4 种园林树木树冠微环境与增湿降温的关系1)
2015-04-03刘瑞文孙晓刚郭太君林萌
刘瑞文 孙晓刚 郭太君 林萌
(吉林农业大学,长春,130118)
园林植物的生态功能在城市环境绿化中发挥着不可或缺的重要作用,园林植物通过吸收CO2释放O2、增湿降温、吸收有害气体、降噪除尘等可以有效地改善环境质量,在城市化加快发展的现代社会中给人们提供一个更好的生活环境。
树冠是园林树木增湿降温和固碳释氧的主要场所。叶幕(树冠)微气候是具有一定结构形式的叶幕,对其自然气候因素经过若干不同级别的“环境—植物”界面过滤后形成的叶幕紧密表面和叶幕内部的微气候条件[1]。由于树冠是一个立体的空间结构,其内部的不同部位叶片所处的微环境也有所不同,从而可能影响不同部位叶片的蒸腾作用能力产生强弱的差别。林萌等[2]指出,9 种园林树木树冠南侧水平方向由外向内不同部位的单位叶面积增湿、降温量的变化,由树冠表层向树冠中心顺次降低,并提出了园林树木增湿、降温量评价的适宜取样部位。冷平生等[3]对5 种园林树木向阳成熟叶片蒸腾特性的研究发现,蒸腾速率主要受光合有效辐射、叶片温度和气孔导度等的影响。王华田等[4]研究发现,辐射强度和风速是控制林冠蒸腾耗水的决定因素。在果树生产方面,通过对不同树形冠层微气候的研究,总结出有利于提升果实产量和品质的树形结构[5-8]。
园林树木增湿降温生态功能,不仅与园林树木的种类有关,也受环境条件的制约。由于树木的生长特性不同,其树冠内不同方位和部位接受的微环境亦有差别,可能影响树冠不同方位各部位的增湿降温生态效益。因此,文中旨在通过对4 种园林树木树冠不同方位各部位微环境和增湿降温的研究,分析园林树木增湿降温生态功能的合理取样部位以及增湿降温生态功能与树冠微环境的关系,以期更好地为园林树木增湿降温生态功能的评价和提高园林树木增湿降温生态效益提供科学依据。
1 试验地概况
试验地位于吉林农业大学校园内。吉林农业大学位于吉林省长春市的东南部,属温带大陆性季风气候,四季分明。年平均降水量522 mm,年平均气温4.8 ℃,最高气温39.5 ℃,最低气温-39.8 ℃,日照时间2 688 h。
试验选择2013 年8 月下旬天气晴朗的日期进行(共进行8 d),期间的环境条件平均值为:空气温度30.9 ℃;光照强度1 641.95 μmol·m-2·s-1;空气相对湿度44.37%;风速0.86 m·s-1;大气中CO2摩尔分数394.85 μmol·mol-1。
2 材料与方法
选择4 种常见的园林树木作为试验材料(表1)。
将树冠在水平方向上分为东侧、南侧、西侧、北侧四个方位,再将每个方位由外向内分为表层(距树冠表面10 cm,记为表层A),之后再三等分,依次记为外层(B)、中层(C)和内层(D)。分别取上述各层的中间部位叶片,具体分层位置及各测定点的位置如图1 所示[2]。

表1 试验材料基本情况
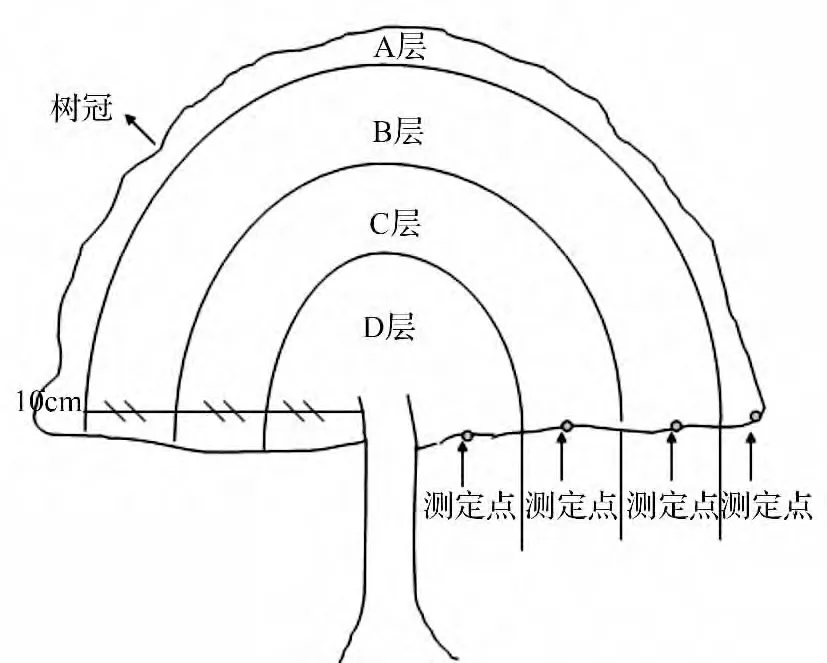
图1 树冠分层及测定点分布模式图
采用美国PP Systems 国际有限公司生产的CIRAS-2 便携式光合作用测定系统,测定蒸腾速率、光照强度、空气温度、叶面温度、空气相对湿度、空气CO2摩尔分数、细胞间隙CO2摩尔分数、气孔导度;使用苏州特安斯有限公司生产的TASI-8818数字式风速计,测定树冠不同部位风速。测定时间段为每天的08:00—16:00,每2 h 测定一次,每天测定两个树种,每个树种8 d 分别测定4 次,每个树种后两次测定顺序与前两次相反。每种树木选择长势相似、生长环境相同的3 株,每个测定点选取3 片叶片,每个叶片3 次取值,最后结果取平均值。
增湿、降温量的计算:根据测定各树种各测定点的蒸腾速率,计算试材当日蒸腾总量。公式[9]为:
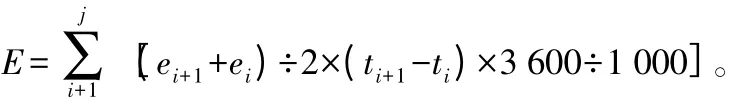
式中:E 为测定日的蒸腾总量(mol·m-2·d-1);ei为初测点的瞬时蒸腾速率(mmol·m-2·s-1);ei+1为下一测点的瞬时蒸腾速率(mmol·m-2·s-1);ti为初测点的瞬时时间(h);ti+1为下一测点的时间(h);j为测试次数。根据下述公式计算日蒸腾量(g·m-2·d-1)。
日蒸腾水分质量:WH2O=E×18。
水分蒸发吸收热量:M=WH2O×L×4.18。
式中:M 为单位叶面积每日吸收的热量,单位为J·m-2·d-1;L 为蒸发耗热系数(L=597-0.57×t,t 为测定日的温度);4.18 为1 cal=4.18 J。由此可计算出各植物每平方米叶片在测定日吸收热量的值。又因植物蒸腾消耗热量(M)是取自于周围1 000 m3的空气柱(10 m2×100 m),使该气柱温度下降。故气温下降值用下式计算[8]。
蒸腾吸热的气温下降值:ΔT=M/PC。
式中:ΔT 为下降温度(℃);PC为空气容积热容量,为1 256 J·m-3·h-1。
测定数据的统计分析通过EXCEL 软件、SPSS数据处理软件进行。
3 结果与分析
3.1 树冠不同部位和不同方位增湿、降温量的变化
树冠同一部位不同方位间单位叶面积增湿、降温量的变化,由大到小呈现为南侧、东侧、西侧、北侧;同一方位不同部位间由大到小呈现为表层、外层、中层、内层的变化规律(表2)。如树冠南侧、东侧、西侧、北侧各方位的单位叶面积增湿量依次为3.62、3.30、2.65、2.07 kg·m-2·d-1,降温量依次为0.29、0.26、0.21、0.17 ℃;树冠南侧的单位叶面积增湿、降温量较北侧分别提高了74.88%、70.59%。表层、外层、中层和内层的单位叶面积平均增湿量依次是4.13、3.24、2.55、1.70 kg·m-2·d-1,降温量分别为0.33、0.26、0.20、0.15 ℃;表层较内层单位叶面积增湿、降温量分别提高了142.94%、120.00%。树冠整体单位叶面积增湿量平均为2.91 kg·m-2·d-1、降温量平均为0.235 ℃,其与树冠不同部位比较,表层和外层单位叶面积平均增湿、降温量较平均值分别提高了41.92%、40.43%和11.34%、10.64%,中层和内层较平均值分别下降了12.37%、14.89%和41.58%、36.17%;与树冠不同方位比较,树冠南侧和东侧单位叶面积平均增湿、降温量较平均值分别提高了24.4%、23.4%和13.4%、10.64%,树冠西侧和北侧较平均值分别下降了8.93%、10.64%和28.87%、27.66%。
上述结果说明,树冠不同部位间的增湿、降温量差异程度大于树冠的不同方位,树冠西侧和外层更接近树冠不同部位及不同方位单位叶面积增湿、降温量的平均值。表明在进行园林树木增湿降温生态功能评价时,合理的取样部位应是树冠西侧外层。以树冠西侧外层为园林树木增湿降温生态功能评价的取样部位,4 种试材的单位叶面积增湿、降温量在不同树性间差异较大,而在相同树性间基本相似。如属于乔木型的桃叶卫矛和山杏树冠西侧外层单位叶面积增湿量平均为3.16 kg·m-2·d-1、降温量平均为0.25 ℃,属于灌木型的大花水桠木和红瑞木树冠西侧外层单位叶面积增湿量平均为2.7 kg·m-2·d-1、降温量平均为0.21 ℃,前者是后者的1.2 倍;属于灌木型的大花水桠木和红瑞木单位叶面积平均增湿、降温量分别为2.64 kg·m-2·d-1、0.21 ℃和2.72 kg·m-2·d-1、0.22 ℃;属于乔木型的山杏和桃叶卫矛分别为3.08 kg·m-2·d-1、0.25 ℃和3.18 kg·m-2·d-1、0.26 ℃,差别都不大。
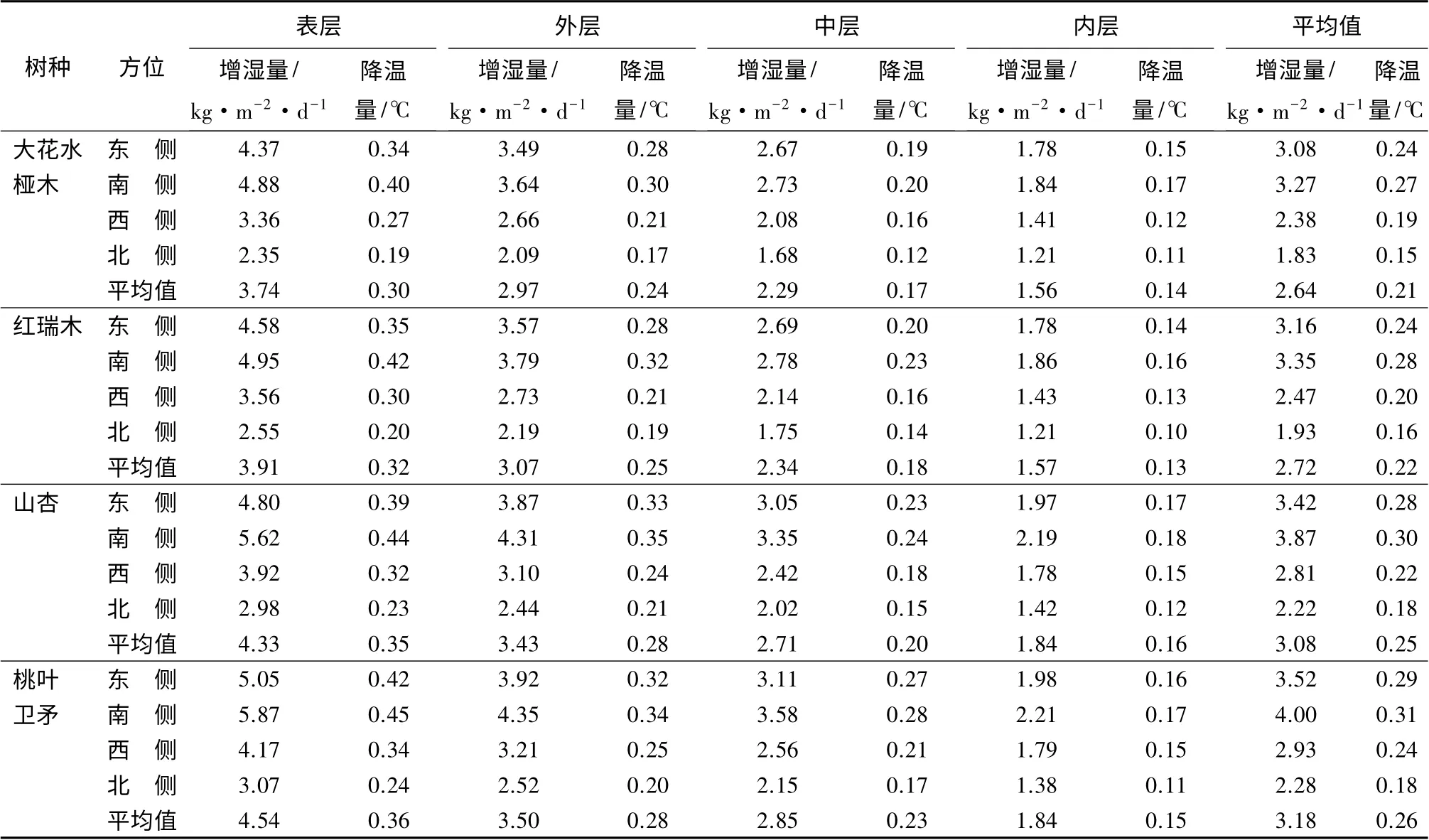
表2 不同树种树冠不同方位及不同部位叶片单位叶面积增湿、降温量的变化
3.2 树冠不同部位微环境的变化
以树冠不同方位各测定部位空气CO2摩尔分数(CR)、光照强度(Q)、空气温度(TI)、叶面温度(TC)、空气相对湿度(RH)和风速(WS)6 个微环境,以及叶片的胞间CO2摩尔分数(CI)和气孔导度(GS)的平均值,统计分析树冠不同部位上述8 个测定指标由树冠表层至内层的变化特点,结果呈现出比较明显的变化规律(表3)。各树种CR的平均值由表层至内层基本相同。各树种CI和RH的平均值由树冠表层至内层呈逐渐升高的趋势,除山杏不同部位的RH差异不明显外,表层均显著低于其他各部位。如CI外层至内层较表层平均提高了18.52%~26.46%,RH提高了6.17%~10.40%。而各树种Q、TI、TC、GS和WS的平均值由树冠表层至内层呈现大多数下降的趋势,除山杏表层的TC与外层差异不明显外,表层均极显著高于其他各测定部位。各树种Q 的平均值外层至内层较表层平均下降了43.66%~95.22%;GS和WS的平均值分别较表层平均下降了22.14%~60.34%和25.61%~66.67%;TI和TC的平均值分别较表层降低1.7 ~2.1 ℃和1.4 ~1.8 ℃。在测定的8 个指标中,4 个树种不同部位与其平均值比较,基本表现出与外层接近,仅CI(大花水桠木、山杏)、GS(红瑞木、山杏)、RH(山杏)与中层更接近一些。
乔木与灌木相比,两种乔木与灌木树冠表层与内层微环境的变化幅度,Q、GS和RH表现为灌木大于乔木的趋势,CI、TI、TC和WS表现为乔木大于灌木。如两种灌木内层的Q 较表层下降了99.10%,两种乔木下降了91.23%;两种灌木内层的CI较表层提高了13.06%,两种乔木为24.38%。

表3 不同树种树冠不同部位微环境的变化
3.3 树冠不同方位微环境的变化
以不同树种树冠各方位表层、外层、中层和内层各测定部位CR、Q、TI、TC、RH、CI、GS和WS的平均值,分析比较树冠东侧、南侧、西侧和北侧4 个方位微环境的变化规律,其结果呈现出与树冠不同部位微环境相似的变化规律(表4)。各树种CR的平均值在树冠东侧、南侧、西侧和北侧4 个方位间基本相似,其他各测定指标在不同方位间多表现出显著或极显著差异。其中,各树种Q、TI、TC、GS和WS的平均值由大到小的顺序均表现为树冠南侧、东侧、西侧、北侧;各树种CI和RH的平均值与上述微环境变化相反,由大到小的顺序均呈现北侧、西侧、东侧、南侧;树冠不同方位上述8 项测定指标与全株各方位平均值比较,无论是各个树种,还是4 个树种的平均值,均表现出树冠西侧与各方位的平均值比较接近。
乔木与灌木相比,两种灌木与乔木树冠北侧较南侧微环境的变化幅度,呈现为Q 和TC灌木(-94.93%和-2.53 ℃)与乔木(-94.14%和-2.50 ℃)相似,WS灌木(-66.67%)小于乔木(-81.67%)的变化幅度,其他表现为灌木大于乔木的。
综合表3 和表4,对比分析树冠不同部位与不同方位的微环境差异,进一步探讨园林树木增湿、降温量与树冠微环境之间的关系,以及增湿降温生态功能评价的合理取样方法。CR在树冠不同部位以及不同方位间基本相同,说明树冠中CR的变化与取样方法基本无关。Q、GS、TI、TC和WS表现为表层和树冠南侧最高,内层和树冠北侧最低;CI和RH表现为内层和树冠北侧最高,表层和树冠南侧最低。在树冠不同部位及不同方位的微环境变化幅度方面,树冠表层与内层、树冠南侧与北侧比较,均呈现Q、GS和WS下降幅度较大,CI、TI、TC和RH下降幅度较小。
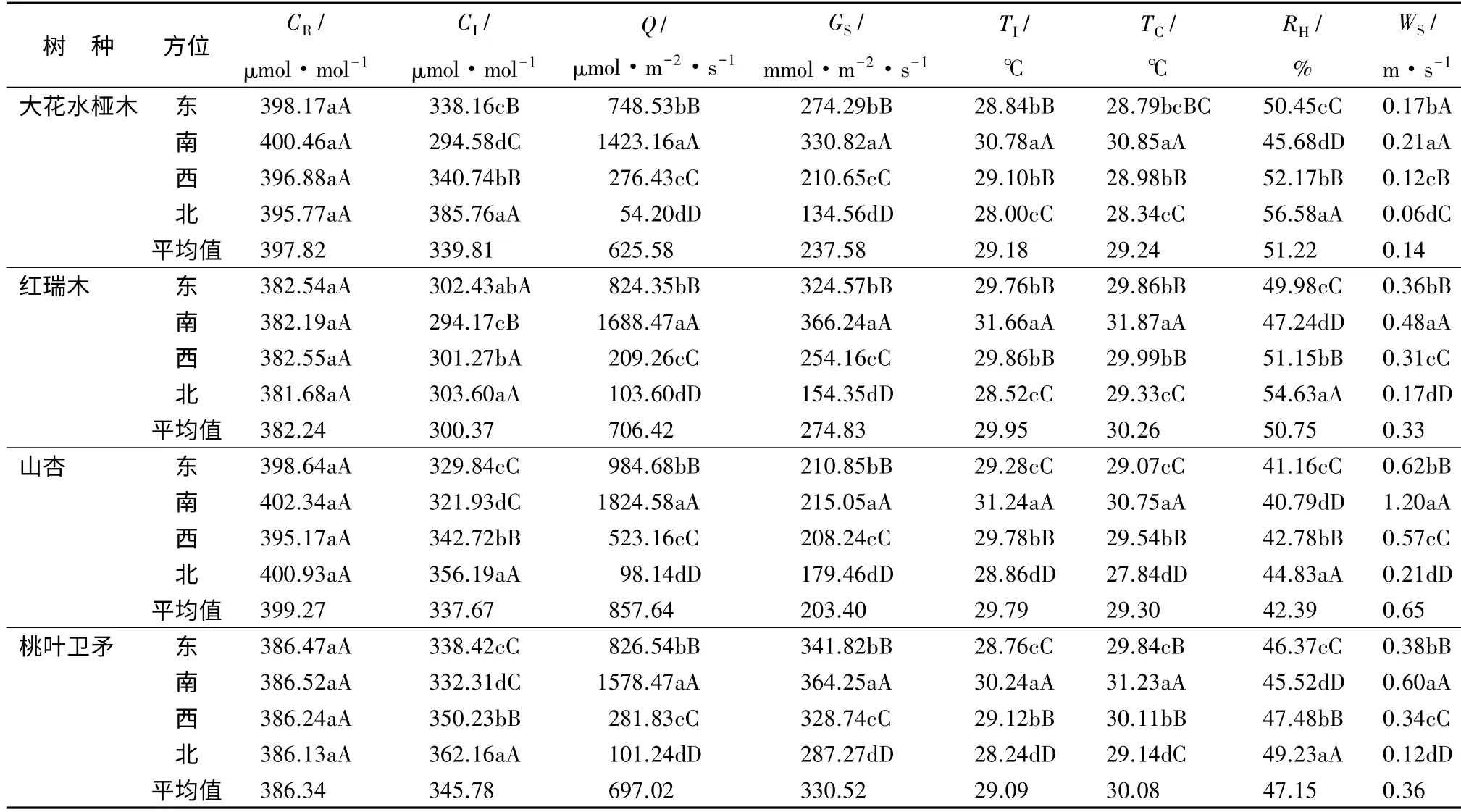
表4 不同树种树冠不同方位微环境的变化
3.4 树冠微环境及其与增湿、降温量之间的相关性
综合分析4 个树种树冠不同方位各测定部位CR、Q、TI、TC、RH和WS6 个微环境因子,以及CI和GS与增湿、降温量之间的相关性可以得出,Q、GS和WS与增湿、降温量之间呈极显著正相关,相关系数达0.97 以上;CI和RH与增湿、降温量之间分别呈显著和极显著的负相关,相关系数分别达到-0.93和-0.95;CR、TI和TC与增湿、降温量之间的相关性不明显(表5)。
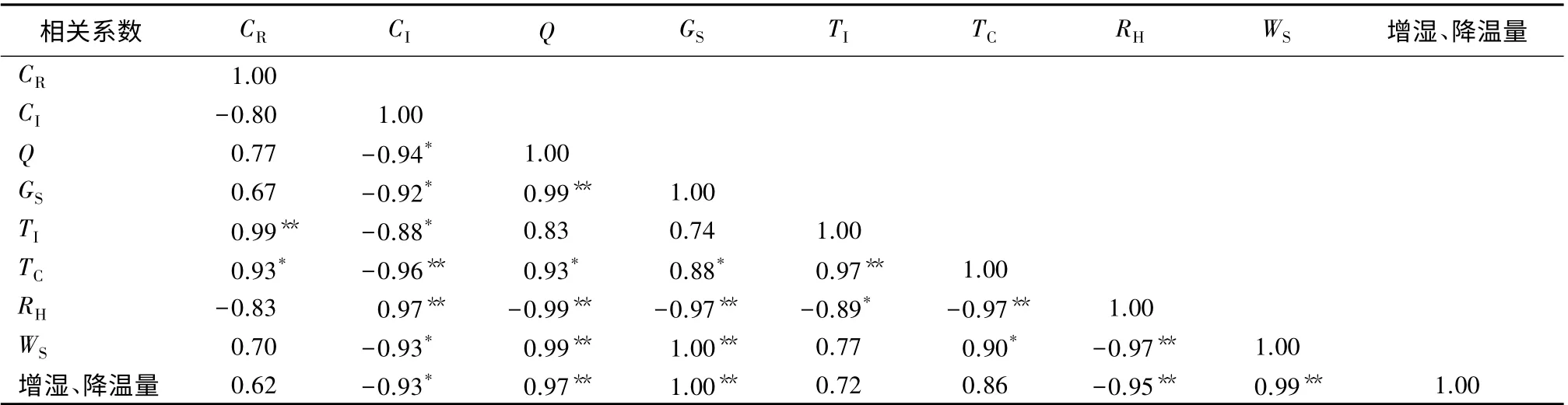
表5 树冠微环境及其与增湿、降温量之间的相关性
各测定因子之间的相关性,Q、GS、TC和WS之间分别呈极显著或显著的正相关,RH、CI与Q、GS、TC、WS分别呈极显著或显著的负相关,而RH与CI之间呈极显著的正相关。上述结果表明,Q、GS、TC和WS对提高园林树木的增湿降温生态功能具有积极的作用。
4 结束语
人们在进行园林树木增湿降温生态功能评价时,通常取树冠向阳面外缘叶片的蒸腾速率来计算全树增湿、降温量[10-14]。由于树冠不同部位环境条件的不同,导致树冠由外至内蒸腾速率依次降低,据此,林萌等[2]提出树冠外围南侧外层是园林树木单位叶面积的增湿、降温量评价的合理取样部位。本研究从树冠不同方位及其不同部位单位叶面积增湿、降温量的变化,均以外层与各方位及其不同部位的平均值比较接近,进一步验证了林萌等[2]提出取样部位的合理性。由于树冠不同方位和部位的微环境,以及不同微环境与叶片单位面积增湿、降温量的相关性的不同,会显著影响园林树木增湿降温生态功能评价科学性。如树冠不同方位和不同部位微环境的变化与其平均值比较,在测定的8 个因子中,除CR在树冠不同部位以及不同方位间基本相同外,其他因子均以树冠西侧和外层更接近平均值,树冠南侧和外层极显著高于或低于平均值。这一结果为进一步证明树冠西侧外层作为园林树木增湿降温生态功能评价的合理性提供了佐证。
以树冠西侧外层为园林树木增湿降温生态功能评价的取样部位,4 种试材的单位叶面积增湿、降温量呈现为乔木型大于灌木型,这与李辉等[15]的报道相同。但树冠表层至内层单位叶面积增湿、降温量的下降幅度上,则表现为灌木型大于乔木型,树冠不同部位单位叶面积增湿、降温量的变化幅度大于不同方位,这与树冠微环境的变化相一致,这可能是导致上述变化的重要原因之一。另一方面,可能是由乔、灌木的冠幅、干高以及投影系数的不同导致的。此外,树冠不同部位微环境的不同也可能导致叶片结构等发生变化,进而影响单位叶面积增湿、降温量。
在不同方位各测定部位CR、Q、TI、TC、RH和WS微环境因子,以及CI和GS与增湿、降温量之间,Q、GS、WS与增湿、降温量之间呈极显著的正相关,这与冷平生等[3]、王华田等[4]报道相同;CI和RH与增湿、降温量之间分别呈显著和极显著的负相关。同时,各测定因子之间又相互影响,共同制约园林树木增湿降温生态功能的强弱。上述结果表明Q、GS和WS对提高园林树木的增湿降温生态功能具有积极的作用,CI和RH通过影响Q、GS和WS的水平而显著降低园林树木增湿、降温量。因此,可以通过适宜的栽植密度和冠层结构提高园林树木增湿降温生态功能。
[1] 张大鹏.叶幕微气候及其调控生物学研究中的基本概念和内涵[J].葡萄栽培与酿酒,1993,65(2):1-4.
[2] 林萌,郭太君,代新竹.园林树木增湿降温生态功能评价方法的研究[J].生态学报,2014,34(19):269-278.
[3] 冷平生,杨晓红,胡悦,等.5 种园林树木的光合和蒸腾特性的研究[J].北京农学院学报,2000,15(4):16-17.
[4] 王华田,马履一.利用热扩式边材液流探针测定树木整株蒸腾耗水量的研究[J].植物生态学报,2002,26(6):661-667.
[5] 刘娟.轮台白杏树冠微气候与生长发育和产量品质的关系[D].乌鲁木齐:新疆农业大学,2013.
[6] 杨振伟,周延文,付友.富士苹果不同冠形微气候特征与果品质量关系的研究[J].应用生态学报,1998,9(5):533-537.
[7] 张海成,苏淑钗,雷恒久,等.板栗叶幕微气候与品质的关系[J].果树学报,2010,27(4):537-541.
[8] Hampson C R,Quamme H A,Brownlee R T. Canopy growth,yield,and fruit quality of‘Royal Gala’apple trees grown for eight years in five tree training systems[J]. The American Society for Horticultural Science,2002,37(4):627-631.
[9] 赵萱,李海梅.11 种地被植物固碳释氧与降温增湿效益研究[J].江西农业学报,2009,21(1):44-47.
[10] 廖荣,崔洁,卓丽环,等.成都市32 种立体绿化植物降温增湿效应比较研究[J].江苏农业科学,2012,40(6):178-182.
[11] 王忠君.福州国家森林公园生态效益与自然环境旅游适宜性评价研究[D].北京:北京林业大学,2004.
[12] 汪成忠.上海八种园林树木生态功能比较研究[D].哈尔滨:东北林业大学,2009.
[13] 陈少鹏,庄倩倩,郭太君,等.长春市园林树木固碳释氧与增湿降温效应研究[J].湖北农业科学,2012,51(4):750-756.
[14] 陈娟,史红文,廖建雄,等.武汉市春季49 种园林植物的光合和蒸腾特性研究[J].河南大学学报:自然科学版,2013,43(1):62-68.
[15] 李辉,赵卫智,古润泽,等.居住区不同类型绿地释氧固碳及降温增湿作用[J].环境科学,1999,20(6):41-44.
