短期暴露于城市污水处理厂尾水的文蛤抗氧化酶响应
2014-04-26苏恩萍孟范平付文超李永富中国海洋大学海洋环境与生态教育部重点实验室山东青岛266100
苏恩萍,孟范平,孙 婷,付文超,王 群,李永富,胡 恒 (中国海洋大学海洋环境与生态教育部重点实验室,山东 青岛 266100)
短期暴露于城市污水处理厂尾水的文蛤抗氧化酶响应
苏恩萍,孟范平*,孙 婷,付文超,王 群,李永富,胡 恒 (中国海洋大学海洋环境与生态教育部重点实验室,山东 青岛 266100)
为研究城市污水处理厂(STPs)尾水是否会引起海洋生物的氧化逆境,将取自青岛市李村河污水处理厂的尾水用海水稀释到不同浓度(0%、1%、5%、10%、20% V/V), 对文蛤(Meretrix meretrix)暴露培养15d, 分析其内脏中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)、谷胱甘肽还原酶(GR)的活性. 结果表明:SOD活性随尾水暴露时间延长基本表现为“抛物线型”的响应特征,体积比10%、20%的尾水暴露6~12d期间,文蛤体内SOD活性均受到诱导,最大增幅为15.88%;CAT和GPx对尾水的主要响应特征是抑制效应,分别在暴露9d、12d时最为明显,最大抑制率分别为64.38%和52.39%;较高浓度(20%)的尾水能够持续抑制GR活性,而其他处理组中,GR对尾水表现为“抛物线型”的响应特征,即:暴露3d或6d时活性增加,而后显著降低.GR活性的最大诱导出现在暴露3d和6d时,分别比对照组上升81.47%和80.91%.研究结果进一步表明STPs不能有效降解有害化学物质以消除尾水的毒性, 文蛤内脏SOD、CAT、GPx和GR活性能够敏感指示尾水诱导的氧化逆境.
城市污水处理厂;尾水;文蛤;抗氧化酶
双壳类动物由于其分布广泛、营固着生活及其高富集性、能抵抗高浓度化学污染并能从周围环境蓄积大量外源性物质等特点,现已被广泛用作海洋环境质量的指示生物[1-3].文蛤(Meretriχ meretriχ)又名花蛤,属软体动物门、双壳纲、真瓣鳃目、帘蛤科、文蛤属.生活在有淡水注入的内湾及河口附近的细沙质海滩,是我国沿海滩涂养殖的主要经济贝类之一.
城市污水处理厂(STPs)尾水的排放造成重金属、表面活性剂、氯代烃、多环芳烃(PAHs)、某些药物和内分泌干扰物等微量污染物进入受纳水体中,成为水环境中化学污染物的重要来源.而现有的污水二级处理工艺以及先进处理技术(高负荷活性污泥法等)均无法对其进行有效去除[4-5].大量研究表明:过渡金属、PAHs、有机氯和有机磷农药、多氯联苯和其他异型物质都能对生物体产生氧化压力,并引起各种有害影响,如对膜脂、DNA和蛋白质产生损伤等[6].损伤程度取决于生物细胞中具有重要功能的抗氧化酶在氧化逆境条件下的可诱导性[7].任何暴露于污染物的生物体内经常会出现这种重要的逆境响应,因此,包括抗氧化酶在内的抗氧化响应常被用作生态毒理学研究中诊断环境污染对生物体不利影响的生物标志物[8].抗氧化酶(Antioxidant enzyme)是超氧化物歧化酶(SOD)、谷胱甘肽还原酶(GR)、谷胱甘肽过氧化物酶(GPχ)、过氧化氢酶(CAT)等的统称.一旦在体内形成过氧化物(该物质能引起氧化压力),这些抗氧化酶即刻发挥功效,利用氧化还原作用将过氧化物转换为毒害较低或无害的物质.目前国外许多国家已经开始利用抗氧化酶评价 STPs尾水对海水的生态效应
[9-10],而国内的研究相对较少,很大程度上限制了尾水对海水污染影响评价的发展.
青岛目前已有五家污水处理厂,日处理污水近百万 t.这些污水经二级处理后大多排海,能否造成海洋生物抗氧化酶活性的变化尚无相关资料加以判断.本研究以青岛海域常见的双壳类动物文蛤为研究对象,以青岛李村河污水处理厂的尾水为实验材料,配制尾水体积比(EVR)为0%、1%、5%、10%、20%的尾水-海水混合液,对文蛤连续暴露培养15d,定期采样测定内脏中 4种抗氧化酶活性随尾水体积比和暴露时间的变化情况,以探讨尾水对双壳类动物抗氧化防御系统的可能影响,为今后进行尾水的海洋生态毒性效应评价提供依据.
1 材料与方法
1.1 尾水-海水混合液配制及实验动物驯养
天然清洁海水:取自青岛市石老人海域(pH值7.90±0.02、盐度32‰).
尾水:采自青岛李村河污水处理厂,处理工艺为 A2/O(厌氧-缺氧-好氧活性污泥法)+ MBBR(膜生物反应器).每隔6h采集1次,将24h内采集的 4个尾水样品等体积混合.运回实验室后,于4℃保存,当天使用.
尾水与海水混合液:将新鲜采集的尾水静置沉淀1h,取上清液(盐度约为1‰)与天然清洁海水进行混合,考虑到供试动物文蛤正常生长代谢所需的盐度范围为21‰~36‰[11],为避免尾水过量混入海水后引起盐度低于21‰的情况出现进而干扰文蛤抗氧化酶对尾水毒性的真实响应,暴露实验中采用的最大尾水体积比(EVR)为20%,并依次设置0%、1%、5%、10%、20%五个梯度.依次向各体积比的混合液中加入相应量的海水晶(山东潍坊市海佳海水晶厂以纯净海水为原料生产),溶解混均,使各体积比处理组的盐度均保持为32‰.
供试文蛤:平均壳长为 40.1mm,购自水质清洁的青岛市崂山湾大管岛海域养殖区,于冰上保存运输.挑选健康、大小基本一致的蛤仔,在温度(15±1)℃下,用天然清洁海水驯养7d.期间每天更换新鲜的天然清洁海水,定时定量投加实验室保种培养的海水小球藻(Chlorella pacifica)作为文蛤的饵料,投加密度为 1.3×107个/(L⋅d),同时利用空气泵持续充氧,使DO保持在(6±0.5)mg/L.
1.2 暴露培养
挑选健康、反应灵敏、大小一致的文蛤随机投放于15个10L玻璃缸中,每个玻璃缸投放90只,加入相应体积比的尾水-海水混合液(每个体积比设3个平行),在上述温度条件下连续培养15d,每24h 更换1次相应体积比的混合液.每天定时定量投入实验室保种培养的海水小球藻(Chlorella pacifica)作为文蛤的饵料,投加密度为1.3×107个/(L⋅d).
对于每个尾水暴露实验组或空白对照组,在培养开始时以及开始后每隔 3d,随机取出 30只动物个体(每个处理组的3个玻璃缸中各随机取出10只),立即在冰上解剖,摘取内脏,切细混合均匀,于-80℃保存,用于抗氧化酶分析.
1.3 生物标志物分析
称取一定量的内脏,采用高鹰[12]的方法制备提取液,按质量体积比 1:4加入 pH7.8的Tris-HCl(0.02mol/L)缓冲溶液进行匀浆,然后,在转速10000r/min、4℃下离心15min,取上清液测定SOD、CAT、GPχ、GR活性.
基于酶试剂盒在测定抗氧化酶活性方面的快速、简便、精密度高、易于标准化等优点,SOD、CAT、GPχ、GR及上清液中蛋白质含量均采用南京建成生物工程研究所生产的试剂盒测定,单位分别为U/mg蛋白质、U/g蛋白质、U/mg蛋白质、U/g蛋白质、g/L.
1.4 数据统计分析
抗氧化酶测定结果表示:对于每个体积比的尾水暴露组或空白对照组,各暴露时间制备的内脏样品中每种抗氧化酶均平行测定3次,以(平均值 Xi±标准差 SD)表示该尾水暴露组(或空白组)的内脏中该抗氧化酶的活性.
抗氧化酶测定结果的差异显著性判断:用单因素方差分析中 Games-Howell(方差非齐性)或Tukey(方差齐性)检验方法对暴露培养中每种抗氧化酶活性随尾水体积比、暴露时间的变化差异性检验.统计显著性水平为P<0.05.数据统计分析使用SPSS 18.0软件.
相关性分析:采用双变量 Pearson相关性分析,研究呈单向响应的生物标志物测定值与尾水体积比的关系,统计显著性水平为 P<0.05和P<0.01.
2 结果与讨论
在城市化高度发展和人口密集的沿海地区,污染物随着STPs出水(或尾水)的排放而进入受纳水体中,成为重要污染源[13-14].目前,虽有研究发现作为尾水化学组成成分的壬基酚(NP)、有机氯农药和PAHs等化学物质能够通过过氧化作用产生生物毒性[15-18],但是有关 STPs尾水对双壳类动物氧化逆境因子影响的系统研究尚不多见.
2.1 尾水-海水混合暴露对文蛤内脏SOD活性的影响
图1为暴露于尾水-海水混合液后文蛤的内脏SOD活性随尾水体积比(EVR)的变化情况.暴露前的文蛤内脏SOD活性为51.57U/mg蛋白质,在不含尾水的海水(对照组)中培养期间,SOD活性保持在54.41~57.75U/mg蛋白质之间.在尾水-海水混合体系暴露3d后,文蛤内脏SOD活性仅在EVR为10%的处理组中被诱导(P<0.05),比对照组提高5.64%;暴露6d后,有3个处理组的SOD活性显著高于对照组的水平,最大增幅为15.88%;暴露9d后,所设置的4个尾水体积比处理均诱导了内脏SOD活性,最大增幅出现在EVR1%的处理组,为12.98%.随后,尾水对SOD的诱导逐渐减弱,暴露12d时,只有体积比为10%、20%的两个处理组表现出高于对照组的SOD活性,且增幅分别仅为6.87%和5.66%;至暴露15d时,除了EVR为 1%的处理组显著高于对照组外,其他处理组的SOD活性均低于对照组(EVR5%、10%处理组的降幅分别为 9.63%和 7.67%,P<0.05)或与对照组差异不大(EVR20%处理组,P>0.05).
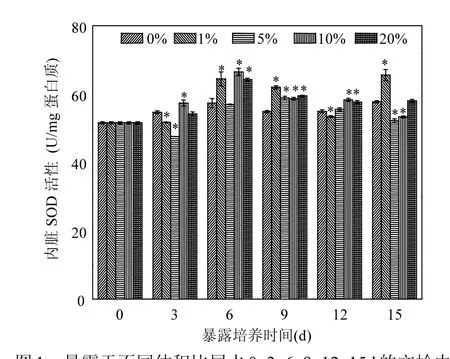
图1 暴露于不同体积比尾水0, 3, 6, 9, 12, 15d的文蛤内脏SOD活性变化Fig.1 Variation of SOD activities in the viscera of mussels exposed to serial dilutions of STP effluent on days 0, 3, 6, 9, 12and 15*表示在P<0.05水平上与对照组差异显著,下同
SOD主要存在于细胞质内以及线粒体内外膜之间,其作用是催化超氧阴离子自由基发生歧化反应生成 H2O2和.有关海水重金属
[20]、NP[21]等对双壳类动物 SOD活性影响的许多研究报道:随着暴露时间延长或者浓度增加,诱导幅度逐渐变小,甚至出现抑制,呈现“抛物线型”的剂量-效应关系.本研究发现(图 1),各体积比的尾水处理组中,随着暴露时间延长,SOD活性基本表现为“抛物线型”的响应趋势,只是EVR1%处理组在暴露末期仍有较大幅度诱导,使剂量效应趋势与其他处理组有所不同.SOD的诱导效应多出现在第6、9、12d,而暴露初期和后期的活性较低,个别处理组甚至受到一定程度抑制.在同样的暴露时间下,由于尾水中污染成份之间的复杂作用,SOD活性对不同剂量(体积比)尾水并未体现出“抛物线型”响应特征,仅在暴露15d时有相对明显的低促高抑现象.总体上,在为期15d的暴露期间,文蛤SOD活性对尾水的响应以诱导为主,活性最大增幅为 15.88%,出现在暴露第6d的EVR10%处理组.SOD活性的上升,意味着尾水中的污染成分进入蛤体后能够产生一定量的⋅,需要较高SOD活性予以清除.
2.2 尾水-海水混合暴露对文蛤内脏CAT活性的影响

图2 暴露于不同体积比尾水0, 3, 6, 9, 12, 15d的文蛤内脏CAT活性变化Fig.2 Variation of CAT activities in the viscera of mussels exposed to serial dilutions of STP effluent on days 0, 3, 6, 9, 12and 15
由图 2可知,暴露前文蛤内脏过氧化氢酶(CAT)活性为80.77U/g蛋白质,随后的培养期间,对照组的CAT活性变化较小(78.68~84.51U/g蛋白质).经尾水-海水混合体系暴露3d后,各处理组的文蛤内脏 CAT活性均受到明显抑制(P<0.05),与对照组相比,EVR 1%、5%、10%、20%的处理组中CAT活性分别降低44.28%、45.57%、23.41%和39.08%;暴露6d时,各处理组的CAT活性均出现回升,EVR 10%、20%与对照组无显著差异(P>0.05);继续延长暴露时间到培养结束,各处理组的CAT活性无一例外地再次受到明显抑制,且抑制程度基本上呈现随EVR增大而提高的趋势,其中,第 9d和 12d的活性降幅最大, EVR20%的处理组中CAT活性分别降为对照组的 53.38%和 35.62%.暴露 15d时,各处理组的CAT活性虽然有所上升,但仍然显著低于对照组.
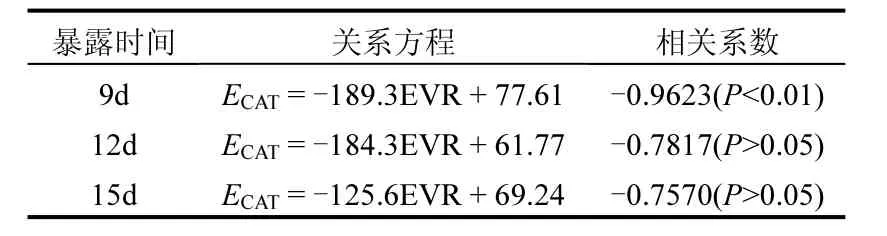
表1 暴露于尾水后文蛤内脏CAT活性与EVR的相关性分析Table 1 Correlative relationship between EVR and CAT activities in the viscera of Meretriχ meretriχ exposed to effluent
CAT是一类末端氧化酶,主要功能是将体内的H2O2分解成O2和H2O,减轻H2O2对细胞的氧化损伤.有学者认为,CAT通常是最早受到诱导的抗氧化酶之一[22-23].但是,以往文献中,关于双壳类动物CAT对各种污染物响应特征的研究结果各不相同;较低浓度重金属诱导双壳类CAT活性的情况十分少见,随着海水重金属浓度的增加或暴露时间延长,双壳类CAT活性一般会受到较大抑制[24-28].但是,关于有机污染物的研究发现[18],暴露于PAHs(蒽、芘)和有机氯农药(六六六、艾氏剂、狄氏剂、p,p′-DDT)的翡翠贻贝(Perna viridis)内脏 CAT活性均受到显著诱导,而且这种情况只发生于暴露开始后一周内.本研究的尾水暴露期间,未观察到任何处理组的文蛤CAT活性被诱导,相反,CAT活性受抑制的现象非常普遍(只有第6d的EVR10%、EVR20%处理组与同期对照组差异不显著).暴露初期的CAT活性抑制程度与EVR之间无明显规律;而在暴露9~15d时,高EVR处理组的CAT活性一般受到较大抑制,表现出强烈的剂量效应关系.这种响应特征不同于前人研究中有机污染物引起 CAT活性明显上升的情况,而更类似于大多数种类的重金属对 CAT的抑制效应.相关性分析表明,暴露9d时的文蛤内脏CAT活性与尾水EVR具有极显著(P<0.01)的负相关性,而在其他暴露时间两者均不存在显著相关性(P>0.05).文蛤CAT在某一暴露时段对尾水的显著相关性响应特征,对于尾水排放海域污染程度的评价分级是有利的.
2.3 尾水-海水混合暴露对文蛤内脏GPχ活性的影响

图3 暴露于不同体积比尾水0, 3, 6, 9, 12, 15d的文蛤内脏GPχ活性变化Fig.3 Variation of GPχ activities in the viscera of mussels exposed to serial dilutions of STP effluent on days 0, 3, 6, 9, 12and 15
正式培养之前,文蛤内脏的谷胱甘肽过氧化物酶(GPχ)活性为10.08U/mg蛋白质.虽然培养期间活性稍有提高,但波动幅度不大(11.46~13.87U/mg蛋白质).与 CAT类似,大多数处理组的GPχ活性在暴露3d后即受到显著抑制,相对于同期的对照组,降幅在8.29%~15.65%,高EVR处理组的抑制率较高;暴露时间延长至 6d、9d时,除了EVR10%的处理组持续对GPχ活性产生抑制(8.47%、9.34%)外,其他处理组的GPχ活性大多回升到与对照组差异不显著的水平,暴露9d时EVR1%处理组的GPχ活性(13.58U/mg 蛋白质)甚至比同期的对照组(11.46U/mg 蛋白质)增加18.50%,即受到明显诱导;但是,当暴露时间继续延长时,所有处理组的GPχ活性又受到显著抑制,其中,以 EVR5%的处理组降幅最大(暴露 12d、15d时分别为52.39%和34.29%).
GPχ能够通过催化还原型谷胱甘肽(GSH)与H2O2或氢过氧化物反应(ROOH+2GSH→ROH+ H2O+GSSG),有效阻止自由基引起的氧化损伤,而 GSH 自身被氧化为氧化型谷胱甘肽(GSSG)[25].可见,其与 CAT具有相似的抗氧化功能,即具有共同的催化底物.图3的结果显示,尾水-海水混合体系培养初期和后期,各处理组的文蛤内脏GPχ活性均普遍受到抑制,且以后期受到的抑制程度较高,虽然暴露中期(第6d和第9d)的GPχ活性有所恢复,EVR1%处理组在第9d的GPχ活性甚至高于同期对照组,但是其在培养结束时均显著低于对照组.化学污染物对海洋动物 GPχ活性的抑制作用常见于国内外室内和现场调查的文献中.Richardson等[18]分别采用含PAHs、有机氯农药的海水培养翡翠贻贝4周,发现鳃GPχ活性与软组织中芘、萤蒽(P<0.001)、蒽、苯并芘(P<0.01)、六六六、狄氏剂、p,p’-DDT(P<0.05)等污染物含量之间均存在显著负相关性.孟范平等(2011)[20]综合国内外暴露于海水重金属后双壳类抗氧化酶响应的研究结果认为,重金属离子(Cu2+、Pb2+、Hg2+)的单一暴露一般会抑制双壳类鳃和消化腺中GPχ活性.Rocher等[26]对法国塞纳河口及塞纳湾不同站位的采样分析表明,斑马贻贝(Dreissena polymorpha)和紫贻贝(Mytilus edulis)的鳃 GPχ活性与软组织中重金属总含量呈显著负相关(r= -0.73,P<0.05).另外,本研究将图2、图3中5个暴露时刻(不含零时刻)测定的25个活性数值进行相关性分析发现,CAT和GPχ活性之间存在显著正相关性(r=0.558,P<0.01),表明尾水暴露过程中,两者活性的变化具有一定同步性.暴露初期尾水对两种酶的抑制效应可能是因为刚接触尾水不久,文蛤对进入体内的污染成分难以适应所致;后期的抑制效应则是因为较长时间的尾水暴露后,有机、无机污染成分在体内蓄积而产生过多 H2O2或氢过氧化物等活性氧,攻击酶分子而引起两种抗氧化酶的活性降低,此时的内脏已处于一定的氧化逆境状态.
2.4 尾水-海水混合暴露对文蛤内脏GR活性的影响
由图4可见,暴露培养之前的文蛤谷胱甘肽还原酶(GR)活性为13.17U/g蛋白质,实验期间对照组的GR活性基本稳定在13.28~15.00U/g蛋白质之间.暴露3d时,EVR 1%的处理组GR活性受尾水作用而显著提升,比同期对照组增加81.47% (P<0.05),但是EVR 5%、10%的尾水处理未显著影响到GR活性(P>0.05),EVR 20%的尾水处理则使GR活性降为8.49U/g 蛋白质.暴露6d时,除了EVR 20%处理组的GR活性(3.56U/g 蛋白质)仍受到较大抑制外,其余处理组的 GR活性均出现上扬,增幅在31.54%~80.91%.暴露9d后,多数处理组的GR活性受到一定程度抑制,其中,EVR20%的处理组活性下降幅度最大,第9,12,15d时的GR活性分别比同期的对照组下降69.72%、47.97%和61.25%.

图4 暴露于不同体积比尾水0, 3, 6, 9, 12, 15d的文蛤内脏GR活性变化Fig.4 Variation of GR activities in the viscera of mussels exposed to serial dilutions of STP effluent on days 0, 3, 6, 9, 12and 15
GR能够将GSSG再生为GSH,以保持氧化逆境下适宜的GSH/GSSG比值,维持细胞内一定的-SH水平,因此,逆境条件下GR的诱导十分重要,其活性已应用于某些海域现场监测评价中,被认为是一种较为可靠的氧化逆境标志物[27].本研究中,当EVR达到20%时,无论暴露时间长短,GR活性均受到抑制.在其他处理组中,GR活性在尾水暴露初期(3d和 6d)多受到诱导(少数与同期对照组差异不大),而暴露后期的大多数处理组中则观察到内脏GR活性显著降低(P<0.05),只有少数处理组的活性与同期对照组差异不显著.因此认为,文蛤内脏GR对EVR≤10%的尾水的响应符合“抛物线型”的时间效应特征,但是在剂量效应上,这种响应特征仅见于暴露初期(第3d和第6d).低浓度污染物或暴露初期对双壳类动物GR活性的刺激性,已在室内研究[28]和现场调查[29-31]中得到证实.暴露初期文蛤内脏中较高的 GR活性,反映了GSSG向 GSH的快速转化,有助于维持机体内GSH含量稳定.而高剂量污染物(或高EVR尾水)以及暴露后期的GR活性受抑,则意味着细胞正常代谢所需的 GSH/GSSG比例难以维持,导致GSSG积累,使文蛤内脏细胞处于氧化胁迫状态.
3 结论
3.1 SOD活性随尾水暴露时间延长基本表现为“抛物线型”的响应特征.EVR10%、20%的尾水暴露6~12d期间,SOD活性均受到诱导,最大增幅为15.88%.
3.2 CAT和GPχ对尾水的主要响应特征是抑制效应,分别在暴露9d、12d时最为明显,最大抑制率分别为64.38%和52.39%.
3.3 EVR20%的尾水能够持续抑制 GR活性.EVR≤10%的处理组中,GR对尾水基本表现为“抛物线型”的时间效应特征,而同样形式的剂量效应只出现在暴露初期.GR活性的最大诱导出现在暴露3d和6d时,分别比对照组上升81.47%和80.91%.
[1] Fernandez B, Albentosa M. Integrated assessment of water quality of the Costa da Morte (Galicia, NW Spain)by means of mussel chemical, biochemical and physiological parameters[J]. Ecotoxicology, 2010,19:735-750.
[2] Vethaak A D, Lahr J, Schrap M, et al. An integrated assessment of estrogenic contamination and biological effects in the aquatic environment of the Netherlands [J]. Chemosphere, 2005,59:511-524.
[3] Li X M, Zhang Q H, Dai J V. Pesticide contamination profiles of water, sediment and aquatic organisms in the effluent of Gaobeidian wastewater treatment plant [J]. Chemosphere, 2008, 72:1145-1151.
[4] Watkinson A J, Murby E J, Kolpin D W. The occurrence of antibiotics in an urban watershed: From wastewater to drinking water [J]. Sci. Total Environ., 2009,407:2711-2723.
[5] Bolong N, Ismail A F, Salim M R. A review of the effects of emerging contaminants in wastewater and options for their removal [J]. Desalination, 2009,239:229-246.
[6] 王丽平,郑炳辉,孟 伟.环境污染物对水生生物产生氧化压力的分子生物标志物 [J]. 生态学报, 2007,27(1):380-388.
[7] Di Giulio R T, Washburn P C, Wenning R J, et al. Biochemical responses in aquatic animals: a review of determinants of oxidative stress [J]. Environ. Toxicology and Chemistry, 1989,8:1103-1123.
[8] Letendre J, Chouquet B, Manduzio H, et al. Tidal height influences the levels of enzymatic antioxidant defences in Mytilus edulis [J]. Marine Environmental Research, 2009,67:69-74.
[9] Zhi-Hua L, Vladimir Z, Jan T. Evaluating environmental impact of STPs situated on streams in the Czech Republic: An integrated approach to biomonitoring the aquatic environment [J]. Water Research, 2011,45:1403-1413.
[10] Steven B, Christopher H, Benat Z. Integrated biomarker assessment of the effects exerted by treated produced water from an onshore natural gas processing in the North Sea on the mussel Mytilus edulis [J]. Marine Pollution Bulletin, 2011,62:327-339.
[11] Tang B J, Liu B Z, Yang H S, et al. Oxygen consumption and ammonia-N excretion of Meretrix meretrix in different temperature and salinity [J]. Chinese Journal of Oceanology and Limnology, 2005,23(4):469-474.
[12] 高 鹰.海水铜铅及环境因子对菲律宾蛤仔生物标志物的影响研究 [D]. 青岛:中国海洋大学, 2011,22-23.
[13] Watkinson A J, Murby E J, Kolpin D W, et al. The occurrence of antibiotics in an urban watershed: From wastewater to drinking water [J]. Sci. Total Environ., 2009,407: 2711-2723.
[14] Bolong N, Ismail A F, Salim M R, et al. A review of the effects of emerging contaminants in wastewater and options for their removal [J]. Desalination, 2009,239:229-246.
[15] 高永刚,李正炎.壬基酚对栉孔扇贝组织抗氧化酶活性的影响[J]. 中国海洋大学学报, 2006,36(S):135-138.
[16] Chen F, Gao J, Zhou Q. Toxicity assessment of simulated urban runoff containing polycyclic musks and cadmium in Carassius auratus using oxidative stress biomarkers [J]. Environmental Pollution, 2012,162:91-97.
[17] Gagne F, Blaise C, Andre C. Occurrence of pharmaceutical products in a municipal effluent and toxicity to rainbow trout (Oncorhynchus mykiss) hepatocytes [J]. Ecotoxicology and Environmental Safety, 2006,64:329-336.
[18] Richardson B J, Mak E, Luca-Abbot S B De, et al. Antioxidant responses to polycyclic aromatic hydrocarbons and organochlorine pesticides in green-lipped mussels (Perna viridis): Do mussels“integrate” biomarker responses? [J]. Marine Pollution Bulletin, 2008,57:503-514.
[19] Manduzio H, Monsinjon T, Rocher B, et al. Characterization of an inducible isoform of the Cu/Zn superoxide dismutase in the mussel Mytilus edulis [J]. Aquatic Toxicology, 2003,6(64):73-83.
[20] 孟范平,高 鹰,赵顺顺,等.双壳类分子生物标志物对海水重金属的响应评述 [J]. 中国海洋大学学报, 2011,41(5):100-109.
[21] Cossu C, Doyotte A, Jacquin M C, et al. Glutathione reductase, selenium-dependent glutathione peroxidase, glutathione levels, and lipid peroxidation in freshwater bivalves, Unio tumidus, as biomarkers of aquatic contamination in field studies [J]. Ecotoxicology and Environmental Safety, 1997,28:122-131.
[22] Regoli F, Gorbi S, Frenzilli G, et al. Oxidative stress in ecotoxicology: from the analysis of individual antioxidants to a more integrated approach [J]. Marine Environment Research, 2002,54:419-423.
[23] 赵元凤,吕景才,宋晓阳.海洋污染对毛蚶过氧化氢酶影响研究[J]. 环境科学学报, 2002,22(4):534-536.
[24] 王 凡,赵元凤,吕景才,等.铜污染对扇贝内脏团抗氧化酶活性的影响 [J]. 水产科学, 2008,27(12):622-624.
[25] Regoli F, Principato G. Glutathione, glutathione-dependent and antioxidant enzymes in mussel, Mytilus galloprovincialis, exposed to metals under field and laboratory conditions: implications for the use of biochemical biomarkers [J]. Aquatic Toxicology, 1995,31:143-164. [26] Rocher B, Goff J L, Peluhet L, et al. Genotoxicant accumulation and cellular defence activation in bivalves chronically exposed to waterborne contaminants from the Seine River [J]. Aquatic Toxicology, 2006,79:65-77.
[27] Meyer J N, Smith J D, Winston G W, et al. Antioxidant defenses in killifish (Fundulus heteroclitus) exposed to contaminated sediments and model prooxidants: short-term and heritable responses [J]. Aquatic Toxicology. 2003,65(4):377-395.
[28] Verlecar X N, Jena K B, Chainy G B N. Modulation of antioxidant defences in digestive gland of Perna viridis (L.), on mercury exposures [J]. Chemosphere, 2008,71:1977-1985.
[29] Torres M A, Testa C P, Gaspari C. Oxidative stress in the mussel Mytella guyanensis from polluted mangroves on Santa Catarina Island, Brazil [J]. Marine Pollution Bulletin, 2002,44:923-932.
[30] Porte C, Biosca X, Sole M, et al. The integrated use of chemical analysis, cytochrome P450 and stress proteins in mussels to assess pollution along the Galician coast (NW Spain) [J]. Environmental Pollution, 2001,112:261-268.
[31] Cheung C C C, Zheng G J, Li A M Y, et al. Relationships between tissue concentrations of polycyclic aromatic hydrocarbons and antioxidative responses of marine mussels, Perna viridis [J]. Aquatic Toxicology, 2001,52:189-203.
Responses of the antioxidant enzymes in Meretriχ meretriχ exposed to effluent from municipal sewage treatment
plants for short-term.
SU En-ping, MENG Fan-ping*, SUN Ting, FU Wen-chao, WANG Qun, LI Yong-fu, HU Heng
(Key Laboratory of Marine Environment and Ecology, Ocean University of China, Ministry of Education, Qingdao 266100, China). China Environmental Science, 2014,34(2):511~517
In order to investigate if effluent from municipal sewage treatment plants (STPs) cause oxidative stress in marine organisms, Meretriχ meretriχ were exposed to effluent from the Licunhe STP in Qingdao City at various concentrations (0%, 1%, 5%, 10% and 20% V/V), diluted in seawater, for 15days. Antioxidant enzymes analyzed in the viscera of mussels were superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPχ) and glutathione reductase (GR). Results showed that the variation trend of SOD activities with exposure time exhibited a parabolic curve shape with a maximal increment of 15.88%, which was induced from day 6 to day 12 in both exposure groups (10% and 20% effluent). Both CAT and GPχ were inhibited by the effluent with the highest inhibition 64.38% and 52.39%, at the day 9 and day 12, respectively. At higher effluent concentration (20%), GR activity was inhibited constantly during the exposure period. However, GR activities responded in a parabolic curve shape in other exposure groups where an initial increase was observed at day 3 or day 6 followed by a significant decrease after day 9, and the highest increment were 81.47% and 80.91%, at day 3 and day 6, respectively. Current experimental results suggest that STPs do not have the capacity to biodegrade harmful chemicals sufficiently to prevent toxic action of effluent, and SOD, CAT, GPχ and GR activities in viscera of M. meretriχ are sensitive indicators of oxidative stress induced by effluent.
municipal sewage treatment plants (STPs);effluent;Meretriχ meretriχ;antioxidant enzymes
X835
:A
:1000-6923(2014)02-0511-07
苏恩萍(1990-),女,山东招远人,中国海洋大学硕士研究生,主要从事环境生物监测技术研究.
2013-06-21
国家自然科学基金项目(41240040)
* 责任作者, 教授, fanpingm@tom.com
