木聚糖酶生产工艺研究进展及性能优化
2011-03-09贺丹艳罗永发
贺丹艳 罗永发
1 前言
以植物性饲料为主的配合饲料,其原料中往往含有较多的非淀粉多糖(NSP),尤其是谷类作物及其副产品。而对于单胃动物而言,其消化道缺乏消化NSP的酶,因此,此类物质被认为是一种重要的抗营养因子。阿拉伯木聚糖是小麦类饲料中的主要NSP,其可以通过结合大量的水分(Geissmann等,1973)增加消化道食糜的粘性,降低食糜混合均匀性、饲料通过速率和肠道氧化作用,从而造成微生物大量繁殖并在小肠中发酵(Preston等,2001)。为了消除木聚糖的抗营养作用,木聚糖降解酶被广泛应用于生产,尤其是在国外,木聚糖酶的生产已比较成熟。文中在前人研究的基础上综述了木聚糖酶生产工艺的最新研究进展、产酶条件的优化及其分子改造手段,以期为木聚糖酶的生产、性能优化提供一些参考。
2 木聚糖酶的生产工艺
2.1 木聚糖酶
木聚糖酶是可将木聚糖降解成低聚木糖和木糖的一类酶的总称。糖苷键的彻底水解需要一系列主链和支链酶的共同作用,内切-1,4-β-木聚糖酶(EC 3.2.1.8),β-木糖苷酶(EC 3.2.1.37)和外切-1,4-β-木聚糖酶是降解主链最主要的酶(Paula Sá-Pereira等,2003)。内切-1,4-β-木聚糖酶以内切方式作用于木聚糖主链内部的β-1,4-木糖苷键,使木聚糖降解为短链的低聚糖;β-木糖苷酶是外切糖苷酶,通过催化低聚木糖的还原末端来释放木糖残基(Motoki Wakiyama等,2010)。外切-1,4-β-木聚糖酶(EC3.2.1.92),主要作用于木聚糖和木寡糖的非还原端,产物为木糖。木聚糖的结构及酶作用位点如图1所示(Joselena,1992)。

图1 木聚糖的结构及酶作用位点
另外,根据氨基酸序列相似度,可将木聚糖酶分为F/10家族和G/11家族。F/10家族分子量高(>30 kDa),等电点低,G/11家族分子量低(19~25 kDa),等电点高。与G/11家族相比,F/10家族有更好的催化功能和较低的底物特异性(P.Biely等,1997)。
2.2 产酶菌种
近年来,多种木聚糖酶陆续从微生物中被分离出来,其中包括细菌、放线菌、真菌以及某些酵母(Sunna等,1997)。多数细菌和真菌分泌胞外木聚糖酶,将木聚糖水解为单糖,单糖再进入微生物细胞供代谢用。也有一些微生物如瘤胃细菌、嗜饱粘液球菌及黑曲霉中的一些种类,也产生胞内酶(周秀梅,2008)。真菌木聚糖酶的活性通常要高于细菌木聚糖酶(陆健等,2001),细菌产生的木聚糖酶的pH值稳定性和热稳定性比真菌高,最适作用温度通常与产酶微生物的最适生长温度成正向相关。丝状真菌如曲霉属(Aspergillus sp.)和木霉属(Trichodermasp.),由于能分泌胞外木聚糖酶,且产酶水平高于酵母菌和细菌而格外引起研究人员的关注(Fang等,2007),但是丝状真菌产生木聚糖酶时往往伴随着纤维素酶的产生。一些杆菌和真菌微生物可以产生无纤维素酶活性的木聚糖酶(陆健等,2001)。
2.3 微生物发酵生产
目前木聚糖酶的生产主要依靠真菌、细菌等微生物发酵生产。按照发酵工艺的不同可分为固态发酵和液体发酵两种。固态发酵(SSF)是指微生物在无流动水的潮湿基质上生长,SSF的优点是产物回收的液体需求量少,底物便宜,培养成本低等(Q.K.Beg等,2001),但不易控制,酶系复杂,纤维素等酶含量高,难以精酶化。目前国内普遍采用固态发酵。液体发酵具有如下特点:①在液态环境中,菌体、底物、产物(包括热量)易于扩散,使发酵在均质或拟均质条件下进行,便于检测、控制,容易扩大生产规模;②液体输送方便,易于机械化操作;③产品易于提取精制。但缺点是对设备的要求较高,成本高,技术要求也较严格(Garjek W,1987;Roche N 等,1994)。
2.4 木聚糖酶的分离、纯化
木聚糖酶分离纯化的一般步骤是粗酶预处理、粗酶沉淀、色谱分离。纯化的早期步骤通常使用低分辨率、非专一性的粗分离纯化手段,如硫酸铵盐析、超滤、透析等以达到浓缩、去除大部分杂质和色素的目的;随后常用的色谱有离子交换、凝胶过滤、疏水色谱、亲和色谱和色谱聚焦(周玉恒等,2005)。
2.4.1 传统的非特异性分离
传统的非特异性分离方法一般包括硫酸铵沉淀、超滤浓缩、离子交换层析和凝胶过滤层析。
硫酸铵沉淀是酶分离纯化中常用的方法,可以同时达到浓缩与粗分离的目的。Gashaw等(2006)将从Bacillus halodurans S7获得的内切-1,4-β-木聚糖酶(分子量为43 kDa)粗酶液进行硫酸铵盐析,回收率为66.5%,并将比活从8.9 U/mg提高到了13.2 U/mg。但对于低分子量木聚糖酶的纯化来说,可能是限制其回收率的主要因素。Paula S.P.等(2002)在纯化Bacillus subtilis的XylⅡ时,发现硫酸铵沉淀这一步使酶的回收率降低了62%,并且其它的浓缩方法回收率也不高。
在一定的条件下,采用不同切割分子量的超滤柱对木聚糖酶进行分级分离,可对木聚糖酶起到一定的分离精制作用(刘蕾等,2010)。Puji Rahayu等(2010)从Bacillus sp.AQ-1中分离木聚糖酶,采用切割分子量为30 kDa的聚醚砜膜进行超滤,分离出了大小分别为15.7 kDa和57.7 kDa的木聚糖酶A和木聚糖酶B。采用超滤方法有时会造成目的酶的大量损失,可能与酶的结构或者膜孔的分布不均有关。同时,目的木聚糖酶结构中如果含有纤维素结合区时,膜材料最好不要选择纤维素膜,因为由于木聚糖酶与膜的吸附作用可能造成大量损失(孙雷等,2005)。
离子交换层析技术是木聚糖酶分离纯化中常采用的方法。根据目的酶的等电点选择合适的分离树脂,对于弱酸性及中性的木聚糖酶多采用阴离子交换树脂,如 DEAE-Toyopearl 650S(Motoki Wakiyama 等,2010);对于弱碱性的木聚糖酶多采用阳离子交换树脂,如 DEAE-Sepharose CL-6B(Fang 等,2008)。Ashwani等(2010)在分离Bacillus subtilis ash的木聚糖酶时,采用CM-Sephadex C-50离子交换一步法,回收率为 43%,纯化倍数为 10.5。Suchita Ninawe等(2008)将Streptomyces cyaneus SN32产生的木聚糖酶粗酶液经过硫酸铵沉淀后,再用DEAE Sepharose进行离子交换,得到的产物回收率为43.62%,纯化倍数为2.254。
凝胶过滤是一种根据分子大小不同分离纯化木聚糖酶的常用分离方法之一。经过凝胶过滤层析后木聚糖酶的回收率一般都较低,这可能由于木聚糖酶结构中含有纤维素结合区 (Cellulose-Binding Domain,CBD),而葡聚糖是由α-葡萄糖残基构成的,两者可以相互结合(孙雷等,2005)。Wang等(2010)将 Bacillus sp.NTU-06产生的木聚糖酶经过离子交换,然后将活性部分注入Sephadex G-200凝胶,结果回收率为16.1%,纯化倍数为36.7,并且凝胶SDS-PAGE电泳后,得到的条带颜色变浅,这是由于活性木聚糖酶使凝胶中的木聚糖基质发生水解。
2.4.2 特异性分离纯化
应用特异性步骤分离纯化木聚糖酶,一是利用木聚糖酶自身结构上可以与多糖结合的能力。另外,是利用基因技术使木聚糖酶结构中带有亲和标记(孙雷等,2005)。这些特异性分离纯化方法主要有与多糖特异性结合亲和层析、双水相萃取、三相分离等。
除了催化区,一些木聚糖酶还含有非催化区即糖类结合域(carbohydrate-binding module,CBM),CBM可以提高木聚糖酶与不溶性基质结合的能力(Liu等,2010)。CBM主要包括纤维素结合域CBD(cellulosebinding domain)和木聚糖结合域XBD(Xylan-Binding Domain)。CBD可专一地吸附于可溶或不可溶的纤维素上。一些CBD不可逆地结合在纤维素上,此类CBD可用于固定化。具有此CBD的木聚糖酶或者含此CBD标签的酶可固定到纤维素上。另一些CBD可逆结合到纤维素上,可作为一种亲和标签,在分离和纯化上更为有用(周玉恒等,2005)。木聚糖酶中的XBD较CBD而言较少存在,其原因可能是木聚糖多聚物中带有许多类型的取代基,由于取代基的变化从而使能够结合所有木聚糖蛋白区域的进化成为不可能(BlackG.w 等,1995)。
双水相系统(Aqueous Two-Phase System,ATPS)主要是通过混合两种低浓度的不相溶高聚物溶液或者一种高聚物溶液和一种盐溶液形成。ATPS系统分离的基础是两相之间物质的选择性分配,由与相位系统特征相关的一系列参数和物质以及他们的相互作用控制(Albertsson,1986)。与其他分离纯化相比,由于ATPS系统的含水量高,处理时间短,能量消耗低,所以其具有更好的生物兼容性(Andersson等,1990)。Yang等(2008)利用聚乙二醇(PEG)/硫酸铵系统纯化来自嗜热拟青霉(Paecilomyces thermophila)通过固态发酵产生的一耐热木聚糖酶,研究了6个包含不同的PEG分子量、硫酸铵浓度的ATP系统,结果在pH值7.2、12.5%PEG-4000、25%硫酸铵和50%酶溶液时最优,回收率为98.7%,纯化倍数为5.54。
三相系统与ATPS不同,即在上下相之间有一界面相,一般由蛋白质水溶液中加入一种盐(一般是硫酸铵)和一种有机溶剂(一般是t-丁醇)形成(Dennison等,1997)。Ipsita等(2004)先将 Aspergillus niger木聚糖酶用8 mol/l尿素和100 mmol/l二硫苏糖醇变性,再进行TPP纯化,木聚糖酶达到了93%的回收率和21倍纯化,且实现了该酶变性后的复性。
除了上面的技术外,Everaldo Silvino Dos Santos等(2002)曾尝试用扩张床吸附技术(Expanded bed adsorption,EBD)纯化木聚糖酶,虽然最大只有30倍的纯化和21.8%的回收,但此法具有既经济又省时,木聚糖酶发酵液不需沉淀澄清的优点。
3 木聚糖酶的优化
3.1 产酶条件的优化
3.1.1 筛选菌株
要提高木聚糖酶的活力及性能,第一步就是要筛选出高产菌株和特异性菌株(如耐热性能好、pH值适应范围广等)。
3.1.1.1 自然选育
自然选育是指对微生物细胞群体不经过人工处理而直接进行筛选的育种方法。虢国成(2008)利用透明圈比较法和DNS法相结合,筛选出26株产木聚糖酶菌,并从中分离、纯化出一株高产木聚糖酶菌-S6。代义等(2008)以半纤维素、桦木木聚糖为唯一碳源,两步透明圈法从10种混合土壤样品中,分离到一株木聚糖酶高产菌株HJ-04,通过形态学观察、生理生化特征及16SrDNA序列分析,鉴定为短小芽孢杆菌。赵峰等(2008)采用木聚糖为唯一碳源的平板初筛培养基以及摇瓶复筛液体培养基,从保藏的菌株中筛选到产木聚糖酶活力较高、菌丝生长快的粗糙脉孢菌(Neurospora crassa)菌株。另外,已报道的耐热真菌有嗜热毛壳菌(Humicola insolens)、特异腐质霉(Humicola insolens)、嗜热羊毛状腐质霉(Humicola lanuginosa)、灰色腐质霉(Humicola grisea)、嗜热拟青霉(Melanocarpus albomyces)、埃默森篮状菌(Talaromyces emersonii)、丝衣霉状篮状菌(Talaromyces byssochlamydoides)、疏绵状嗜热丝胞菌 (Thermomyces lanuginosus)、耐热子囊菌(Thermoascusaurantiacus)等。
3.1.1.2 人工诱变
关于高产菌株的筛选目前已有很多报道,人工诱发基因突变是主要改良途径之一。徐同宝等(2009)以黑曲霉 (Aspergillus niger)XE6为出发菌株,经微波(MW)和硫酸二乙酯(DES)诱变处理,选育出一株遗传性状稳定的高产木聚糖酶菌株mAn1。周欣等(2009)以短小芽孢杆菌 (Bacillus pumilus)HJ-04为出发菌株,通过紫外诱变的方法筛选得到一株木聚糖酶高产菌株B-6,其酶活提高约30%。刘唤明等(2008)以木聚糖酶酶活为20000 U/g的黑曲霉P14为出发菌株,通过离子注入诱变,得到产木聚糖酶酶活32000 U/g的菌株L15。
3.1.1.3 构建基因工程菌
微生物产生木聚糖降解酶系比较复杂,大多还同时产生纤维素酶,这些酶性相近,使分离纯化木聚糖酶比较困难,利用基因重组技术,将产生木聚糖酶的基因在合适的宿主菌中表达,可比较容易地得到纯化。Panbangred和Fukusaki(1985)首次利用大肠杆菌表达矮小芽孢杆菌(Bacillus.pumilus IOP)β-木聚糖酶。随后已有数十种不同来源的木聚糖酶基因在原核生物表达系统中实现了表达。Wang等(2010)将从Anoxybacillus sp.E2中克隆的木聚糖酶基因xynE2导入大肠杆菌BL21(DE3)中表达,纯化后得到的重组XynE2在pH值6.6~8.6范围内能保持最大活力的90%,在pH值4.6~12.0的范围内仍能保持80%的活力1 h。Qiu等(2010)从Streptomyces megasporus DSM 41476基因文库中筛选出xynAM6基因,并在毕赤酵母GS115中成功表达其成熟肽,结果表明,该酶温度适应范围广(50~80℃范围内能保持最大活力的60%),并在60℃和70℃时有良好的热稳定性,在pH值4.0~11.0范围内保持稳定。
3.1.2 发酵条件的优化
3.1.2.1 选择合适的诱导物
由细菌和真菌产生的木聚糖酶通常是诱导型的,但也有很少一部分木聚糖酶是组成型的。木聚糖作为高分子的多聚物不能够透过细胞壁,在少量组成型酶的作用下释放出低聚糖碎片,这些碎片包括木糖、木二糖、低聚木糖等(Bastawde,1992)。其作用机制见图2(Polizeli等,2005)。
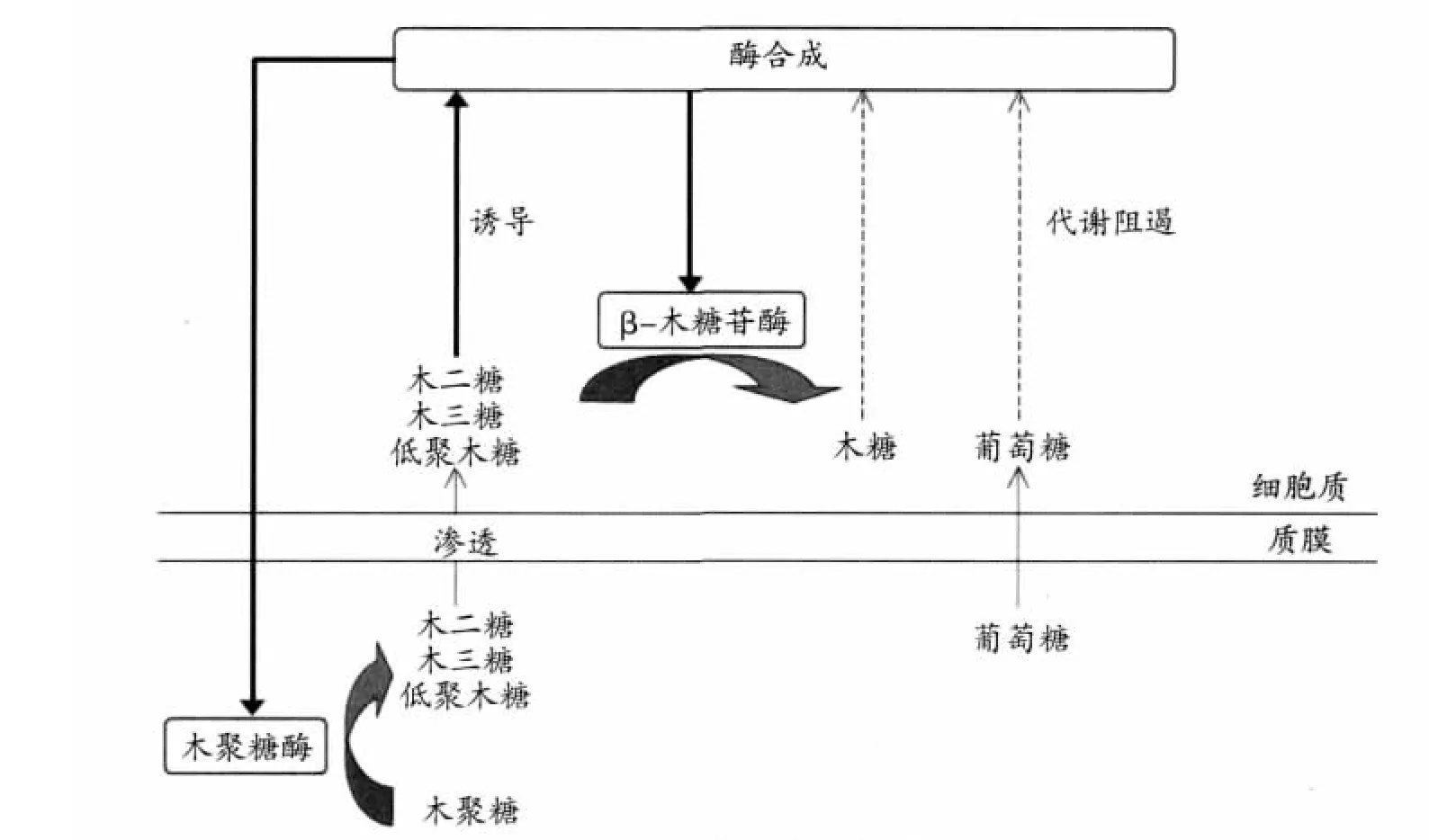
图2 木聚糖酶的作用机制
当单糖如木糖、半乳糖、果糖、葡萄糖等作为唯一碳源时,一般都不发挥诱导作用,而且由葡萄糖引起的代谢阻遏是木聚糖酶生物合成中的普遍现象。在海枣曲霉(Aspergillusphoenicis)中,由木聚糖、木糖和β-甲基木糖苷诱发的木聚糖酶活性主要是胞外的,向木聚糖或者木糖基质中加入1%的葡萄糖抑制木聚糖酶的表达,但这种抑制可以通过添加cAMP或者联丁酰cAMP 得到缓解 (Ana等,2008);Kohji Miyazaki等(2005)研究发现,对于布氏普雷沃氏菌B14(Prevotella bryantii B14),木糖、木二糖、木三糖、木四糖、木戊糖、树胶醛糖、葡萄糖醛酸皆不能诱导其产生木聚糖酶,并且当葡萄糖水平达到0.1%(w/v)时即会产生代谢阻遏或者诱导排斥,从而造成木聚糖酶表达的延迟。
一般来讲,木聚糖酶的诱导是复杂的,其诱导水平也因微生物的不同而相差较大,对于某一种微生物可以产生最大木聚糖酶活性的诱导物可能是另外一种微生物产木聚糖酶的抑制剂。在白腐真菌粗毛栓菌(Trametes trogii)(Levin 等,1998)、 泡盛曲霉(Aspergillus awamori)(Siedenberg 等,1998)、链霉菌sp.QG-11-3(Beg等,2000)中诱导物一般是木聚糖,而在黄纤维单胞菌(Cellulomonas flavigena)(Avalos等,1996)中木聚糖的诱导效果则不好。β-D吡喃木精基可以作为木聚糖酶组的诱导物 (Ghosh等,1994;Rizzatti等,2001),但在一些微生物中它将导致内切木聚糖酶的代谢阻遏(Flores等,1996;Mach 等,1996)。另外,木聚糖酶的其他一些诱导物如L-山梨糖、各种木寡糖、木质纤维素残渣等都有过报道。
3.1.2.2 选择合适的氮源
氮源是构成细胞原生质和酶蛋白的主要原料,因此对微生物的生长和酶的合成也会有所影响。据Haapala等(1996)研究表明,以固定化里氏木霉QM 6a制备木聚糖酶时,氮源对产酶的影响比pH值更为重要。一般来说,单一物质作氮源时有机氮源优于无机氮源。Sushil等(2010)以短小芽孢杆菌SV-85S(Bacillus pumilus SV-85S)为出发菌株,以1.0%的小麦麸为碳源,研究蛋白胨、酵母提取物、牛肉膏、以及硝酸钾、硫酸铵、硝酸钠对木聚糖酶活性的影响,结果表明,添加有机氮源时酶的滴定度更高,另外,不同氮源的组合效应表明,蛋白胨、酵母提取物和硝酸钾组合时酶的产量最高;对于环状芽孢杆菌AB16(B.circulans AB16)和喜热噬油芽孢杆菌 (Geobacillus thermoleovorans)而言,最好的氮源则是胰蛋白胨。尽管如此,但也有一些研究结果得出无机氮源和有机氮源的效果无显著差异,或者无机氮源优于有机氮源。在苜蓿根腐病菌F7(Fusarium solani F7)的培养基中分别以蛋白胨、尿素、酵母提取物、硝酸钠、硫酸铵、硝酸铵和牛肉膏为氮源,结果表明,蛋白胨和硝酸钠的效果最好,并且硝酸铵和尿素也能产生相当数量的木聚糖酶(Gupta等,2009);短小芽孢杆菌HJ-04(Bacillus pumilus HJ-04)NH4NO3最利于产酶,总体是无机氮源比有机氮源更利于HJ-04产酶(代义等,2008)。黑曲霉 (Aspergillus niger)XE6以NH4NO3为氮源时菌株的产酶活性显著高于其它氮源(徐同宝等,2009)。
3.1.2.3 固定化
在实际应用中,将微生物或者酶固定在固态材料上具有许多优点,包括酶的重复利用、易于产品分离、提高酶的稳定性等。在固定化研究中,可把微生物的整个细胞固定在支持物上,酶本身也可固定在某些可溶或不溶的聚合物上(Beg等,2001)。为此,选择合适的技术和支持物是非常必要的。在大多数已知的载体中,明胶和壳聚糖通常比较理想,因为它们生物兼容性好并且无毒(Achouri等,2009)。在 Manrich等(2010)的研究中,将木聚糖酶固定在环氧壳聚糖上,固定率达到100%,并且能保留64%的酶活,而将其固定在乙醛琼脂糖上时,固定率和稳定性都较差。Mohamed Guerfali等(2009)发现,与其他支持物相比,将嗜热踝节菌(Talaromyces thermophilus)的 β-木糖苷酶固定在用戊二醛处理的壳聚糖上时,固定率和酶活均最高,分别达到94%和87%,并且在循环25次后仍能保留最初酶活的94%。另外,与游离酶相比,固定酶的热稳定性提高了,固定酶在pH值8.0和53℃时达到最大活力,而游离酶达到最大活力的pH值和温度分别为7.0和50℃。已知报道过的固定化材料还有海藻酸钠(Sanjay Kar等,2008)、聚氨基甲酸乙酯泡膜(PUF)(Uma Gupta等,2008)、米碳管(Shweta Shah等,2008)、Eudragit L-100(周玉恒等,2007)、尼龙布(Haapala,1996)等。
除了固定化材料的固有性质外,对其进行适当修饰也可改善其性能。支持物与酶之间的共价键数量由支持物的活化度(支持物表面的醛基数目)和酶分子的氨基数目决定(J.M.Guisán,1988),可以通过增加酶分子表面的氨基数目来提高固定化酶的性能。Manrich等(2010)将木聚糖酶固定在乙醛琼脂糖上时,固定率和稳定性都较差,这主要是因为酶分子的赖氨酸基团的数目较低,最后经过乙二胺修饰,酶的稳定性提高了40倍,并且固定率达到了100%。
尽管固体支持物有很多优点,但一些缺点还是不可避免的,比如木聚糖基质的固态性能会限制酶的通过,并且由于结合酶不易与大体积的不溶性基质接触,使木聚糖酶的效率有可能降低。为了克服这点,有些学者提出了一种新的酶固定化技术,即基于人工油体(AOBs)的表达/纯化系统。在这个系统中,种子油体的一种独特的结构蛋白油脂蛋白被作为载体,油脂蛋白的亲脂部分嵌入三酰甘油中,两个两性手臂突出于油体表面,从而,目标蛋白可以融合到油脂蛋白的N或者C末端形成不溶性重组蛋白,而后在大肠杆菌中表达(Hung等,2008)。
3.2 木聚糖酶的分子改造
大多数天然酶的活力较低,而且催化特性也往往不能满足人们的需求,因此对酶分子进行改造具有十分重要的意义。几十年来对酶分子的改造工作可以归纳为以下几个方面:酶的化学修饰、定点突变、杂合进化和定向进化等。
3.2.1 定点突变
酶本身的一级结构对其耐热性具有重要作用。酶的一级结构中某些关键区域的个别氨基酸改变,就会引起高级结构的变化,使酶蛋白结构中的氢键、离子键或疏水键稍有增加,从而提高整个分子的热稳定性(连惠芗,2009)。Zhang等(2010)将来自嗜温菌橄榄灰绿链霉菌 (Streptomyces olivaceovirdis)的木聚糖酶SoxB的N末端第33位氨基酸残基用来自嗜热菌以褐色高温单胞菌(Thermomonospora fusca)的木聚糖酶TfxA相应残基取代,发现其热稳定性大大提高。Kim等(2010)研究表明,Cohnella laeviribosi HY-21产生的木聚糖酶iXylc的色氨酸W89A突变使其对PNP-吡喃木糖基的催化活性提高了3.4倍,而W217A和W315A突变则使其催化活性提高了大约8.3倍。
3.2.2 定向进化
酶的体外定向进化,又称分子进化,就是人为地创造特殊的进化条件,模拟自然进化机制(随机突变、重组和自然选择),在体外对酶基因进行随机诱变,用一定的筛选或选择方法定向选出所需性质的突变酶。目前已发展出多种定向进化的策略,如易错PCR(erro-prone PCR)、DNA 重排(DNA shuffling)、体外随机引发重组(random-priming in vitro recombination,PRP)、交错延伸(stagger extension process,StEP)等(连惠芗,2009)。表1列出了一些木聚糖酶体外定向进化的成功实例。

表1 木聚糖酶体外定向进化的成功实例
4 结语
国外对木聚糖酶的研究较早,在1992年就已经实现了酶制剂的工业化生产(Senior等,1992)。随着分子生物学的发展和众多新的试验方法的引入,对木聚糖酶的研究已经达到了分子生物学基因工程的水平。国内对木聚糖酶的研究起步较晚,作为一种工业酶制剂在我国尚属空白。虽然木聚糖酶在众多的领域都有极大的应用价值,其研究也取得了一定的进展,但目前仍然存在着一些缺点,如稳定性差、使用效率低、半衰期短等,使酶的生产和使用受到极大限制。今后需从以下几个方面来改造木聚糖酶,以拓宽木聚糖酶的应用范围:①木聚糖酶的耐热性一般不好,在生产和处理过程中稳定化技术研究是今后研究方向之一,可以考虑通过包被等方法加以解决;②运用基因工程手段从根本上改善木聚糖酶的性质,使其具有更高的热稳定性,并拓宽其pH值适应范围等;③寻找更多的产酶优化手段,来高效地生产高活力的木聚糖酶。
