河南省生猪产业技术体系(S2012-06);河南省科技攻关项目(142102110047);郑州科技创新团队(141PCXTD513)
作者简介:郭吉利(1989-),女,河南巩义人,硕士研究生,主要从事动物遗传育种与繁殖学研究。
*通讯作者:李新建(1977-),男,副教授,硕士生导师,主要从事动物遗传育种学研究。
收稿日期:(2016-03-30)
