杉木—光皮桦混交林对微生物量碳氮与酶活性的影响1)
2015-04-03周静
周静
(四川师范大学,成都,610101)
袁华明 李佳泳 张炜 龙汉利 李晓清 陈忠
(四川青川县林业局) (四川省林业科学研究院) (内江市东兴区林业局)
微生物是全球生态系统的重要组成部分,在物质循环、能量流动过程中担任着极其重要的角色,并且是地球碳氮生物化学循环过程中的主要驱动者[1-2],而它对环境变化极其敏感,其呼吸强度是衡量土壤微生物总的活性指标,所以长期以来,与之相紧密联系的土壤养分、微生物生物量和酶活性的研究受到广泛重视[3-4]。土壤微生物生物量是土壤有机质中最活跃和最易变化的部分[5],土壤微生物生物量含量的高低是衡量土壤肥力的重要指标之一,土壤微生物量碳、氮是土壤碳、氮的重要贮备库[6]。土壤中的各种生物化学反应除了受土壤中微生物本身影响之外,各种各样的土壤酶也是土壤生物活性的主要调节者,在土壤有机质转化和养分循环中起着极其重要的作用,其活性高低可反映土壤中各种生物化学反应的强度和方向[7-9],能较好地反应土壤肥力,而地上植被的类型则对土壤微生物活性的活跃程度有重要意义[6]。
杉木(Cunninghamia lanceolata)是我国西南地区主要的造林树种[8],也是在四川地区广泛种植的商品林树种,该树种具有产量高、生长快、材质好、用途广等特点,但由于四川部分山地土壤贫瘠且自肥能力较差,林分结构单一的杉木纯林出现了生长量下降,林分抗逆性较差,林区生态系统恶化,林地土壤肥力衰退的恶性循环。通过调整林分结构,引入阔叶林树种(如桦木、桤木)搭配杉木组成杉木混交林模式可以解决山地人工林地力衰退,促进林木生长进而提高蓄积量,改善生态环境等问题[10]。由于混交林和纯林树种组成的差异性会影响林下土壤微生物生物量、土壤酶活性,进而进一步影响林下土壤有机质及养分转化。鉴于此,本研究基于川北地区杉木低效林块状混交改造技术示范研究,通过对杉木—光皮桦(Betula luminifera)混交林(以下简称杉桦混交林)对土微生物量碳、氮、有机质、有效氮以及几种重要的土壤酶活性进行研究,探讨杉桦混交模式在这种山地条件下提高地力、促进土壤养分循环与转化的作用机制,为川北山地地区杉木低效林的改造提供理论依据,也为该山区杉木混交林土壤微生物量碳氮与酶活性的相关研究提供理论依据,并为该地区的生态环境恢复重建提供科学依据,促进该地区的可持续发展。
1 研究区概况
研究区位于广元市青川县孔溪乡和兴村,地处四川盆地北部边缘,白龙江下游,川、甘、陕三省结合部,地貌以低山为主。该地区属亚热带温润季风气候,春迟、夏短、秋凉、冬长,年平均降水量1 021.7 mm,年均日照时间1 292 h,年无霜期243 d。现阶段,研究区内以杉木纯林、光皮桦纯林以及杉木—光皮桦混交林为主。土壤主要为山地黄壤,土层比较浅,多数为50 cm 左右。
2 研究方法
2.1 样地设置
试验地位于广元市青川县孔溪乡和兴村,该林分于20 世纪70 年代营造杉木纯林,当时的初植密度4 300 株·hm-2,1997 年间伐补种杉木、光皮桦。在研究区内选择坡位、坡向、坡度等均一致的立地条件,同属1997 年造林且生长状况及管理经营水平大致相同的杉木、光皮桦纯林以及杉桦混交林(1 ∶1混交),分别设立3 块20 m×20 m 的样地。样地基本概况见表1。
2.2 土壤采集
2013 年11 月在每个设置的标准地内,按照“S”形选择9 个采样点,根据其土层深度和土色等特征,将土壤层划分0 ~5、>5 ~20、>20 ~40 cm 3 个层次,采用直径为6 cm 的土钻采集各土层土壤,将同一标准地内各层土壤分别混合后按四分法取1.0 kg 土样,将取好的土样放入灭菌封口袋中置于有冰袋的保鲜盒中运回实验室,去掉可见植物残根和粗石粒,过2 mm 筛后混合均匀,均分为两份:一份在常温下风干,用于测定土壤有机质、土壤有效氮;另一份置于4 ℃保存,用于土壤微生物生物量、土壤酶测定。

表1 川北地区样地基本概况
2.3 室内分析
土壤有机质由过100 目筛的土样选用重铬酸钾氧化-外加热法测定。有效氮由过100 目筛的土样采用碱基解扩散法测定[11]。土壤酶、微生物生物量测定方法主要参考《土壤与环境微生物研究方法》[12]:①脲酶(URE)活性采用柠檬酸比色法;②过氧化氢酶(CAT)采用邻苯三酚比色法;③蔗糖酶(INV)采用3,5-二硝基水杨酸比色法;④酸性磷酸酶(ACP)采用的是对硝基苯磷酸二钠比色法;⑤土壤微生物生物量(SMBC、SMBN)采用氯仿熏蒸法提取测定。
2.4 数据分析
试验数据采用SPSS 20.0 分析,研究数据整理及制图在Excel 2007 上进行。不同造林模式的土壤微生物量碳氮和土壤酶活性的差异评价采用单因素方差分析和Duncan 新复极差法,其相关性分析采用两变量相关分析法。
3 结果与分析
3.1 不同造林模式对土壤有机质及有效氮的影响
在各造林模式下,随着土层深度的加大土壤有机质、有效氮质量分数均呈现极显著降低的趋势(p<0.01);在各相同土层,杉桦混交林的土壤有机质、有效氮质量分数均显著高于杉木纯林和光皮桦纯林(p<0.05),在0~5 cm 土层表现最为明显,杉桦混交造林模式下的土壤有机质分别高于杉木纯林和光皮桦木纯林24.4%和15.7%,土壤有效氮分别高于杉木纯林和光皮桦纯林33.7%和28.7%(表2)。
3.2 不同造林模式对土壤微生物生物量的影响
在不同造林模式下不同土层土壤微生物量碳、氮均呈显著性差异(p<0.05)。杉桦混交林模式下的土壤SMBC(平均259.23 mg·kg-1)、SMBN(平均33.28 mg·kg-1)均显著高于光皮桦纯林与杉木纯林模式(p<0.05)(表3)。在各造林模式内,0~5 cm 土层SMBC、SMBN 质量分数最高,说明土壤微生物量具有表聚性。随着土层深度的加深,微生物生物量碳、氮呈显著下降趋势(p<0.05)。

表2 不同造林模式下土壤有机质和有效氮质量分数

表3 不同造林模式下土壤微生物生物量
3.3 不同造林模式下土壤酶分布特征
各林地土壤酶活性随着土层深度加深而减小,但各造林模式下土壤酶活性有差异(表4)。杉桦混交林以及杉木、光皮桦纯林的各林地土层间CAT 活性无显著性差异(p>0.05),也不存在规律性,另外,相同土层杉桦混交林土壤CAT 活性比杉木、光皮桦纯林略高但差异不显著(p>0.05)。相同土层杉桦混交土壤URE 活性均高于杉木、光皮桦纯林,且差异显著(p<0.05),杉桦混交模式0 ~5 cm 土层URE活性最高,达到0.53 mg·g-1。杉桦混交模式与杉木、光皮桦纯林模式相比很大程度上提高了土壤URE 活性,进而改善土壤氮素供应。杉桦混交模式各层土壤INV 活性均显著高于杉木、光皮桦纯林模式(p<0.05),杉桦混交模式0 ~5 cm 土层INV 活性最高,达到31.79 mg·g-1;但随着土壤土层深度的增加,土壤提供微生物碳源、氮源的能力逐渐减弱,使得各模式林地土壤INV 活性显著降低(p<0.05),杉桦混交模式>20 ~40 cm 土层的INV 活性下降至11.56 mg·g-1,杉木纯林>20 ~40 cm 土层INV 活性则最低,为7.54 mg·g-1。不同林地模式、不同土层ACP 活性差异显著(p<0.05),杉桦混交模式土壤ACP 活性(平均6.53 mg·g-1)显著高于杉木(平均3.06 mg·g-1)和光皮桦(平均4.13 mg·g-1)纯林模式(p<0.05)。
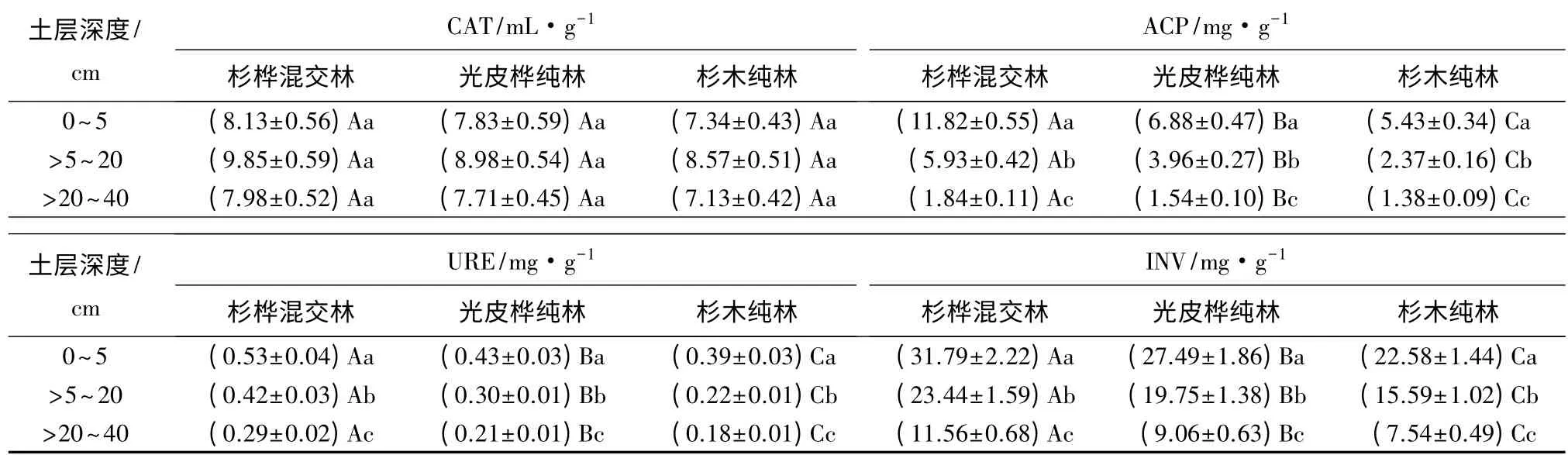
表4 土壤酶活性分布
3.4 土壤微生物特性与土壤酶活性的相关性
由土壤微生物特性与土壤酶活性的相关性分析(表5)可知,SMBC、SMBN、有机质、有效氮4 个因素,两两之间均呈极显著正相关。INV 仅与SMBC、有机质呈显著正相关。URE 仅与SMBN、有效氮呈显著正相关。ACP 与SMBC、有机质呈极显著正相关。CAT 与SMBC、SMBN、有机质以及有效氮无显著相关性。土壤酶之间也存在一定相关性,INV 与URE 之间存在显著正相关性,ACP 与INV、URE 存在极显著正相关性,而CAT 与ACP、INV、URE 均不存在相关性。

表5 土壤微生物特性与土壤酶活性的相关系数
4 结论与讨论
在相同土层,杉桦混交造林模式下土壤有机质、有效氮质量分数均显著高于杉木、光皮桦纯林模式;杉桦混交模式林地内土壤微生物生物量显著高于杉木、光皮桦纯林模式。其原因可能由于在杉桦混交林模式林地内,土壤碳库、氮库从林木凋落物和根系分泌物得到不断补充,使得土壤微生物生物量显著增大。可见,杉桦混交模式优于光皮桦纯林、杉木纯林模式。杉木纯林在引进光皮桦后改善了林分结构,改变了凋落物组成,增加了凋落量[13],即杉桦混交造林模式比光皮桦、杉木纯林更利于土壤有机质、有效氮的积累,杉桦混交林模式相比于杉木、光皮桦纯林,土壤肥力程度、微生物生物量得到显著提高,土壤健康状况得到改善,这与许多学者[14-15]研究结果相一致。
本研究发现,不同模式及其相同模式不同土层的林地土壤酶活性存在显著差异。杉桦混交林地内土壤CAT 活性均略高于杉木、光皮桦纯林,杉桦混交林地内土壤ACP、INV、URE 活性显著高于杉木、光皮桦纯林。CAT 参与腐殖质的合成其活性与土壤腐殖化强度和有机质的积累紧密相关[16];URE 是一种酰胺酶,是土壤中唯一对尿素水解起重要作用的关键性酶,可将土壤中的有机氮水解为植物可直接利用的有效态氮,对提高氮素的利用率和促进土壤氮素循环具有重要意义[17];INV 直接参与土壤有机物质的代谢过程,其强弱可作为土壤质量、熟化程度和肥力水平的评价指标[18];磷酸酶能促进有机磷化合物的水解,土壤的磷酸酶活性大小可以表征土壤肥力状况的好坏(特别是磷的状况)。杉桦混交模式在引进光皮桦后,增加了主林层的生物多样性,改善林分结构,改变了林内环境,增加了林内散射光强度,提高了林内温度以及水分状况,使得土壤生物活性增加,从而促进各种土壤酶活性增强[19]。种种迹象表明,杉桦混交林土壤肥力、土壤质量及土壤健康均优于杉木、光皮桦纯林,与万福绪等[16]研究结果大致相同。本研究发现,随着土层的加深各模式林地ACP、INV、URE 活性显著降低,这分别与陈永川[20]等、闫晗等[21]的研究结论一致。然而随着土壤深度的增加,CAT 活性无差异性,可能是由于CAT 在土壤中起清除H+毒害作用,而各个模式林地土壤的H+浓度处于较低水平,对土壤无毒害作用,所以各土层中CAT 活性未表现出差异性。
本研究发现,微生物生物量与土壤酶活性以及土壤酶活性之间存在一定相关性,与李东坡等[22]的研究基本一致。土壤微生物量是土壤活性养分储存库,是植物赖以生存的源泉[23-24]。土壤酶参与土壤中各种生化反应,如动、植物残体和微生物的分解,以及合成有机化合物的水解与转化[25-26]。由此可见,各种微生物、土壤酶都不是单一的个体,是不可分割的整体,在相互制约和相互促进中发挥着作用。
总之,杉桦混交模式较杉木、光皮桦纯林模式,土壤微生物生物量、有机质、有效氮以及土壤酶活性都得到改善,土壤肥力也有较大的提高,更加符合山地造林的可持续发展。本研究受时间、空间条件限制,研究内容还有待完善加强,但也从侧面为杉桦混交造林模式比杉木、光皮桦纯林模式速生丰产提供了一定理论依据。因此,建议加强杉桦混交林的营造,达到生态和经济效益双丰收。
[1] 孔维栋.极地陆域微生物多样性研究进展[J].生物多样性,2013,21(4):456-467.
[2] 李廷茂,胡江春,汪思龙,等.森林生态系统中土壤微生物的作用与应用[J].应用生态学报,2004,15(10):1943-1946.
[3] 许景伟,王卫东,李成.不同类型黑松混交林土壤微生物、酶及其与土壤养分关系的研究[J].北京林业大学学报,2000,22(1):51-55.
[4] 马丹,罗成德,雷波,等.川西亚高山岷江冷杉林土壤生物肥力特征研究[J].西北农林科技大学学报:自然科学版,2014,42(3):85-92.
[5] Warder D A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J].Biological Reviews,1992,67(3):321-358.
[6] 马文文,姚拓,靳鹏,等.荒漠草原2 种植物群落土壤微生物及土壤酶特征[J].中国沙漠,2014,34(1):176-183.
[7] Max M C,Wood M,Jarvis S C. A micro plate flurimetric assay for the study of enzyme diversity in soils[J].Soil Biology & Biochemistry,2001,33(12):1633-1640.
[8] 肖慈英,阮宏华,屠六邦.宁镇山区不同森林土壤生物学特性的研究[J].应用生态学报,2002,13(9):1077-1081.
[9] 胡斌,段昌群,王震洪.植被恢复措施对退化生态系统土壤酶活性及肥力的影响[J].土壤学报,2002,39(4):604-608.
[10] 林同龙.杉木人工林近自然经营技术的应用效果研究[J].中南林业科技大学学报,2012,3(32):11-16.
[11] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[12] 李振高,骆永明,滕应.土壤与环境微生物研究法[M].北京:科学出版社,2008.
[13] 俞新妥.杉木栽培学[M].福州:福建科学技术出版社,1997.
[14] Burke I C,Lauenroth W K,Riggle R,et al. Spatial variability of soil properties in the shortgrass steppe:the relative importance of topography,grazing,microsite,and plant species in controlling spatial patterns[J].Ecosystems,1999,2(5):422-438.
[15] 樊后保,李燕燕,苏兵强,等.马尾松-阔叶树混交异龄林生物量与生产力分配格局[J].生态学报,2006,26(8):2463-2473.
[16] 万福绪,杨东.苏北海堤杉木杨树混交林林木生长及土壤肥力研究[J]南京林业大学学报:自然科学版,2006,30(2):43-46.
[17] Bohme L,Langer U,Bohme F. Microbial biomass,enzyme activities and microbial community structure in two European longterm field experiments[J].Ecosystems and Environment,2005,109(1):141-152.
[18] 李亮,包耀贤,廖超英,等.乌兰布和沙漠东北部沙区人工林土壤微生物及酶活性研究[J].西北植物学报,2010,30(5):987-994.
[19] 陈莉莉,王得祥,于飞,等.松栎混交林土壤微生物数量与土壤酶活性及土壤养分关系的研究[J].土壤通报,2014,45(1):77-84.
[20] 陈永川,杨春霞,黎小清,等.管理模式对胶园土壤酸性磷酸酶活性动态的影响[J].热带作物学报,2013,34(7):1213-1217.
[21] 闫晗,葛蕊,潘胜凯,等.恢复措施对排土场土壤酶活性和微生物量的影响[J].环境化学,2014,33(2):327-333.
[22] 李东坡,武志杰,陈利军.有机农业施肥方式对土壤微生物活性的影响研究[J].中国生态农业学报,2005,13(2):99-101.
[23] 薛萐,刘国彬,戴全厚,等.不同植被恢复模式对黄土丘陵区侵蚀土壤微生物量的影响[J].自然资源学报,2007,22(1):20-27.
[24] 周丽霞,丁明懋.土壤微生物学特性对土壤健康的指示作用[J].生物多样性,2007,15(2):162-171.
[25] Moscatelli M C,Fonck M,Angelis P,et al. Mediterranean natural forest living at elevated carbon dioxide:soil biological properties and plant biomass growth[J].Soil Use and Management,2001,17(3):195-202.
[26] 刘纯,刘延坤,金光泽.小兴安岭6 种森林类型土壤微生物量的季节变化特征[J].生态学报,2014,34(2):451-459.
