雅鲁藏布江上游高寒草甸地上生物量与多样性沿海拔梯度的变化特征
2024-03-08王晓丽马玉寿王彦龙秦金萍董瑞珍
张 敏,王晓丽,马玉寿,王彦龙,秦金萍,董瑞珍
(青海大学 青海省畜牧兽医科学院,青海省高寒草地适应性管理重点实验室,西宁 810016)
植物群落特征及其物种多样性在群落生态学研究中占有重要地位[1]。物种多样性可以表述群落的复杂程度和物种多少,是反映群落特征的重要指标之一[2]。进行群落物种多样性研究不仅能更好地反映群落在组成、结构、功能和动态等方面的异质性,也可反映不同自然地理条件与群落的相互关系及其发展变化[3]。植物群落结构是草地生态系统的重要组成部分,对研究草地类型、物种组成和结构具有重要意义[4]。高寒草甸是青藏高原主要的草地类型之一,是高寒草地生态系统的重要组成部分。在全球重视生态环境健康的大背景下,地上生物量和物种多样性作为植物群落的重要指标,促使其成为生态学研究的热点问题。根据目前已有研究,草地地上生物量与海拔间主要表现为无显著关系、先降低后增加、先增加后降低、正相关和负相关5种关系。物种多样性随海拔梯度的变化主要有与海拔高度负相关、与海拔高度正相关、在中等海拔高度最大、在中等海拔高度最小和无规律变化5种模式。在物种多样性与地上生物量的相互关系表现为单峰关系、正相关关系和负相关关系。例如潘晓婷等[5]和刘冲等[6]研究中地上生物量与多样性指数呈显著正相关关系,随着海拔上升,物种多样性指数均呈先上升后下降再上升的趋势,地上生物量呈先下降后上升的变化特征。张静等[7]研究发现草本层物种丰富度在海拔梯度上呈明显的“偏锋”格局。王金兰等[8]和Wilson等[9]研究表明地上生物量随海拔升高呈先显著增加后显著降低的变化趋势,多样性指数和丰富度指数均随海拔升高呈先增加后减小的变化趋势,多样性指数与地上生物量呈显著正相关。雅鲁藏布江作为青藏高原最重要的河流,研究雅鲁藏布江流域草地资源对保护青藏高原生态环境具有重要意义。目前,国内外对该区域不同海拔梯度下植物丰富度、植物物候、植被覆盖度变化等方面对植物多样性开展相关研究,但研究对象多基于林下草地、沙化地区、公路沿线及整体草地群落,主要以雅鲁藏布江上游高寒草甸为对象,分析不同海拔下物种多样性和地上生物量及两者之间的相互关系的研究较少[10-12]。鉴于此,本研究以雅鲁藏布江上游高寒草甸为研究对象,分析地上生物量和物种多样性沿海拔变化的特征及环境因子对地上生物量和物种多样性的影响,以期为雅鲁藏布江上游植物资源的合理利用和物种多样性保护提供依据。
1 材料和方法
1.1 研究区概况
日喀则地区位于青藏高原西南部,南北地势较高,其间为藏南高原和雅鲁藏布江流域,雅鲁藏布江是青藏高原最重要的河流,同时也是中国重要的国际河流之一,其发源于西藏自治区日喀则市仲巴县海拔5 590 m 的杰马央宗冰川,由西向东横贯西藏南部,物种丰富。研究区位于西藏自治区日喀则地区雅鲁藏布江流域(图1),共12个研究样点(表1),经 度83°44′18″—89°32′36″E,纬 度27°58′45″—29°52′23″N,海拔3 774~4 589 m,年均降水量149.36~293.99 mm,年均气温(-3.01)℃~5.14℃。雅鲁藏布江上游流域高寒草甸植物资源丰富,主要物种有高山嵩草(Kobresia pygmaea)、藏北嵩草(K.littledalei)、矮生嵩草(K.humilis)和华扁穗草(Blysmus sinocompressus)等。
图1 研究区域Fig.1 Map of the area where this study was carried out
1.2 植被调查
于2020年8—9月在植物生长高峰期进行植物群落调查[13]。沿海拔梯度设置12个样点,每个样点3个重复,调查点分布信息如表1所示。高寒草甸植被均匀,草地平坦。
年均降雨量和年均气温数据来自IWMI在线气候服务平台(http://wcatlas.iwmi.org/Default.asp)。
植被盖度:选择群落结构和组成分布均匀的区域,每个采样点随机3个50 cm×50 cm 的样方,将样方框平铺于草地,5 cm×5 cm 作为1个单位,将样方框分为100个单位,测定总盖度和各物种的分盖度。植物经济功能群分为禾本科、莎草科、阔叶型可食草和毒害草4种[14]。
植物株高:在样地内随机选取同种植物10株,利用直尺测量其自然高度,样地内不足10株的全部测量,为减小测量误差,将直尺零刻度线至边缘空白处去除,求平均值作为该物种的株高。
地上生物量:分种类齐地面剪取样方内各物种的地上部分,剪下后称量各物种的鲜重,然后带回室内,于75℃下烘干至恒重,记录干重,作为各物种的地上生物量。地上总生物量采用各物种地上生物量干重之和计算。
1.3 植物群落多样性分析[15]
重要值(I P):I P=(相对高度+相对盖度+相对生物量)/3
Shannon-Weiner指数(H):H=-∑P i×lnP i
Simpson指数(D):D=1-∑P i2
Pielou指数(J):J=H/lnS
Patrick指数(R):R=S式中:P i为物种i的相对重要值;S为所调查样方内物种总数。
1.4 数据分析与处理
1.4.1 回归分析
线性和非线性回归分析。采用单因素方差分析(ANOVA)进行F检验,模型成立的前提条件是通过F检验,即对应的P值小于0.05。第一步:如果上表显示通过F检验,则说明模型成立,反之说明模型完全无意义;第二步:如果未通过F检验,进行其他方式的拟合。模型质量分析和模型对比判断。第一步,分析模型拟合情况,即通过R2分析模型拟合情况;第二步,对拟合模型进行多次分析时对比AIC和BIC 值,此两值越低越好,以此综合判定最佳方程。
根据回归分析研究X对Y的影响关系。如果P值小于0.05或0.01,则说明X对Y有影响关系。对相关指标进行回归分析,不同回归模型的AIC 和BIC指数如表2所示。海拔与地上生物量最优拟合方程为二次项回归方程。海拔与Shannon-Weiner指数和Patrick 指数最优拟合方程为线性回归方程。海拔与Simpson指数和Pielou指数最优拟合方程为三次项回归方程。地上生物量与Shannon-Weiner指数和Simpson指数最优拟合方程为三次项回归方程。地上生物量与Patrick指数和Pielou指数最优拟合方程为线性回归方程。

表2 模型判断指标Table 2 Indicators for model judgment
1.4.2 数据处理
用Excel 2019 对数据进行记录整理。利用SPSS 26.0 进行单因素方差分析(one-way ANOVA)、线性和非线性回归分析和Pearson相关性分析:(1)地上生物量和多样性指标数据进行单因素方差分析(one-way ANOVA);(2)对群落的地上生物量和多样性指数进行回归分析:地上生物量、Shannon-Weiner指 数、Simpson 指 数、Pielou 指 数、Patrick指数与海拔之间的关系,地上总生物量与多样性指数之间的关系;(3)对地上生物量、多样性指数和环境因子进行Pearson相关性分析。采用Origin 2021进行绘图。
2 结果与分析
2.1 高寒草甸植物群落组成
在西藏日喀则地区雅鲁藏布江流域共调查12个样地,调查物种225种,主要物种如表3所示。高寒草甸优势种以莎草科植物为主,包括高山嵩草(K.pygmaea),藏北嵩草(K.littledalei),矮生嵩草(K.humilis)和华扁穗草(B.sinocompressus)等。高山嵩草的主要伴生种包括华扁穗草、刺鳞蓝雪花(Ceratostigma ulicinum)、丝颖针茅(Stipa capillacea),藏北嵩草和矮生嵩草的主要伴生种为青藏薹草(Carex moorcroftii),华扁穗草的主要伴生种为尾穗嵩草(K.cercostachy)。

表3 高寒草甸主要物种Table 3 Main species of the alpine meadows
2.2 海拔对高寒草甸地上生物量和物种多样性的影响
2.2.1 地上生物量
地上生物量随海拔变化如图2所示。在回归分析中,最优拟合曲线为二项式。海拔对地上生物量的解释度为22%,虽然地上生物量的二次回归曲线拟合呈U 形,但二者没有显著相关性(P>0.05),不能说明随海拔升高地上生物量先降低后增大的变化趋势。地上生物量最大值为4 542 m,最小值为4 335 m。
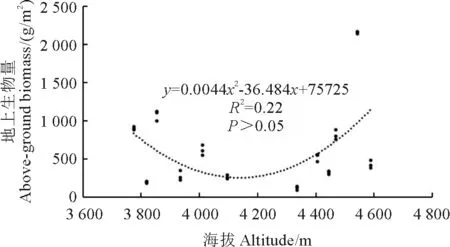
图2 地上总生物量与海拔的关系Fig.2 Total above-ground biomass as a function of the altitude
2.2.2 群落多样性
由图3 可知,高寒草甸植物群落Shannon-Weiner指数(H)和Patrick指数(R)的最大值均处于低海拔高度,随海拔升高呈明显的下降趋势。在回归分析中,海拔与Shannon-Weiner指数(H)和Patrick指数(R)的最优拟合为线性回归,显著相关(P<0.05),说明Shannon-Weiner指数(H)和Patrick指数(R)随海拔升高呈线性降低。海拔与Simpson指数(D)和Pielou指数(J)最优拟合曲线为三项式,虽然呈S形,但是P>0.05,不能解释随海拔升高呈现的变化趋势。

图3 不同海拔梯度下植物群落多样性指数变化情况Fig.3 Diversity indices of plant communities at different altitudes
2.3 高寒草甸地上生物量与植物群落多样性的关系
地上生物量和多样性关系如图4所示。Shannon-Weiner指数(H)、Simpson指数(D)与地上生物量回归分析中拟合最优为三次多项式,Pielou指数(J)与地上生物量拟合最优为一次函数,均显著相关(P<0.05)。Shannon-Weiner 指 数(H)、Simpson指数(D)和Pielou指数(J)随着地上生物量变化呈减小趋势,Shannon-Weiner指数(H)对地上生物量的解释达到70%。Patrick指数(R)与地上生物量最优拟合为一次函数,但是P>0.05,不能解释随Patrick指数(R)增加地上生物量呈线性下降的变化趋势。

图4 地上生物量和物种多样性的关系Fig.4 Relationship between above-ground biomass and species diversity
2.4 地上生物量、多样性指数与环境因子的关系
不同海拔下地上生物量、多样性和环境因子的相关性分析如表4 所示。地上总生物量与Shannon-Weiner指数(H)、Simpson指数(D)、Pielou指数(J)呈显著负相关(P<0.01)。Shannon-Weiner指数(H)与海拔和气温之间呈显著负相关。Shannon-Weiner指数(H)和海拔之间的相关系数值为-0.385,P<0.05。

表4 高寒草甸地上生物量、多样性指数与环境因子的Pearson相关性分析Table 4 Pearson correlation analysis of above-ground biomass,diversity index,and environmental factors of alpine meadow
Shannon-Weiner指数(H)和气温之间的相关系数值为0.602,P<0.01。Simpson指数(D)与海拔、降水和气温之间无显著性。Pielou 指数(J)与降水之间呈显著正相关(P<0.05)。Patrick 指数(R)与海拔、气温之间的具有相关性。Patrick指数(R)和海拔之间的相关系数值为-0.343,P<0.05,说明Patrick 指数(R)和海拔之间显著负相关。Patrick指数(R)和气温之间的相关系数值为0.588,P<0.01,Patrick指数(R)和气温呈显著正相关关系。
总生物量与海拔、降水和气温之间无显著性,相关系数值分别是0.190,-0.079,-0.192,均接近于0,并且P>0.05,表明总生物量与海拔、降水和气温之间均无相关关系。
3 讨论
3.1 雅鲁藏布江上游高寒草甸植物群落特征
高寒草甸是青藏高原主要的草地类型之一,是高寒草地生态系统的重要组成部分。雅鲁藏布江流域中上游的高寒草甸主要以高山嵩草,藏北嵩草为主[16]。本研究中雅鲁藏布江上游主要植物包括高山嵩草,藏北嵩草,矮生嵩草和华扁穗草。主要伴生种包括华扁穗草、刺鳞蓝雪花、丝颖针茅,青藏薹草,尾穗嵩草等。崔恒心等在调查中表明西藏高寒草甸主要植物有高山嵩草、矮生嵩草、藏北嵩草、华扁穗草、圆穗蓼(Bistorta macrophylla)和黑褐穗薹草(Carex atrofusca)等[17],可能由于调查局限性,未能涉及圆穗蓼和黑褐穗薹草等植被类型。
本研究调查显示样地中优势种主要以莎草科植物为主,这与上述研究结果吻合,尽管杂类草的入侵会影响莎草科植物的优势地位,但其在植物群落中依然保持着较高的生物量,这可能是因为莎草科在植物群落中占据着优势地位,也可能由于高寒草甸在长期群落演替过程中形成较稳定的生态系统[18-22]。
3.2 不同海拔梯度下雅鲁藏布江上游高寒草甸地上生物量和多样性变化特征
海拔对植物的影响是热量、水分、养分和光照等多个环境因子的综合效应,决定着植物种的分布格局,研究表明草地中物种地上生物量随着海拔变化有5种趋势:无显著关系、先降低后增加、先增加后降低、正相关和负相关的变化特征[23]。白晓航等[24]和牛钰杰等[25]在研究中指出在一定海拔范围内,不同物种对海拔的响应呈现多种变化趋势,同一地区出现多种变化趋势并不矛盾,这可能与物种适应特征和分布范围不同有关。郭建兴等[26]和常风[27]研究结果表明在草地生态系统中植被地上生物量随海拔变化先升高后降低,研究者认为是区位、气候条件和人类活动等因素共同作用所导致,经度、纬度和植被盖度也是导致地上生物量变化的原因。也有研究认为水分是造成地上生物量变化的主要原因,其次土壤养分也有可能影响地上生物量[28],已有研究表明因特定生态环境和研究尺度的差异,研究人员对地上生物量与海拔梯度的关系仍没有统一的定论。在本研究中地上总生物量与海拔之间无显著关系,地上生物量与海拔、气温和降水无显著关系,可能与人为活动有关。雅鲁藏布江流域水源充足,草地资源丰富,畜牧业得到良好发展,沿线的放牧活动会降低地上生物量,导致不同海拔地上生物量呈无规律变化;其次土壤理化性质也可能影响地上生物量,雅鲁藏布江两岸良好的水热条件可以满足较多物种的生存需求,但对于植物良好生长发育的土壤养分需求还未可知,因此该区域植被和土壤理化性质的关系也是将来需要研究的内容。本研究区域跨经度较广,植被的地域性特征较强,导致地上生物量表现出不规律特征。
物种多样性沿海拔梯度的变化格局一直是生物多样性研究的一项重要内容。贺金生等[29]学者指出随海拔高度的变化,物种多样性有5种模式:与海拔高度负相关,与海拔高度正相关,在中等海拔高度最大,在中等海拔高度最小和无规律变化。周芸芸等[30]研究发现多样性指数随海拔升高降低。常学向等[31]的研究中多个多样性指数趋势表现出不一致的情况,卢慧等[32]和林红[33]也发现多样性指数会随海拔梯度的增加表现出正相关及先增加后减少等不同变化趋势,综合多位学者研究结果,均认为海拔梯度会造成降水、气温等环境因子变化[34],加之人类干扰多种因素导致物种分布格局发生相应变化。本研究发现海拔与多样性呈负相关关系,Shannon-Weiner指数和Patrick指数的最大值均处于低海拔高度。在海拔梯度下,环境异质性和对气候的敏感程度和人类活动的扰乱都会造成不同梯度下物种分布发生变化[35],西藏地广人稀,在雅鲁藏布江上游调查过程中发现人为干扰因素并不会对草地生态系统造成严重破坏,虽然会对地上生物量造成影响,但减少物种多样性可能性较低。在本研究中多样性与气温、降水呈显著正相关关系,多样性随气温升高表现出增加趋势,说明水热条件越优越,越适宜更多物种生存,海拔越高,水热条件越差,因此物种多样性越低。
3.3 雅鲁藏布江上游高寒草甸地上生物量和植物群落多样性的关系
在物种多样性与地上生物量的相关研究中,二者的主要关系有:单峰关系[36]、正相关关系[37]、负相关关系[38]。本研究中地上生物量与物种多样性呈负相关关系,物种丰富的植物群落反而表现出低生产力。这种关系的出现可能与群落组成的差异有关,有研究表明优势物种能驱动地上生物量变化[39],在本研究中莎草科植物作为优势种,其绝对的竞争优势导致物种多样性减小、地上生物量增加。地上生物量和物种多样性之间的关系与环境干扰也有关系,当环境对一者起促进作用,对另者起抑制作用,则呈现出负相关关系,若同时起促进作用则呈现正相关关系。水热条件的变化所引起的物种选择、资源竞争、生境变化是影响物种多样性和生产力关系的重要因素。王娟等[40]提出地上生物量与物种多样性之间存在显著的负相关关系,认为增加人为管理措施后,改善土壤养分条件是植物能获取良好生长条件的主要原因。然而刘哲等[41]研究发现地上生物量与物种多样性呈S形曲线,认为放牧是引起该趋势的重要因素。王长庭等[42]认为在青藏高原地上生物量与物种多样性主要呈单峰变化,认为此现象符合“中间高度膨胀”观点。青藏高原物种多样性和地上生物量关系复杂多样,造成的原因可能与空间尺度的分布有关,物种资源竞争、生境变化和人为活动等因素均可影响二者之间的关系[43-44],对多样性和地上生物量的关系仍需进行研究。
4 结论
通过对雅鲁藏布江上游高寒草甸植被群落的调查,结果表明地上生物量与海拔变化不相关,Shannon-Weiner指数(H)和Patrick指数(R)随海拔升高呈下降趋势,地上生物量和物种多样性表现为负相关关系。海拔与多样性呈负相关,气温、降水与多样性呈正相关。海拔、气温和降水与地上生物关系不显著。研究结果对维护青藏高原物种多样性和草地生态保护具有一定的意义。
