蚯蚓原位堆肥与不同比例生物炭对基质理化性质及番茄品质的作用研究
2024-04-17李文慧王继涛安明远徐广亚尹翠游宏建曹云娥
李文慧 王继涛 安明远 徐广亚 尹翠 游宏建 曹云娥,*
(1宁夏大学葡萄酒与园艺学院,宁夏 银川 750021;2宁夏回族自治区园艺技术推广站,宁夏 银川 750001;3宁夏共享人力集团有限公司,宁夏 银川 750027)
番茄(SolanumlycopersicumL.)在我国种植广泛且经济利益较高,近年来,其生产方式基本由传统的露地栽培转变为设施栽培[1]。然而,设施农业高集约化和高复种指数导致土壤微生物群落失衡,土传病害加重及土壤性质破坏等一系列土壤退化问题[2-3]。采用基质栽培是缓解此类问题的有效措施之一[4]。前人研究以菇渣、牛粪、蛭石为原料,进行了番茄基质配方的筛选,发现当牛粪∶菇渣∶蛭石体积比为6∶3∶1时,番茄的产量和品质显著提高[5]。
然而,目前无土栽培相关的国标、行标等都较少,市面销售的栽培基质物料配比、养分构成也不尽相同,难以开展标准化无土栽培体系研究[6]。蚯蚓(Pheretima)作为生态系统中常见的分解者,可以将有机固体废弃物分解转化为腐殖质,故蚯蚓堆肥作为一种富含有机质、养分和腐殖质的新型基质而备受关注[7-8]。相关研究表明,施用蚯蚓堆肥可使黑籽(NigellasativaL.)籽粒氮、磷、钾含量提高13.50%、10.80%和14.10%,黑籽油中的亚油酸、油酸和亚麻酸含量提高53.53%、25.36%和0.60%[9]。为使蚯蚓堆肥长期稳定地向栽培体系输送养分,保证番茄的稳产高质,本研究特引入“蚯蚓原位堆肥”(in-situ vermicomposting)——一种改良的蚯蚓堆肥技术。该技术将蚯蚓及共生菌群引入有机废弃物(畜禽粪便、餐厨垃圾、农田果园残留物等)中,其分解、转化有机废弃物的全过程不仅直接在待处理地块自发完成,亦结合当地耕作制度实时补充有机废弃物物料,以保证养分长期供应充足。此外,生物炭作为一种物理结构稳定、化学性质呈现惰性的富碳材料,被广泛应用于农业研究[10]。生物炭可影响土壤微生物群落结构及多样性,调控环境因子和微生物之间的互作关系,促进并改良微生物生态系统,从而对受微生物驱动的氮、磷、钾营养元素循环转化产生影响,最终改变土壤质量和功能[11]。Huang等[12]发现玉米芯生物炭既促进了细菌的数量和多样性,又能进一步加速脱水污泥蚯蚓堆肥过程中有机物的矿化和腐殖化。同时,生物炭可有效减少NH3的挥发,进而将NH4+转化为NO3-,达到氮肥的缓释效果,为作物和根系生长提供充足的养分,提高氮素利用效率[13-14]。
目前针对蚯蚓(原位)堆肥和生物炭作为栽培基质的应用均较成熟,但将蚯蚓原位堆肥与生物炭配合施用,探究其对养分、菌群及农产品产量与品质构成的影响仍鲜有报道。因此,本研究通过向蚯蚓原位堆肥中添加不同比例的生物炭,探究比较不同处理的理化性质、菌群组成及番茄代谢发育特征,旨在为此后相关研究提供基础数据。
1 材料与方法
1.1 试验地概况
试验于2021年2月24日—7月2日在宁夏贺兰县园艺产业园区的日光温室(106.33°E,38.58°N)进行。该基地属于国家级农业示范园区,年降雨量180~200 mm,80%的降雨量集中在6—8月,年平均气温8.8 ℃,无霜期160~170 d,属典型的中温带干旱气候区,具有大陆性气候特征。
1.2 试验材料
试验番茄品种为粉得力(武汉楚为生物科技股份有限公司);试验所需蚯蚓和牛粪购自宁夏万辉生物环保科技有限公司;生物炭购自上海海诺碳工业有限公司。生物炭和蚯蚓原位堆肥基本特性见表1。

表1 基础材料理化性质Table 1 Physicochemical properties of basic materials
1.3 试验方法
采用单因素随机区组设计。共设6个处理,分别为蚯蚓原位堆肥(CK)、蚯蚓原位堆肥+1%生物炭(C1)、蚯蚓原位堆肥+2%生物炭(C2)、蚯蚓原位堆肥+3%生物炭(C3)、蚯蚓原位堆肥+5%生物炭(C4)、蚯蚓原位堆肥+10%生物炭(C5),每个处理重复5次,随机区组排列。
试验将基质槽一分为二,种植垄和养殖垄尺寸均为4.1 m(长)×0.35 m(宽)×0.2 m(深),对照垄每垄铺设蚯蚓原位堆肥226.73 kg,铺设牛粪0.38 m3。根据等碳原则,种植垄添加生物炭替代蚯蚓原位堆肥中部分总碳,养殖垄投放一定密度蚯蚓处理牛粪产生蚯蚓堆肥。待种植垄一茬作物收获,养殖垄中牛粪全部处理为蚯蚓堆肥后,养殖垄与种植垄模式调换,原养殖垄原位处理生成的蚯蚓堆肥中定植作物,原种植垄中重新铺设牛粪进行蚯蚓原位处理,为下一茬作物定植做准备,最终实现作物原位轮作。番茄单行栽植于种植垄,株距15 cm,行距140 cm,所有处理采用水肥一体化滴管装置进行水肥管理,番茄整个生育期灌水量约1 400 t·hm-2,肥料选用宁夏万辉生物环保科技有限公司生产的茄果类专用滴灌配方肥(24.0% N+3.0% P2O5+21.0% K2O),全生育期共投入氮肥378 kg·hm-2、磷肥47.25 kg·hm-2、钾肥330.75 kg·hm-2。
1.4 测定项目与方法
1.4.1 基质理化性质测定 于番茄盛果期(2021年5月18日),采用5点取样法从栽培基质中部取基质500 g,每个处理5次重复,风干后过1 mm筛。根据马全会等[15]的方法测定基质容重、孔隙度、含水量。使用pH仪和DDS-307A EC仪(上海精密科学仪器有限公司)测定基质pH值和电导率(electric conductivity,EC)(土∶水=1∶10)。有机质含量测定采用重铬酸钾氧化法;全氮(total N,TN)含量测定采用硫酸消煮-凯氏定氮-硼酸指示剂-0.01 mol·L-1硫酸滴定法;全磷(total P,TP)含量测定采用HCIO4-H2SO4消化-钼锑抗比色法[16]。分别采用0.5 mol·L-1K2SO4、0.5 mol·L-1NaHCO3萃取-钼锑法和NH4OAc萃取-火焰光度法测定速效氮(available N,AN)、速效磷(available P,AP)和速效钾(available K,AK)含量[16]。
土壤脲酶活性采用苯酚钠法测定;蔗糖酶活性采用3,5-二硝基水杨酸法测定;过氧化氢酶活性采用高锰酸钾法测定;碱性磷酸酶活性采用磷酸苯二钠法测定[16]。
1.4.2 微生物丰度指标测定 于番茄盛果期(2021年5月18日)采集新鲜基质样品进行微生物群落多样性分析。根据QIAamp DNA MiniKit试剂盒(Qiagenn,德国)说明书进行微生物群落总DNA抽提,使用1%的琼脂糖凝胶电泳检测DNA的提取质量,使用NanoDrop 2000(美国Thermo Fisher Scientific公司)测定其DNA纯度和浓度。采用引物338F(5'-ACTCCTACGGGAG GCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCT AAT-3')对细菌16S rRNA基因V3-V4的可变区进行PCR扩增及高通量测序;引物ITS2R(5'-GCTGCGTTC TTCATCGATGC-3')和ITS1F(5'-CTTGGTCATTTAGA GGAAGTAA-3')对真菌进行PCR扩增及高通量测序。细菌PCR反应体系:5×FastPfu Buffer缓冲液4 μL,2.5 mmol·L-1dNTPs 2 μL,5 μmol·L-1上、下游引物各0.8 μL,FastPfu Polymerase DNA聚合酶0.4 μL,模板DNA 10 ng,ddH2O补足至20 μL。真菌PCR反应体系:10×Buffer缓冲液2 μL,2.5 mmol·L-1dNTPs 2 μL,5 μmol·L-1上、下游引物各0.8 μL,rTAP Polymerase DNA聚合酶0.2 μL,模板DNA 10 ng,ddH2O补足至20 μL。程序为:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,细菌27个循环,真菌35个循环;72 ℃终延伸10 min。测序数据优化处理、操作分类单元(operational taxonomic unit,OTU)聚类分析及注释、多样性及群落结构分析等均在上海美吉生物科技有限公司的I-Sanger平台进行,用QIIME 1.9.1软件进行稀疏分析,以区分OTU丰富度及α多样性指数[覆盖度(Coverage)、Shannon、Simpson、Chao1、Ace指数][17];使用CANOCO 5.0软件对基质微生物门、属水平群落结构、相对丰度等与基质理化性质进行冗余分析(redundancy analysis,RDA)。
1.4.3 番茄生长量、品质和产量测定 于盛果期,每个处理随机选取植株6株,取下大小均匀果实10个,测定果实质量。可溶性固形物含量采用TD-45数字折光仪(中国金科利达公司)进行测定。有机酸和维生素C(vitamin C,Vc)含量分别通过NaOH滴定法和2,6 -二氯苯酚直接滴定法进行测定。此外,分别通过二硝基水杨酸和蒽酮比色法测定硝酸盐和可溶性糖含量[18]。全生育期记录每个处理产量,按照小区面积产量折合成番茄产量。
测定果实品质的同时,在每个处理中随机选取植株6株。用天平测定地上部与地下部鲜重后,每个植株装进信封并标记,将样品置于烘箱105 ℃下杀青30 min,80 ℃烘干至恒重。用电子天平称量地上部与地下部干重。
1.4.4 高效液相色谱-质谱联用(chighperfomanceliquid chromatography-tandem mass spectrometry, LC-MS)检测 在1 000 mg番茄鲜样中加入1.3 mL提取液(甲醇∶水=4∶1,V∶V),利用Wonbio-96c型冷冻组织研磨仪(上海万柏生物科技有限公司)破碎,低温涡旋混匀处理30 min。4 ℃、13 000 r·min-1离心15 min,取上清后重复上述萃取操作3次,将上清液在氮气下混合浓缩。最终,样品用乙腈∶水=1∶100 μL(V∶V)复溶后用于液相色谱-质谱分析。上机完成之后,在美吉生物云平台(https://cloud.majorbio.com)进行差异代谢物分析,筛选出差异代谢物,并通过KEGG(kyoto encyclopedia of genes and genomes)数据库进行代谢通路注释,获得差异代谢物参与的通路[19]。
1.5 数据统计与分析
采用Excel 2021对数据进行整理与统计,采用SPSS 26.0软件对数据进行单因素方差分析(ANOVA)及相关性分析,采用Origin 2021软件作图。
2 结果与分析
2.1 不同比例生物炭对蚯蚓原位堆肥理化性质的影响
由表2可知,添加生物炭增大了蚯蚓原位堆肥的总孔隙度,减小了容重,但各处理间无显著差异(P>0.05)。
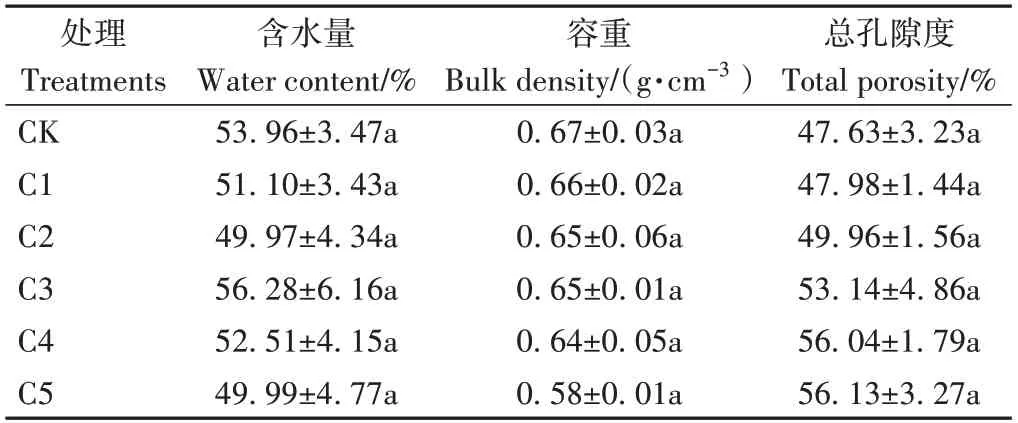
表2 生物炭对蚯蚓原位堆肥物理性质的影响Table 2 Effect of biochar on the physical properties of in-situ vermicomposting
由表3可知,与CK相比,添加生物炭能显著提高蚯蚓原位堆肥的pH和EC值(P<0.05),并且随着生物炭添加比例的增大,pH和EC值呈持续上升趋势。有机质含量也表现出相同趋势,从121.50 g·kg-1增加到310.50 g·kg-1。添加生物炭提高了蚯蚓原位堆肥的养分含量,并且随着生物炭添加比例的增大,蚯蚓原位堆肥中全氮、速效氮、速效磷、全磷和速效钾含量均呈现先增加后降低趋势。其中,全磷、速效磷和速效钾含量以C3处理最高,分别较CK显著提高了46.27%、10.10%和68.43%;全氮和速效氮含量以C4处理最高,分别较CK显著提高了15.96%和92.20%。
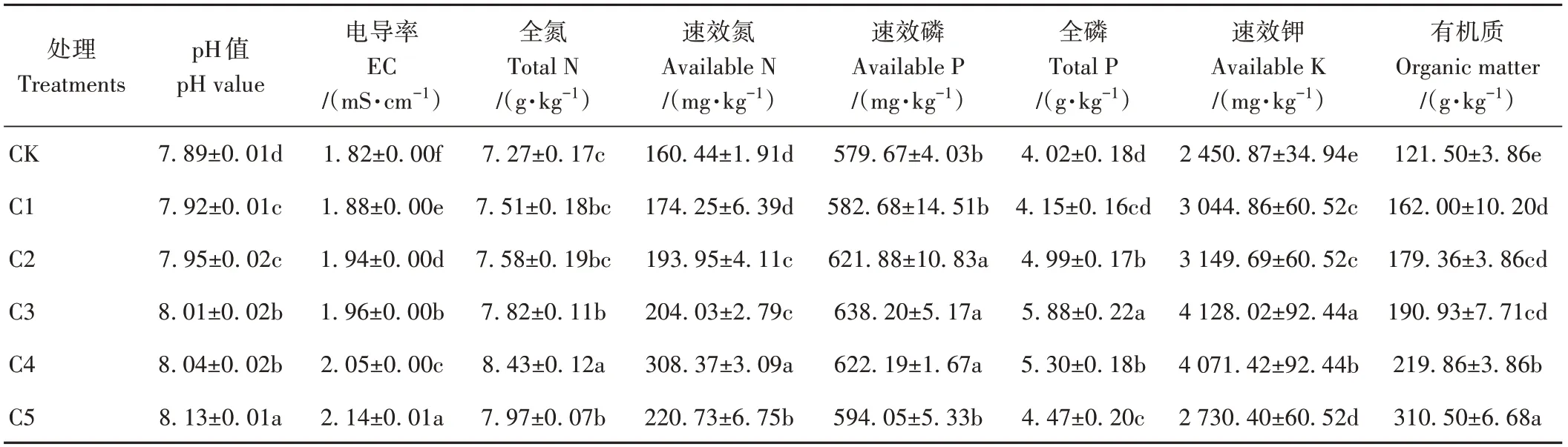
表3 生物炭对蚯蚓原位堆肥化学性质的影响Table 3 Effect of biochar on the chemical properties of in-situ vermicomposting
2.2 不同比例生物炭对蚯蚓原位堆肥酶活性的影响
由图1可知,添加生物炭能提高蚯蚓原位堆肥中的酶活性,并且随着生物炭添加比例的增大,过氧化氢酶活性呈持续增大趋势;蔗糖酶、脲酶、碱性磷酸酶活性均呈先增大后减小趋势。其中,蔗糖酶和碱性磷酸酶活性以C3处理最高,分别较CK显著提高了52.09%和18.66%;脲酶活性以C4处理最高,较CK显著提高了86.49%。
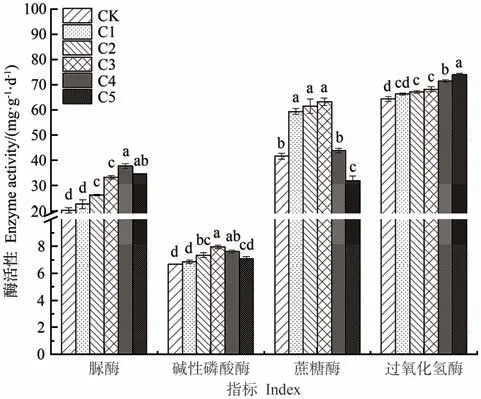
图1 生物炭对蚯蚓原位堆肥酶活性的影响Fig.1 Effect of biochar on enzyme activity of in-situ vermicomposting
2.3 不同比例生物炭对蚯蚓原位堆肥微生物多样性的影响
由图2可知,细菌群落中各处理共产生5 567个OTU,其中共有OTU数量为2 422,占总数的43.51%。CK~C5处理特有OTU数分别为41、46、44、65、51、68。相较CK,C1~C5处理特有OTU数量均有不同程度增加,以C5处理特有OTU数量最多。这可能是因为生物炭多孔的结构有利于微生物的生长发育。由此表明,生物炭能增加蚯蚓原位堆肥中的特有细菌数量,对细菌的产生可能具有一定的促进作用。
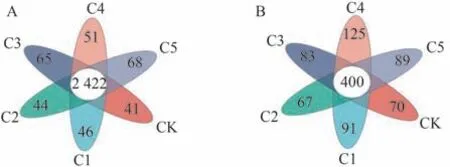
图2 生物炭对蚯蚓原位堆肥细菌(A)和真菌(B)OTU水平的影响Fig.2 Effect of biochar on the OTU levels bacteria (A) and fungi (B) of in-situ vermicomposting
真菌群落中各处理共产生1 671个OTU,其中共有OTU数量为400,占总数的23.94%。CK~C5处理特有OTU数分别为70、91、67、83、125、89,除C2处理外,其他处理的特有OTU数量均高于CK,变化趋势与细菌基本一致,以C4处理特有OTU最多。C5处理特有OTU小于C4处理的原因可能是过量生物炭对真菌的产生具有抑制作用。
为了进一步分析生物炭对微生物群落多样性的影响,采用五种指标(覆盖度、Ace指数、Chao1指数、Simpson指数以及Shannon指数)计算蚯蚓原位堆肥中的细菌和真菌丰度。由表4可知,细菌和真菌覆盖度(Coverage)分别达96%和99%以上,说明蚯蚓原位堆肥中所测出的细菌、真菌序列数足以反映其真实情况。不同处理间细菌Chao1指数和Ace指数无显著差异;随着生物炭添加比例的增大,各处理细菌Shannon指数呈先降低后升高趋势,Simpson指数呈先升高后降低趋势,其中,C3处理的Shannon指数和Simpson指数相较CK差异显著,细菌多样性最低。真菌Chao1指数、Ace指数、Shannon指数和Simpson指数在不同处理间均无显著差异。
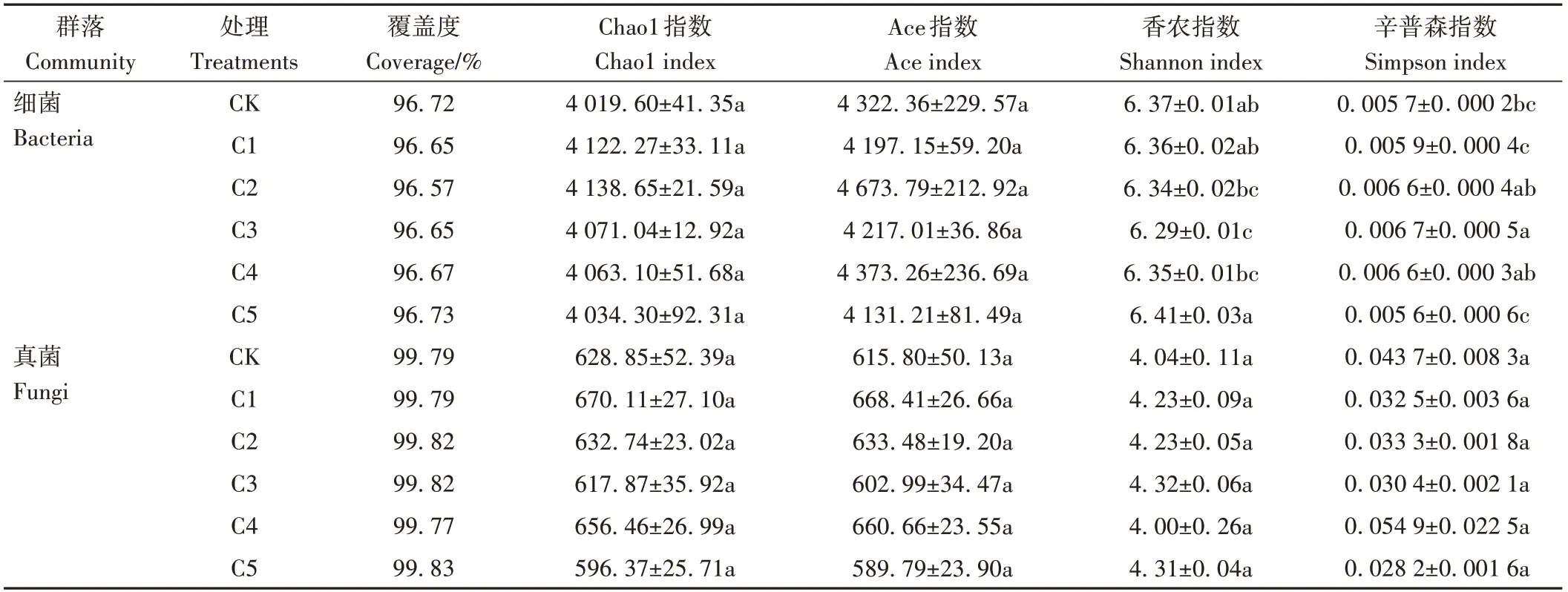
表4 生物炭对蚯蚓原位堆肥细菌和真菌群落Alpha多样性指数的影响Table 4 Effect of biochar on Alpha diversity index of biochar on Alpha diversity index of in-situ vermicomposting with bacterial and fungal communities
2.4 不同比例生物炭对蚯蚓原位堆肥微生物群落组成的影响
由图3可知,在细菌门水平上,不同处理得到12个相对丰度>1%的细菌菌门,其中优势菌门为放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)和厚壁菌门(Firmicutes),共占到全部菌门相对丰度的87.44%~89.51%。CK~C5处理的绿弯菌门(Chloroflexi)相对丰度分别为12.69%、15.87%、13.93%、17.85%、16.39%和26.87%,变形菌门(Proteobacteria)和厚壁菌门(Firmicutes)相对丰度较CK均有所降低,以C3处理最低,分别为17.56%和11.76%。(图3-A)。在属水平上,共有11个优势细菌属(相对丰度>2%),其中相对丰度排名前三的细菌属分别为norank_f__norank_o__Actinomarinales、unclassified_f__Intrasporangiaceae和沉积岩杆菌属(Ilumatobacte)。相较于CK,生物炭处理中的norank_f__norank_o__Actinomarinale相对丰度下降了0.03~0.58个百分点,unclassified_f__Intrasporangiaceae相对丰度增加了0.29~0.59个百分点,以C4处理相对丰度最大(图3-B)。
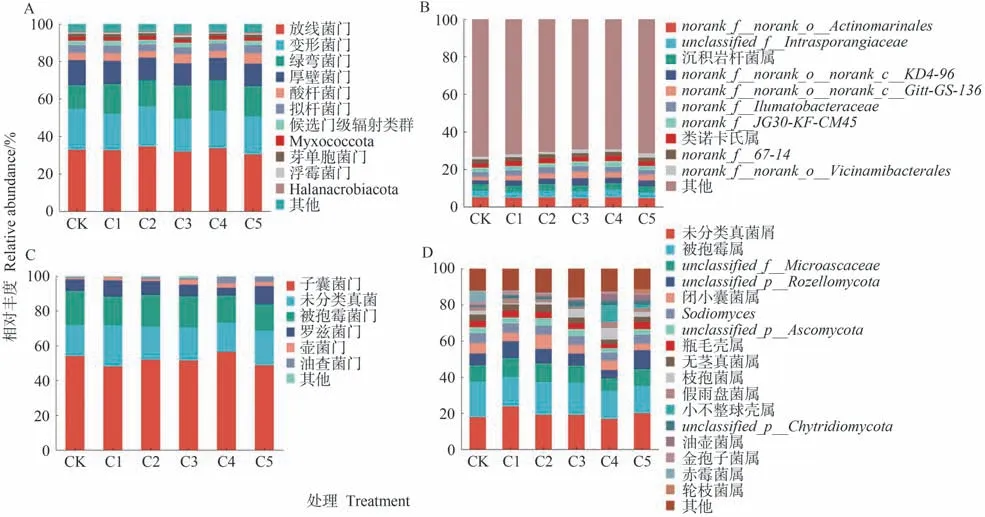
图3 生物炭对蚯蚓原位堆肥细菌和真菌群落组成的影响Fig.3 Effect of biochar on the relative abundance of microorganisms of in-situ vermicomposting at the phylum and genus level
在真菌菌落中,不同处理检测出7个相对丰度>1%的真菌菌门,其中优势菌门为子囊菌门(Ascomycota)、被孢霉菌门(Mortierellomycota)和罗兹菌门(Rozellomycota)。各处理中子囊菌门(Ascomycota)相对丰度为48.05%~56.43%。相较CK,生物炭的添加使被孢霉菌门(Mortierellomycota)相对丰度减少。除C4处理外,其他处理的罗兹菌门(Rozellomycota)相对丰度均高于CK。此外,随着生物炭添加比例的增加,油壶菌门(Olpidiomycota)作为潜在的优势菌门,其相对丰度在C4和C5处理中均高于CK(图3-C)。在属水平上,共有18个优势真菌属(相对丰度>2%),其中相对丰度排名前三的真菌属为未分类菌属(unclassified_k__Fung)、被孢霉属(Mortierella)和罗兹菌属(Rozellomycota)。相较CK,生物炭处理降低了被孢霉属(Mortierella)相对丰度,降幅为1.4~4.09个百分点。除C4处理外,其他处理的未分类真菌属(unclassified_k__Fung)和罗兹菌属(Rozellomycota)相对丰度较均高于CK(图3-D)。
2.5 相关指标的冗余分析
将细菌和真菌门、属水平相对丰度前10的种群与基质理化性质进行RDA分析。结果表明(图4),基质理化性质对细菌群落丰度解释度为40.23%(门)和32.58%(属),真菌群落丰度解释度为60.54%(门)和40.39%(属)。
在门水平上,影响细菌群落相对丰度的主要环境因子是全磷(TP)、速效钾(AK)和速效磷(AP)。其中,放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)相对丰度与TP、AP、AK含量呈正相关,变形菌门(Proteobacteria)相对丰度与TP、AP、AK含量呈负相关(图4-A)。影响属水平细菌群落相对丰度的主要环境因子是全氮(TN)、速效钾(AK)和全磷(TP)。沉积岩杆菌属(Ilumatobacte)、unclassified_f__Intrasporangiace相对丰度与AK含量呈正相关,norank_f__norank_o__Vicinamibacterales相对丰度与AK含量呈负相关(图4-B)。
对门水平的真菌群落变化产生关键作用的环境因子为全氮(TN)、速效氮(AN)和速效钾(AK)。其中,子囊菌门(Ascomycota)相对丰度与TN、AN和AK含量呈正相关,被孢霉菌门(Mortierellomycota)、罗兹菌门(Rozellomycota)相对丰度与AK、AP含量呈负相关(图4-C)。影响属水平真菌群落相对丰度的主要环境因子是全氮(TN)、速效氮(AN)和速效钾(AK)。枝孢菌属(Cladosporiu)相对丰度与TN含量呈正相关,瓶毛壳属(Lophotrichus)相对丰度与TN含量呈负相关,unclassified_p__Rozellomycota相对丰度与AK含量呈负相关(图4-D)。
2.6 蚯蚓原位堆肥与不同比例生物炭对番茄品质和产量的影响
由表5可知,生物炭处理整体显著增加了番茄植株地下部生物量(P<0.05)。与CK相比,C1~C5处理植株地下部生物量整体显著增加16.84%~112.19%。随着生物炭添加比例的增大,番茄植株地下部生物量呈先增大后减小趋势。其中,C2处理番茄植株地下部生物量达到最大值。同时,生物炭处理整体显著提高了番茄的产量(P<0.05)。与CK相比,C1~C5处理番茄产量增加11.89%~36.36%,以C3处理产量最高(9 411.72 kg·667m-2)。
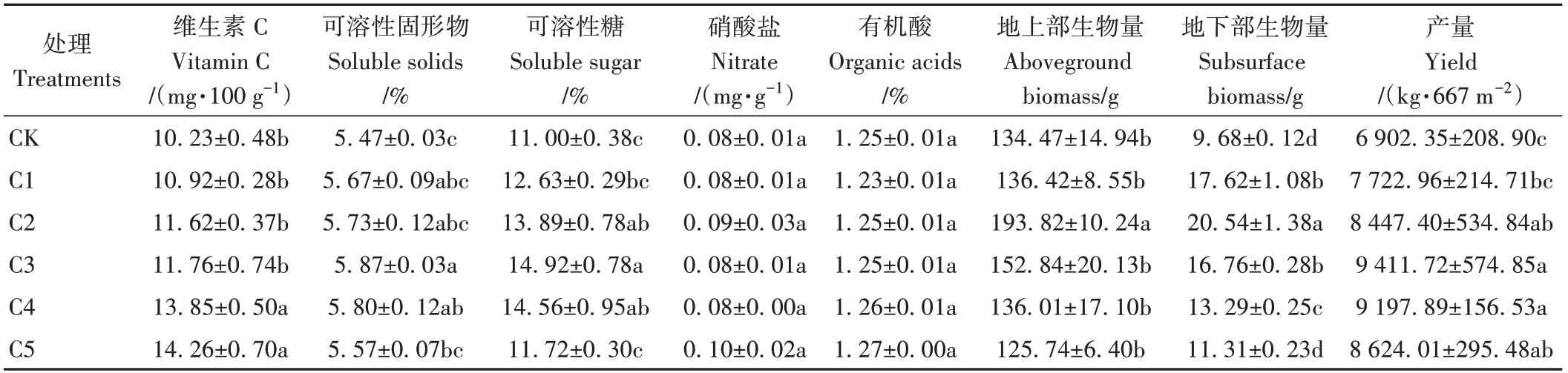
表5 蚯蚓原位堆肥与不同比例生物炭对番茄品质及产量的影响Table 5 Effect of in-situ vermicomposting with different ratio of biochar on quality and yield of tomatoes
添加生物炭均可促进果实品质相关参数的增加。与CK相比,C1~C5处理Vc、可溶性固形物和可溶性糖含量分别增加6.74%~39.39%、0.10~0.40个百分点和0.72~3.92个百分点。随着生物炭添加比例的增大,番茄Vc含量呈持续上升趋势;可溶性固形物和可溶性糖含量呈先增大后减小趋势,二者含量均以C3处理最高,分别为5.87%和14.92%。
2.7 蚯蚓原位堆肥与不同比例生物炭对番茄果实代谢的影响
为了更好地了解蚯蚓原位堆肥与生物炭对番茄果实代谢物的影响,在CK和C3处理之间进行了代谢物分析,通过偏最小二乘法判别分析(partial least squares discriminant analysis,PLS-DA)图来评估植物组织样品之间的差异,结果显示(图5),PC1的贡献率为30.70%,PC2的贡献率为29.50%。从图中易看出C3与CK间分离趋势明显,且因变量拟合指数(R2Y)>0.9,模型预测指数(Q2)>0.5,表明处理之间存在显著差异,模型具备较好的稳定性,获得的数据可继续进行后续分析。此外,潜在结构判别分析(orthogonal partial least squares discriminant analysis,OPLS-DA)建模的监督正交投影也揭示了两样本的显著差异。
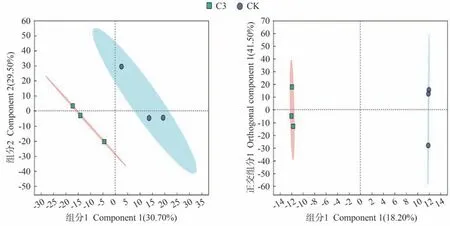
图5 番茄果实的LC-MS评价(n=3)Fig.5 Evaluation of LC-MS of tomato fruits (n=3)
由图6可知,通过变量重要性投影(variable importance in projection,VIP)≥1、P值<0.05、差异倍数(fold change,FC)>1或<1的筛选标准,在C3 vs CK中显著鉴定了16种差异代谢物,其中有14种差异代谢物上调(fold change>1),2种差异代谢物下调(fold change≤1)。由表6可知,相较于CK,C3处理中上调物质包括1-己醇阿拉伯糖基葡糖苷、L-4-羟基谷氨酸半醛、4-羟基双苯喹啉、5-甲氧基水杨酸等,下调物质包括反式祖马林和L-谷氨酰胺。
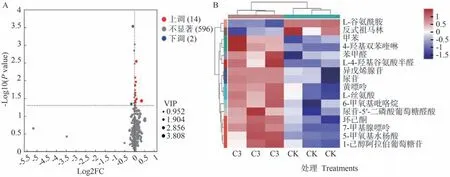
图6 番茄果实差异代谢物火山图(A)和热图(B)代谢途径分析Fig.6 Volcano map (A) and heat map (B) of tomato fruit differential metabolites
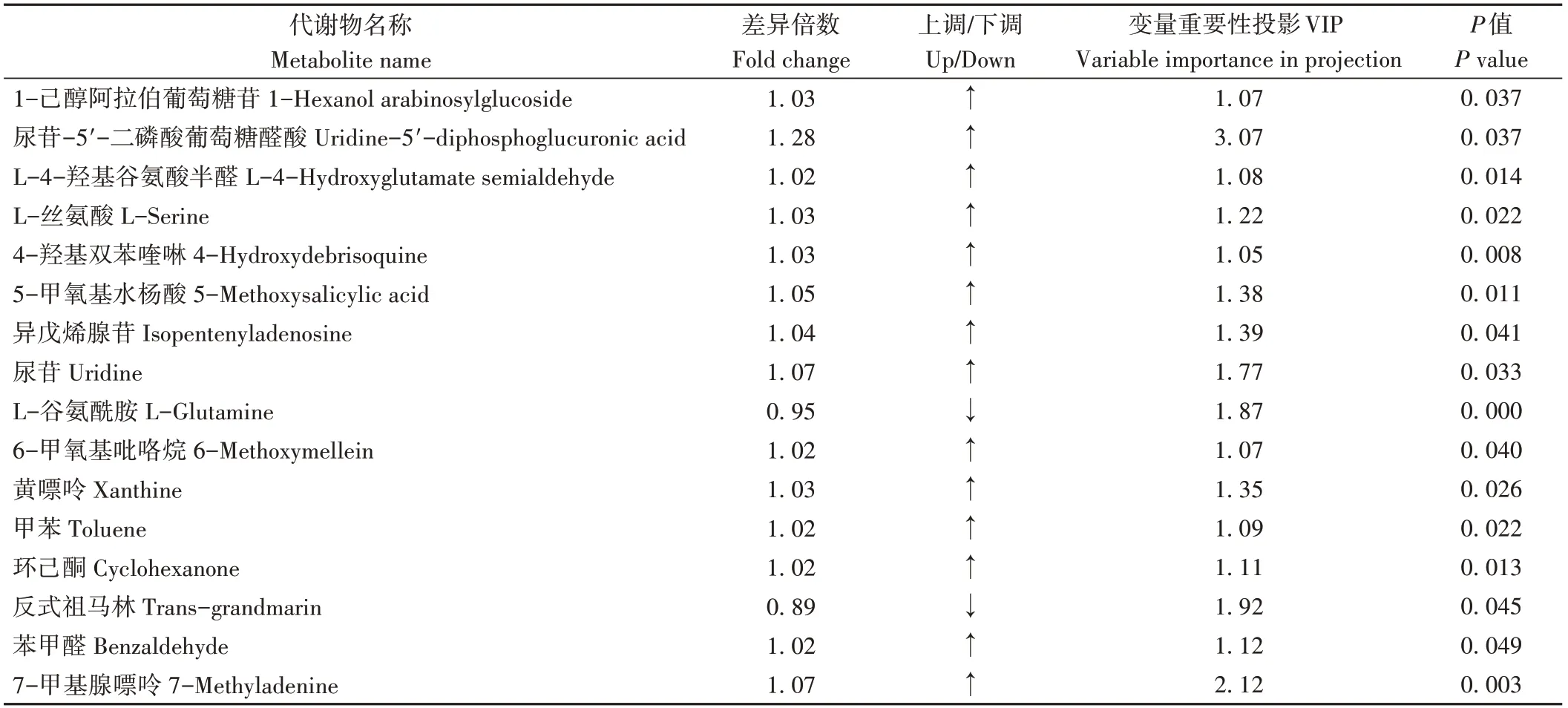
表6 番茄果实差异代谢物Table 6 Differential metabolites of tomato fruit
为进一步了解代谢途径中差异代谢物的变化,将筛选出的差异代谢物映射到KEGG通路上进行注释和通路富集分析。由图7可知,生物炭的施用对氨酰tRNA生物合成、ABC转运体蛋白(ATP-binding cassette)、乙醛酸和二羧酸代谢、嘌呤代谢和嘧啶代谢等代谢途径有极显著影响(P<0.01)。由表7可知,参与乙醛酸和二羧酸代谢、氨酰tRNA生物合成途径的差异代谢物为L-丝氨酸和L-谷氨酰胺;参与ABC转运蛋白途径的差异代谢物为L-丝氨酸、L-谷氨酰胺和尿苷;参与嘌呤代谢途径的差异代谢物为L-谷氨酰胺和黄嘌呤;参与嘧啶代谢途径的差异代谢物为尿苷。
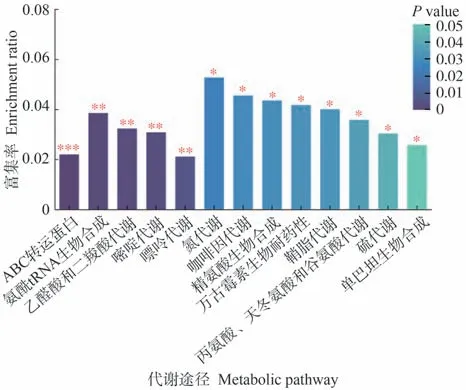
图7 番茄果实差异代谢物途径Fig.7 Differential metabolite pathways in tomato fruits
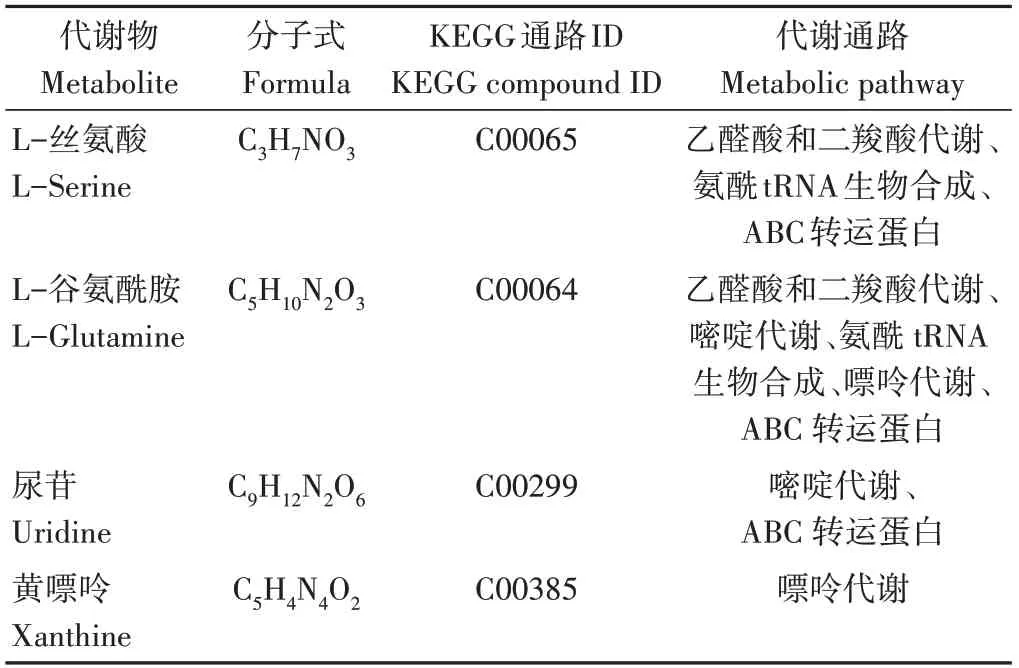
表7 差异代谢物的代谢途径归属Table 7 Metabolic pathway attribution of differential metabolites
3 讨论
研究表明,容重在0.1~0.8 g·cm-3、总孔隙度在54%~96%、气水比在0.25~0.50范围内的基质最适宜植物生长[20]。本研究所有处理的容重和总孔隙度均满足上述要求。生物炭的施入整体显著提高了蚯蚓原位堆肥中的有机质和养分含量,原因可能是生物炭较高的比表面积和吸附能力能固持蚯蚓原位堆肥中的氮、磷、钾离子,减少养分的流失,增加其可利用养分的总量,且生物炭能够延缓养分的释放,使肥效变缓,补充作物后期生长发育所需的养分[21-23]。但随着生物炭施用量的增加,蚯蚓原位堆肥养分含量呈先上升后下降的趋势,并且酶活性的变化趋势与之相似,其潜在原因是过量生物炭会吸附更多酶分子,抑制酶促反应进行,导致酶活性降低[24-25]。
适量生物炭对蚯蚓原位堆肥中的细菌丰富度有显著影响,且细菌α多样性指数大于真菌,这归因于生物炭结构疏松多孔,自身呈弱碱性,配施蚯蚓原位堆肥能够刺激微生物的代谢,促进细菌群落多样性的提升,这与程扬等[26]的研究结果相吻合。赤子爱胜蚓肠道内的细菌类群主要为放线菌门、绿弯菌门和厚壁菌门等,上述菌群也被认为是蚯蚓堆肥中常见的优势微生物[27-28]。这与本研究的结论相似,但各处理间菌群的相对丰度存在差异。一方面,绿弯菌门是一种兼性厌氧菌,可通过反向三羧酸循环来固定碳、氧化硫化物和Fe2+等物质,促进土壤养分循环和碳固定[29],添加生物炭提高了绿弯菌门的相对丰度,这表明生物炭能增加蚯蚓原位堆肥的碳汇,提升有机碳的固持能力。另一方面,生物炭的添加降低了变形菌门和厚壁菌门的相对丰度。变形菌门作为细菌中最大的门类,包括如T.Escherich 1885、沙门氏菌等在内的诸多病原菌,施加生物炭会抑制变形菌门活性,进而降低相关病原菌的相对丰度[30]。此外,Cleveland等[31]发现土壤稳定碳含量的增加会导致γ-变形菌门和厚壁菌门相对丰度的减弱。真菌类群中的优势菌门为子囊菌门,其次是被孢霉菌门和罗兹菌门。子囊菌门的主要功能是参与土壤有机质分解,尤其具有分解木质纤维素的能力,因此子囊菌门相对丰度的提高有利于有机质的矿化和养分释放[32]。被孢霉菌门中含有一些具有溶磷作用的真菌种群,可以溶解土壤中难以利用的磷元素[33]。此外,随着生物炭施用量的增加,油壶菌门的相对丰度提高,这可能是由于生物炭中某种微量物质的积累刺激了油壶菌门的繁殖,但具体原因有待进一步研究。
蚯蚓原位堆肥中添加生物炭提高了番茄的产量和品质。生物炭可加速蚯蚓原位堆肥的腐殖化过程,生物炭的表面也可以被堆肥中的微生物群落和腐殖质氧化,两者的协同作用增加了基质中可用营养物质的数量,促进了番茄植株的生长,改善了番茄的产量和品质[34]。然而,随着生物炭添加比例的增大,番茄产量、可溶性固形物和可溶性糖含量呈先增大后减小趋势,以C3处理最高。因此,生物炭对作物增产效应存在峰值[35]。此外。本研究通过LC-MS测定番茄果实代谢物,结果表明,当生物炭添加比例为3%时,KEGG通路富集产生了5条差异极显著代谢途径(P<0.01),以氨酰tRNA生物合成代谢富集率最高。L-丝氨酸和L-谷氨酰胺参与了此过程,丝氨酸作为生糖氨基酸可参与糖异生途径转变为葡萄糖,而L-丝氨酸又是由糖酵解中间体3-磷酸甘油酸酯通过三个催化步骤从头合成的;谷氨酰胺可经过代谢转化为三羧酸循环的中间产物α-酮戊二酸,进而实现氨基酸与糖类之间的转化。此外,本研究还确定了尿苷-5'-二磷酸葡萄糖醛酸、1-己醇阿拉伯葡萄糖苷等差异代谢物。尿苷-5'-二磷酸葡萄糖醛酸可在UGlc AE酶的作用下转化生成尿苷二磷酸半乳糖醛酸,进入高尔基体内参与果胶的生物合成。同时,它还可生成尿苷二磷酸木糖,间接形成木酮糖[36]。这些物质的转化可能是C3处理糖类物质含量显著高于CK的主要原因(表5)。综上,生物炭的施用可能促进了果实中氨基酸与糖类之间的转化,从而提高了果实品质。
4 结论
本研究结果表明,生物炭可提高蚯蚓原位堆肥中的养分含量和酶活性,优化蚯蚓堆肥菌群结构,进而达到改善番茄品质、提高番茄产量的目标。其中,当生物炭替代比例为3%时,蚯蚓原位堆肥中的全磷、速效磷、速效钾含量及蔗糖酶和碱性磷酸酶活性最高。上述营养状况的改变增加了细菌群落的丰富度和多样性,促使绿弯菌门(Chloroflexi)相对丰度提高,变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和被孢霉门(Mortierellomycota)相对丰度降低。此外,LC-MS分析发现,生物炭通过改变番茄果实尿苷-5'-二磷酸葡萄糖醛酸、L-丝氨酸、L-谷氨酰胺等的代谢水平而影响果实中糖类物质含量,从而保证其品质与产量保持在高水平。
