剑尾鱼精巢的显微和超微结构观察
2012-08-16温茹淑曾德莉方展强
温茹淑,曾德莉,方展强
(1.嘉应学院生命科学学院,广东梅州514015;2.华南师范大学生命科学学院,广东省高等学校生态与环境科学重点实验室,广东广州510631)
剑尾鱼(Xiphophorus helleri),别名剑鱼、青剑,英文名 Swordtail.隶属于鳉形目,花鳉科(Poeciliidae),胎鳉属.原产地墨西哥、危地马拉,是一种热带观赏鱼类,其体型小,繁殖周期短,易饲养.其被开发作为水环境污染“活检测器”的理想实验鱼,在水生态毒理学研究中的应用亦逐步展开[1].
关于硬骨鱼类精巢组织学和形态学的研究已有较多的报道[2-3],多集中在卵生、体外受精类型的硬骨鱼类.对卵胎生、体内受精鱼类的研究则较少[3].剑尾鱼属卵胎生,体内受精鱼类,其鳃、肝脏和肾脏的组织结构及超微结构已有研究报道[3-6],尚未见有关其精巢组织结构研究的报道.本研究通过对雄性剑尾鱼精巢的形态学、组织学、超微结构等方面的研究,以期对剑尾鱼的繁殖生物学提供基础资料,为进一步调控鱼类的生殖活动,提高鱼类的繁殖性能和生产水平,为剑尾鱼的实验动物化提供基础理论.
1 材料与方法
1.1 实验用鱼
实验用的剑尾鱼(Xiphophorus helleri)于2009年6月取自珠江水产研究所,为同批孵化的性成熟雄性个体,体长为6.35 ±0.5 cm,体质量为1.6 ±0.6 g.
1.2 形态学观察
将麻醉的剑尾鱼置于解剖盘中,解剖观察精巢和卵巢的外形特征,对其形态特征进行描述并拍照.然后将其保存在体积分数为5%的福尔马林液(含质量分数0.65%NaCl)中,以备观察.
1.3 组织学观察
活体取精巢进行常规石蜡切片,用Bouin氏液固定24 h,经各级乙醇冲洗、脱水,经二甲苯透明,54~56℃石蜡包埋,制成5~7μm切片,H—E染色,中性树胶封片.置Olympus BX45-32P02显微镜进行显微观察,用尼康数码相机进行拍照.
1.4 超微结构观察
部分精巢样品经0.2 mol/L磷酸缓冲液(pH 7.4)漂洗,投入体积分数4%的戊二醛低温固定15 h,后置体积分数1%的锇酸固定3h,逐级乙醇脱水,环氧丙烷透明,Epon812包埋,切成40~60 nm超薄切片,醋酸双氧铀及Reynold柠檬酸铅复染,置日立H-300型透射电镜观察及拍照.
2 结果与分析
2.1 精巢的外部形态
剑尾鱼属卵胎生鱼类,70 d龄左右出现雄性性征,即尾鳍出现剑尾,雄鱼具生殖足(由臀鳍演变而来).在解剖镜下观察发现雄性剑尾鱼腹腔有剑尾鱼精巢1对,约0.8 cm,呈乳白色.位于腹腔背侧,消化道背面,肾脏腹面.左右精巢(T)紧密排列,呈2条索状结构,左右精巢前端游离于腹腔,在中部开始汇合成1条输精管(VD),开口于由臀鳍形成的生殖足.精巢其横切面呈卵圆形,左右精巢大小略有差异,精巢外被一层结缔组织膜,表面光滑,有背腹之分,背部拱起,腹部较平坦,腹面有一纵向凹陷,其中有血管分布(图版1).
2.2 精巢的组织结构
剑尾鱼精巢的横切面呈卵圆形,内部充满了精小囊(SC),精子发育的全过程在精小囊内进行(图版2).剑尾鱼精子的发育过程历经初级精原细胞、次级精原细胞、初级精母细胞、次级精母细胞、精子细胞、成熟精子和精子束.在同一精小囊中,精子的发育基本同步,但在不同精小囊中生殖细胞的发育不一致.生殖季节有不同发育阶段生精细胞构成的精小囊(图版3).在光镜下观察成熟雄鱼的精巢切片,可见处于不同发育阶段的精细胞.
精原细胞是生精细胞中体积最大的细胞,在精巢中可见到2种类型的精原细胞,包括初级精原细胞(SG1)和次级精原细胞(SG2).
初级精原细胞 细胞体积大,紧贴生殖上皮,常1~3个分散存在,呈圆形、椭圆形或梭形,核椭圆形,居中或偏于一侧,核膜清晰,细胞质几乎不染色,可见到被染成淡红色的细胞膜(图版4).
次级精原细胞 靠近生殖上皮,细胞呈圆形或椭圆形,核圆形,核膜较初级精原细胞模糊,细胞质染色也较淡,常几个聚集在一起外包以极薄的结缔组织膜组成一个较小的精小囊(图版5).
初级精母细胞 精小囊体积明显增大,初级精母细胞(SC1)体积比初级精原细胞小,细胞呈圆形,核仁消失,细胞核圆形或椭圆形,可看见细胞核内染色较深的染色体,核周围间隙呈囊泡状(图版6).
次级精母细胞 精小囊的体积达到最大,次级精母细胞(SC2)体积略小于初级精母细胞,细胞与细胞之间界限清楚,细胞多为圆形,细胞核位于一侧,呈圆形或略椭圆形,核膜较清晰(图版7).
精子细胞 精子细胞(ST)小于次级精母细胞,是各期生精细胞中体积最小的生殖细胞,精子细胞中细胞核占细胞体积的绝大部分,核物质浓缩,细胞质减少(图版8).精子细胞不再分裂.它要经过精子形成过程才能成为有功能的精子.
成熟精子 成熟精子(SP)头部呈椭圆形,并形成细长的鞭毛(图版9).精子鞭毛为典型的9+2结构,中间为轴丝.
精子束 精子呈放射状整齐排列于精小囊内,头部(↓)指向囊壁,尾部(↓)指向精小囊中央腔(图版10).
精小管内有许多精小囊,精小囊按照精子的发育阶段呈规律分布(图版11,12).从同一切面可看出,发育早期(↓)的细胞往往处于精巢的外围,而发育晚期(↓)的细胞往往处于精巢的中间部位.因此可推断,从精子发生到成熟精子的形成,精小囊是逐渐向中央腔方向移动,在迁移过程中逐渐发育的,在精子成熟过程中,精小囊慢慢演变成包裹成熟精子的精包.最后精子以精包为单位被释放到与输精管相连接的输精管中央腔(↓)中(图版13).待精子完全成熟后,精包破裂.精子被释放出来,并通过输出管排出体外.由此可说明,剑尾鱼的精巢属于小管型精巢.
2.3 精子发育的超微结构
剑尾鱼的精细胞(↓)经过发育变态后成为形态复杂的成熟精子(图版14),精巢内成熟精子呈放射状整齐而又紧密排列于精小囊内,头部朝向管壁支持细胞,并埋于支持细胞中,尾部朝向精小囊中央腔,并呈多层排列(图版15).剑尾鱼的精子分为头部、中段、尾部3个部分,成熟精子的头部细长,呈椭圆形,只有1个由核蛋白组成染色质致密的核,其外有1层质膜,精子前端没有顶体,后端有一较浅的植入窝;中段紧接精子头部,直径较精子头部略大,线粒体多,其轴丝为典型的“9+2”结构(图版16);尾部的轴丝的横切面也是典型的“9+2”双联管结构,轴丝外包绕着线粒体鞘,外侧由1层鞘膜包围,末段轴丝裸露(图版17).
3 讨论
3.1 剑尾鱼精巢形态结构的特点
硬骨鱼类的成熟精巢体积一般较小,大多呈长囊状,且成对存在,仅件黄鳝(Monopterus albus)的除外,其精巢单一,位于身体右侧,为长圆柱形[7].一般硬骨鱼类的精巢在尾端合并,呈“Y”型.但一些鱼类,如乌鳢(Ophicephalus argus)[8]、金色小沙丁鱼(sardineUa aurita)[9]精巢在后端并不合并,而是分别开口于泄殖腔.本文观察到成熟剑尾鱼的精巢呈2条紧密排列的索状结构,精巢中部汇合成一较短的输精管,开口于由臀鳍形成的生殖足,但精巢在外观上并不成“Y”型.同属于硬骨鱼的褐菖鮋(Sebastiscus marmoratus)[3]、孔 雀 鱼 (Poecilia reticulata)[10],其精巢分别呈梭形和肾形,在精巢的末端有左右2条输精管平行排列,并向鱼体后端延伸,最后2条输精管汇合,经泄殖孔开口于体外.由此可见,在卵胎生鱼类中,其精巢的形态和结构也存在显著差异.而鲇形目鱼类如长吻鮠(Leiocassis longirostris Gianther)[2]、黄颡鱼(Pelteobagrus fulvidraco Richardson)[10]等精巢往往分支,黄颡鱼精巢每侧向外伸出约26个叶状分支,叶状分支基部宽扁,向末端逐渐变细,贮精囊由左右侧生精部在近肾脏末端处合并,形成输精管,在输精管两侧也伸出约14对细长的指状分支.而长吻鮠[11]精巢分支更典型,精巢高度分支呈指状,分支之上还有分支.而鲇形目的革胡子鲇(Clarias leather)等鱼的精巢形态则仅精巢外侧缘有少量较短的齿状突起[12].
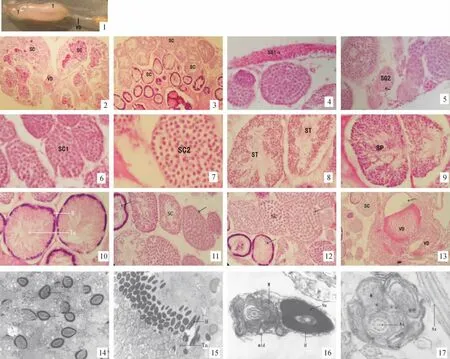
图版 剑尾鱼精巢的显微和超微结构
3.2 剑尾鱼精巢类型的划分
关于硬骨鱼类精巢类型的划分,一般根据各级生精细胞在精巢内分布的特点而分为小叶型和小管型两种.二者的主要区别在于叶型精巢的精原细胞存在于小叶的边缘处,随着发育的进行,精原细胞和精母细胞形成小囊腔隙,精子细胞和精子逐渐向小囊腔的中央移动,其成熟精子存在于小叶腔和输精小管内.管型精巢的精原细胞只分布在小管的盲端,随着发育,精子细胞和精子逐渐向输出管移动,其成熟精子只存在于靠近输精小管的部位.
本实验对剑尾鱼精巢组织的研究表明,在精小管横切面上存在一种生精细胞,并且从同一切面可看出,发育早期的细胞往往处于精巢的外围,而发育晚期的细胞往往处于精巢的中间部位,从背侧到腹侧依次为精原细胞、精母细胞、精子细胞和成熟精子,从精子发生到成熟精子的形成,精小囊逐渐向中央腔方向移动,逐渐发育,最后成熟精子被释放到与输精管相连接的输精管中央腔中,并通过输精管排出体外.故认为剑尾鱼精巢属于管型结构.大多数硬骨鱼类的精巢为叶型精巢,属管型精巢的鱼类很少.
3.3 精子发生和形成的特点
剑尾鱼精子的发生有如下特点:
(1)同大多数硬骨鱼类一样,剑尾鱼精巢内存在2种类型的精原细胞,即初级精母细胞和次级精母细胞或称之为A型精原细胞和B型精原细胞[13].两者的主要差异表现在核的形态和位置,核质的染色程度以及细胞是否存在于精小囊中.对于鱼类的精原细胞,大多数学者认为有2种类型,在性成熟的雄鱼精巢中初级精原细胞是一种干细胞,只能进行有丝分裂,以维持精巢内精原细胞数量.
(2)在初级精母细胞分裂分化成精子的过程中,细胞体积逐渐变小,核染色质不断地浓缩,细胞形态也由不规则变成椭圆形或者圆形,核膜质膜由模糊逐渐变得清晰.
(3)在剑尾鱼精子的形成过程中,细胞核浓缩以后鞭毛才形成,这与长吻鮠的相似.
(4)在精子成熟的末期精子呈放射状整齐排列于精小囊内,头部指向囊壁,尾部指向精小囊中央腔.这种现象同样出现在隶属于鳉形目同为卵胎生的孔雀鱼[9]和食蚊鱼[11]精子发生过程中.此现象的生物学意义待进一步探讨.
(5)剑尾鱼精子头部细长,尾部中段有发达的线粒体鞘,表明精子细微结构是适应卵胎生发育演化的结果,精子头部没有项体,说明卵胎生鱼类的精子是介于卵生鱼类和胎生动物之间的中间类型[9].
3.4 体内受精方式
研究表明,剑尾鱼有交配器,生殖孔开在臀鳍基部的前面.
[1]温茹淑,方展强,陈庭伟.17β-雌二醇对雄性唐鱼卵黄蛋白原诱导及性腺发育的影响[J].动物学研究,2008,29(1):43-48.
[2]张耀光,罗泉笙,钟明超.长吻鮠精巢发育的分期及精子的发生和形成[J].动物学研究,1992:13(3):281-287.
[3]江寰新,尤永隆,林丹军,等.中华乌塘鳢鱼精巢的形态结构观察[J].福建农林大学学报,2004,33(1):89-93
[4]方展强,邱玫,王春凤.剑尾鱼鳃的光镜、扫描和透射电镜观察[J].电子显微学报,2004,23(5):553-559.
[5]方展强,林敏朝.剑尾鱼肝脏的显微和超微结构[J].电子显微学报,2006,24(3):265-270.
[6]林国辉,邱玫,方展强.剑尾鱼肾脏组织的超微结构观察[J].华南师范大学学报:自然科学版,2011(1):105-108.
[7]张小雪.黄鳝精巢显微和超微结构的研究[J].水利渔业,1994(4):9-14.
[8]冯俊荣.乌鳢精巢发育的研究[J].水利渔业,1995,(5):7-9.
[9]郭明申,刘龙,穆淑梅,等.孔雀鱼精子发生的显微与超微结构[J].河北大学学报,2006,26(6):653-658.
[10]刘文斌.黄颡鱼精巢发育和周年变化及精子的发生与形成[J].湖南师范大学自然科学学报,2004,27(1):66-70.
[11]刘灼见,高书堂,邓青.食蚊鱼的性腺发育及性周期研究[J].武汉大学学报,1996,42(4):487-490.
[12]DODD JM,HOARW S,RANDALLD J,etal.Reproduction in cartilaginous fishes(Chondrichthyes):Fish Physiology,Vol IXA:Reproduction[M].New York:Academic Press,1983:58-59.
[13]张旭晨,王所安.细鳞鱼精巢超微结构和精子发生[J].动物学报,1992,38(4):355-358.
