西番莲花芽分化过程观察及花生长模型的拟合
2024-03-08田青兰周俊妞吴艳艳刘洁云黄伟华张英俊谢文连韦广谭牟海飞
田青兰,周俊妞,吴艳艳,刘洁云,黄伟华,张英俊,谢文连,韦广谭,牟海飞
西番莲花芽分化过程观察及花生长模型的拟合

广西农业科学院生物技术研究所,南宁 530007
【目的】明确我国南方生态区主栽西番莲品种的花芽分化过程,建立西番莲花芽形态分化的花生长模型,为西番莲促花保花提供参考。【方法】以紫果类主栽品种‘台农1号’(TN)和黄果类广西主推新品种‘壮蜜05’(同‘壮乡蜜宝’,MB)为试验材料,通过扫描电镜和石蜡切片解剖观察两品种花芽分化过程,观察茎顶端分生组织以下各节位分化进程,并测量各节位花芽的第一苞片长、叶片长、卷须长;通过将茎顶端分生组织以下第7节记为初始标记节位,此时花芽为第一苞片分化期,跟踪调查标记后0—16 d花芽分化进程及标记后至开花的第一苞片长、花蕾长等5个花形态指标的动态变化,构建花生长模型。【结果】供试的两个西番莲品种花芽分化的主要过程包括第一苞片形成期、额外苞片形成期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期和副花冠形成期等,其中第一苞片形成到雌蕊形成历时10—12 d,TN较MB进程快1—2 d,雌蕊形成是西番莲花芽发育成功的重要标志,此时花芽第一苞片长度为3—4 mm,第一苞片形成到开花历时36—44 d,TN较MB进程快3—4 d。茎顶端分生组织以下第4—5节位可见卷须分化,第6节位花原基分化,着生于卷须旁侧,第7节位可见第一苞片分化,第9节位出现腋生营养分生组织,单独位于卷须和花芽内侧,第10—11节位可见腋芽形成,第14—15节位雌蕊原基分化。利用Origin软件对MB和TN的第一苞片长度、第一苞片宽度、花梗长、花蕾长、花梗和花蕾总长共5个花形态指标和标记后天数进行Logistic模型的非线性回归拟合,决定系数2为0.9524—0.9988,标准化均方根误差()为8.54%—19.62%,模型方程的拟合效果较好。根据模型参数和实际观察,苞片的长度在标记后即第一苞片形成期后11—12 d进入快速增长期,标记后24—26 d进入缓慢增长期后趋于稳定;之后花梗和萼片迅速生长,萼片长度超过苞片,花蕾和花梗总长在标记后24—25 d进入快速增长期,标记后41—42 d进入缓慢增长期到最大值后开花。【结论】西番莲的花芽与卷须紧靠,而与营养芽分开独立;西番莲花芽分化和形态分化可分为3个阶段,第一苞片形成期到雌蕊形成期,历时10—12 d;苞片生长期,历时12—14 d;花梗和花蕾生长期,历时15—17 d。在西番莲实际生产中,可通过花生长模型预测开花时间,通过苞片长度等形态指标判断花芽分化进程,为促花保花方案提供参考依据。
西番莲;花芽分化;花生长模型;形态分化
0 引言
【研究意义】西番莲()是我国南方特色果树,又名百香果[1],果实主要用于鲜食和加工。近年来西番莲种植面积和产量迅速增加,但总体呈现产量低且不稳定等突出问题。花量是产量形成的基础,目前西番莲生产上普遍存在夏季高温和冬季低温花芽分化受限问题,尤其是夏季花量减少制约了产量的增加[2-4]。因此,探明西番莲花芽分化过程和花生长模型对生产上促花保花和增产具有重要意义。【前人研究进展】植物生长到一定阶段便由叶芽生理和组织状态转化为花芽生理和组织状态,发育成花器官雏形,这个过程称作花芽分化[5]。花芽分化是植物生长从营养生长转向生殖生长的重要标志。西番莲的花芽分化包括两个阶段,第一阶段为从花原体到形成具有花器的花芽的过程,相当于一般植物的花器形成阶段;第二阶段为从具有花器的花芽至开花前的过程,相当于一般植物的花器成熟阶段[6]。如梨的花芽形态分化过程包括花序原基分化期、花蕾原基分化期、花萼原基分化期、花瓣原基分化期、雄蕊原基分化期和雌蕊原基分化期,之后进入花芽分化休眠期,直至翌年解除休眠,经历心室形成期、胚珠形成期最终发育完成[7],而西番莲的花芽无休眠期,从当年新生枝条的叶腋发出。国外有研究将紫果西番莲的花器官分化和形成分为苞片形成期、萼片分化期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期、花冠形成期、子房形成期和胚珠形成期,而花芽分化至开花约60 d[8]。张育森等[6]报道的‘台农1号’从具有花器的花芽到开花约1个月。此外,Logistic生长模型在作物生产中有广泛的应用,其中形态模型注重植物器官的形成和三维形态的产生[9],如陈扬等[10]利用有效积温定量模拟夏玉米的生长发育动态;马二磊等[11]建立甜瓜植株和果实的生长发育模型,划分甜瓜植株和果实的生长期,为甜瓜精细化和标准化栽培管理提供参考;石国朝[12]构建了冬枣花生长发育模型,为冬枣设施栽培花期管理提供了基础。【本研究切入点】目前国内关于西番莲的花芽形态分化和花器官形成过程、花形态生长模型未见报道,且对我国南方地区种植的现有品种的花芽分化进程还有待进一步研究。【拟解决的关键问题】本研究选取西番莲紫果类主栽品种‘台农1号’和广西主推黄果类新品种‘壮蜜05’为试验材料,对两品种花芽分化进程的扫描电镜和石蜡切片解剖观察;以花芽分化形态观察和第一苞片长、花蕾长等花形态指标进行Logistic生长模型拟合,以明确我国南方生态区西番莲的花芽分化过程、时间进程、形态分化和生长动态,建立花芽形态指标到分化进程到预测开花时间之间的联系。
1 材料与方法
1.1 试验设计
供试品种为主栽品种‘台农1号’(简称TN,紫色果皮)和自主选育的广西主推品种‘壮蜜05’(同‘壮乡蜜宝’,简称MB,黄色果皮,已获植物新品种权)。试验点位于广西南宁市西乡塘区石埠街道办美丽南方示范园区,试验地0—20 cm土层含全氮0.83 g∙kg-1,全磷0.86 g∙kg-1,全钾48.0 g∙kg-1,水解性氮117.0 mg∙kg-1,有效磷 75.0 mg∙kg-1,速效钾117.0 mg∙kg-1。采用单因素随机区组试验设计,共2个供试品种,每个品种设置3个小区,每小区种植10株,共30株。2021年4月20日移栽,种植行距×株距为1.8 m×1.5 m,对试验地植株统一进行常规水肥管理。
1.2 测定项目与方法
于8月4日开始选择二级蔓统一挂牌标记初始节位的位置,如图1所示初始标记节位处,由图4可知初始标记节位在生长点下第7节,每小区挂牌60个。

大图放大倍数6.7×,比例尺2 mm;小图放大倍数45×,比例尺0.5 mm
The magnification of the large image is 6.7×, the scale is 2 mm; the magnification of the small image is 45×, the scale is 0.5 mm
图1 ‘台农1号’(左)和‘壮蜜05’(右)的初始标记节位
Fig. 1 Initial mark nodes for Tainong No. 1 (left) and Zhuangmi 05 (right)
1.2.1 花芽分化进程电镜扫描观察 于标记当天取样,每小区采集茎尖及下部不同发育阶段的花芽各4个样品,采后立即投入电镜固定液中,于常温固定2 h后置于4 ℃冰箱保存。在奥林巴斯SZ61解剖镜下将茎尖分生组织剥离,将不同发育阶段的花芽迅速剥开,剥离后的样品长度小于3 mm,立即投入电镜固定液中置于4 ℃冰箱保存备用。将样品经过后固定、脱水、干燥、样品导电处理喷金后,置于SU8100扫描电子显微镜下观察采图。同时,每品种采集10个茎尖,置于解剖镜下观察茎尖下各节位的花芽分化进程,并剥离出各节位的叶片、卷须及花芽,拍照并测量叶片长、卷须长、第一苞片长。
1.2.2 花芽分化进程解剖观察 于标记当天开始取样,于标记后0、2、4、6、8、10、12和16 d在每小区分别采集原标记节位的花芽6个,投入FAA固定液中固定,然后于4 ℃冰箱保存备用,样品经过脱水浸蜡、包埋、从花芽中部纵切切片、脱蜡、甲苯胺蓝染色、透明封片制作石蜡切片,置于奥林巴斯SZ61解剖镜(6.7×—45×)和奥特光学BK-FL生物显微镜(40×—200×)下拍照。
1.2.3 花生长数据的调查 从标记当天开始调查,于标记后0、2、4、6、8、10、12、16、20、24、28、32和36 d直至开花,其中TN于标记后36—41 d开花,MB于标记后39—44 d开花;将标记后8 d前的样品于解剖镜下剥离出花芽并进行拍照、测量,后期用游标卡尺于田间测量,测量第一苞片长度、宽度、花梗长、花蕾长,其中花蕾长初期为第一苞片长度,待萼片长度超过苞片长度后,花蕾长为花蕾基部到萼片顶部的长度。
1.3 Logistic方程模拟及有效性检验
采用Logistic模型对第一苞片长度、第一苞片宽度、花梗长、花蕾长、花蕾和花梗总长共5个花生长指标进行非线性回归拟合,回归方程见公式(1)。式中y表示花生长指标,a表示花生长指标的生长上限,b为基础状态参数,k为增长速率系数,e为自然对数底。参考文献[10],根据公式(2)和公式(3)分别求得最大增长速率V1和达到最大生长速率时对应的标记后天数t1。其次,根据公式(4)和公式(5)分别求得生长曲线上的拐点t2和t3,其中0—t2为生长渐增期、t2—t3为生长快增期、t3至开花为生长缓增期,并根据公式(6)求得快增期的平均生长速率V2。






参照文献[10]采用标准化均方根误差()检验模型的有效性,公式如下:

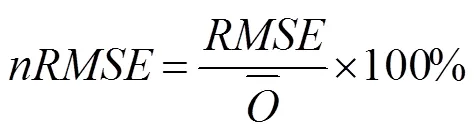
1.4 数据分析
采用Excel软件进行数据的统计、整理和作图,用Adobe Photoshop对图片进行排版,用Origin进行Logistic方程拟合。
2 结果
2.1 西番莲花芽分化过程观察
2.1.1 电镜扫描观察及解剖观察 西番莲茎顶端分生组织(图2-B1、A2)先后经历叶片和托叶的形成、卷须和花芽的形成:托叶2片位于叶片两侧(图2-A1、B1、A2、B2),卷须由卷须分生组织分化而来(图2-C1),随后从卷须旁侧形成一个花原基(图2-D1、C2),位置在叶片内侧。MB和TN的花芽分化进程一致,主要过程为:第一苞片形成(图2-E1、D2),其长度较长,第一苞片基部两侧另外2张苞片形成(图2-E1—G1、D2—F2),其长度较短;3张苞片包裹着花原基(图2-F1、G1、D2、E2),花原基分化出萼片原基(图2-H1),萼片形成(图2-I1、G2),共5张萼片,萼片内侧分化出花瓣原基(图2-J1、H2),共5片花瓣,花瓣内侧分化出雄蕊原基(图2-K1、I2),共5枚雄蕊,雄蕊内侧分化出雌蕊原基(图2-L1、J2、K2),之后雌蕊分化形成柱头、花柱(图2-M1、N1、L2、M2),在雄蕊基部外侧和花瓣基部内侧分化出花冠原基(图2-O1、N2),之后发育成丝状副花冠。
取MB和TN的茎尖及花芽组织制作石蜡切片进行纵切观察,由图3可以看出,西番莲茎顶端分生组织分化及花芽分化经历的主要时期为:叶片和托叶原基形成(图3-A)、卷须原基和花原基形成(图3-A、B)、第一苞片原基形成(图3-C)、额外的苞片原基形成(图3-D)、萼片原基形成(图3-F)、花瓣原基形成(图3-G)、雄蕊原基形成(图3-H)、雌蕊原基形成(图3-J),与扫描电镜观察结果一致。
2.1.2 西番莲茎尖下各节位电镜扫描观察及分化进程描述 取MB和TN茎尖生长点以下3 mm长度样本进行扫描电镜观察(图4),同时对采样的各节位叶片、卷须、苞片长度进行测量(表1)。以茎尖顶端分生组织为第1节位向下观察,不同节位的分化情况大致为:第2节位叶原基分化;第3节位托叶分化;第4、第5节位卷须分化及发育;第6节位花原基形成,着生在卷须旁侧;第7节位第一苞片分化,为本试验中的初始标记节位,此时MB和TN第7节位叶片长平均值分别为1.22 mm和1.24 mm;第8节位额外苞片分化及发育;第9节位出现腋生营养分生组织,单独位于卷须和花芽内侧;第10节位萼片原基分化、腋芽发育;第11节位萼片分化或萼片形成,两品种叶长平均值均为6.8 mm,卷须长3 mm左右,第一苞片长1.35—1.41 mm;第12节位萼片形成或花瓣原基分化;第13节位MB花瓣原基分化,TN雄蕊原基分化;第14节位MB雄蕊原基分化,TN雌蕊原基分化;第15节位MB雌蕊原基分化,此时MB第一苞片长平均为3.64 mm,TN第一苞片长平均为3.23 mm。此外,从第13节位开始,MB的叶长和卷须长明显高于TN,逐渐表现出品种间的差异。
‘壮蜜05’(MB):A1、B1:茎顶端分生组织;C1:卷须分生组织;D1:卷须和花芽形成;E1:第一苞片形成期;F1、G1:第2和第3苞片形成期;H1:萼片原基分化期;I1:萼片形成期;J1:花瓣原基分化期;K1:雄蕊形成期和雌蕊原基分化期;L1:雌蕊形成期;M1、N1:雄蕊、雌蕊发育;O1:副花冠分化期。‘台农1号’(TN):A2、B2:茎尖分生组织;C2:卷须分生组织和花芽原基分化期;D2、E2、F2:第一苞片和额外苞片形成期;G2:萼片形成期;H2:花瓣原基分化期;I2:雄蕊原基分化期;J2、K2:雄蕊形成期和雌蕊原基分化期;L2:雄蕊和雌蕊发育;M2:柱头和花柱分化期;N2:副花冠分化期。l:叶片;ls:托叶,SAM示顶端分生组织;tm:卷须分生组织;f:花原基;t:卷须;b1:第一苞片;b2,、b3:额外的苞片;s:萼片;p:花瓣;st:雄蕊;g:雌蕊;stg:柱头;sty:花柱;c:副花冠
Zhuangmi 05 (MB): A1, B1: Shoot apical meristem; C1: Tendril meristem; D1: Tendrils and flower bud formation; E1: First bract formation stage; F1, G1: 2nd and 3rd bract formation stages, respectively; H1: Sepal primordium differentiation stage; I1: Sepal formation stage; J1: Petal primordial differentiation stage; K1: Stamen formation stage and gynoecium primordium differentiation stage; L1: Gynoecium formation stage; M1, N1: Development of stamens and gynoecia, respectively; O1: Corona differentiation stage. Tainong No. 1(TN): A2, B2:Shoot apical meristem; C2: Tendril meristem and flower bud primordium differentiation stage; D2, E2,F2: The first and additional bracts formation stages; G2: Sepal formation stage; H2: Petal primordial differentiation stage; I2: Stamen primordial differentiation stage; J2, K2: Stamen formation stage and pistil primordial differentiation stage; L2: Development of stamens and gynoecia; M2: Stigma and style differentiation stages; N2: Corona differentiation stage. l: Leaf; ls: Stipule; SAM: Shoot apical meristem; tm: Tendril meristem; f: Flower; t: Tendril; b1: The frist bract; b2, b3: The two additional bract; s: Sepal; p: Petal; st: Stamen; g: Gynoecium; c: Corona; stg: Stigma; sty: Style; c: Corona
图2 ‘壮蜜05’(A1—O1)和‘台农1号’(A2—N2)花芽分化过程电镜扫描观察
Fig. 2 Electron microscopic scanning observation of the flower bud differentiation process of Zhuangmi 05 (A1-O1) and Tainong No. 1 (A2-N2)
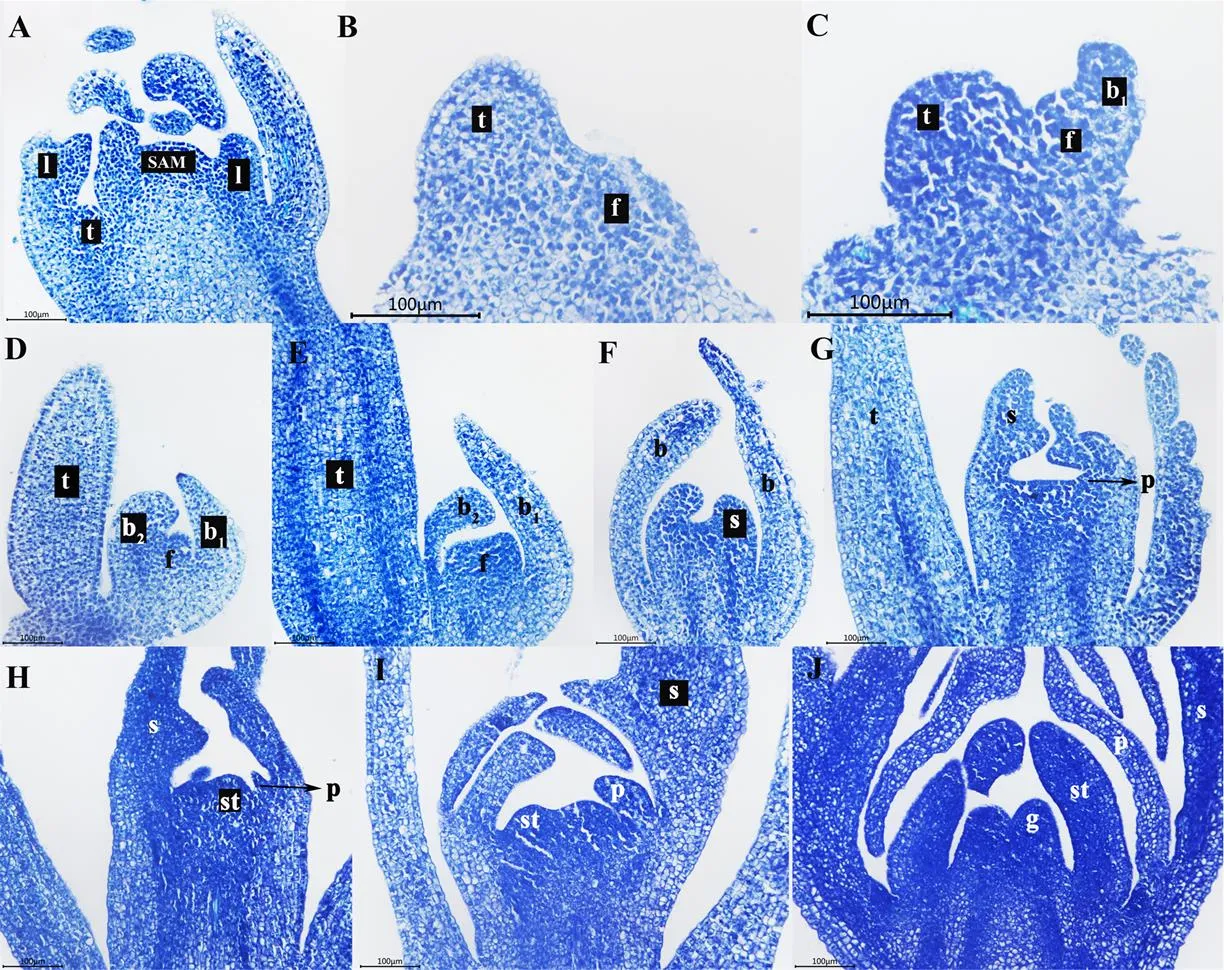
A:茎顶端分生组织SAM,卷须分化期;B:苞片原基分化期;C:第一苞片分化期;D:额外苞片分化期;E:萼片原基分化期;F:萼片分化期;G:花瓣原基分化期;H:花瓣分化期,雄蕊原基分化期;I:雄蕊分化期;J:雌蕊分化期。l:叶片;f:花原基;t:卷须;b:苞片;b1:第一苞片;b2:额外的苞片;s:萼片;p:花瓣;st:雄蕊;g:雌蕊。a—j放大倍数均为200×,比例尺均为100µm
a: shoot apical meristem (SAM), tendrils differentiation stage; b: Bract primordium differentiation stage; c: First bract differentiation stage; d: Extra bract differentiation stage; e: Sepal primordium differentiation stage; f: Sepal differentiation stage; g: Petal primordial differentiation stage; h: Petal differentiation stage, stamen primordium differentiation stage; i: Stamen differentiation stage; j: gynoecium differentiation stage. l: leaf, f: Flower, t: Tendri, b: Bract, b1: The frist bract, b2: The additional bract, s: Sepal, p: Petal, st: Stigma, g: Gynoecium. The a—j magnification is 200× and the scale is 100 µm
图3 西番莲花芽分化过程解剖观察(以‘壮蜜05’为例)
Fig. 3 Anatomical observation on flower bud differentiation of passion fruit (take Zhuangmi 05 for example)

Vm:腋生营养分生组织;Avs:腋芽。带括号的数值表示茎尖以下的节位顺序
Vm: Vegetative meristem; Avs: Axillary vegetative shoot. The numerical values in parentheses indicate the order of nodes below the stem tip
图4 ‘壮蜜05’(A)和‘台农1号’(B)茎尖扫描电镜观察
Fig. 4 Scanning electron microscopy observation of stem tips of Zhuangmi 05 (A) and Tainong No. 1 (B)

表1 西番莲茎尖下不同节位分化进程的描述
2.2 西番莲花芽分化形态变化及花生长模型拟合
2.2.1 西番莲花芽分化进程时间线 MB初始标记时期为第一苞片形成期(图5-A1),标记后2 d额外苞片形成(图5-B1),标记后4 d萼片原基形成(图5-C1),标记后6 d萼片发育或花瓣原基形成(图5-D1),标记后8—10 d雄蕊原基形成(图5-E1),标记后12 d雌蕊形成(图5-F1、G1),标记后16 d花药、柱头形成(图5-H1);TN初始节位为第一苞片形成期(图5-A2),少数为额外苞片形成期,标记后2 d为额外苞片形成期或萼片原基形成期(图5-B2),标记后4 d萼片发育(图5-C2),标记后6 d花瓣原基形成(图5-D2),标记后8 d雄蕊原基形成(图5-e2),标记后10—12 d雌蕊形成(图5-f2、g2),标记后16 d柱头和花柱形成(图5-h2)。因此,西番莲由第一苞片形成期到雌蕊形成期所需时间为10—12 d,TN较MB花芽分化进程快1—2 d。
2.2.2 西番莲花生长曲线分析 图6可以直观看出花生长量和花形态的变化。MB和TN的第一苞片长度及宽度、花梗长、花蕾长、花蕾和花梗总长随标记后天数呈“S”形单向递增趋势。两品种第一苞片长度和宽度在标记后28 d及之后趋于稳定;花梗长从标记后8 d开始伸长,标记后24 d开始迅速伸长;花蕾长、花蕾和花梗总长前期增长较慢,从标记后24 d开始迅速增长。品种间比较可知,标记后28 d及之后,品种间的第一苞片宽度、花梗长、花蕾和花梗总长呈现出显著差异,均表现为TN明显高于MB(图7-B、C、E),而第一苞片长度和花蕾长差异不明显(图7-A、D)。

0 d放大倍数200×,比例尺100 µm;2—6 d放大倍数100×,比例尺200 µm;8—16 d放大倍数40×,比例尺500 µm
0 d magnification 200×, scale 100 µm; 2-6 d magnification 100×, scale 200 µm; 8-16 d magnification of 40×, scale 500 µm
图5 ‘壮蜜05’和‘台农1号’于标记后0—16 d的花芽纵切观察
Fig. 5 Longitudinal observation of flower buds of Zhuangmi 05 and Tainong No. 1 at 0-16 days after marking

图6 ‘壮蜜05’和‘台农1号’标记后4—40 d的花形态
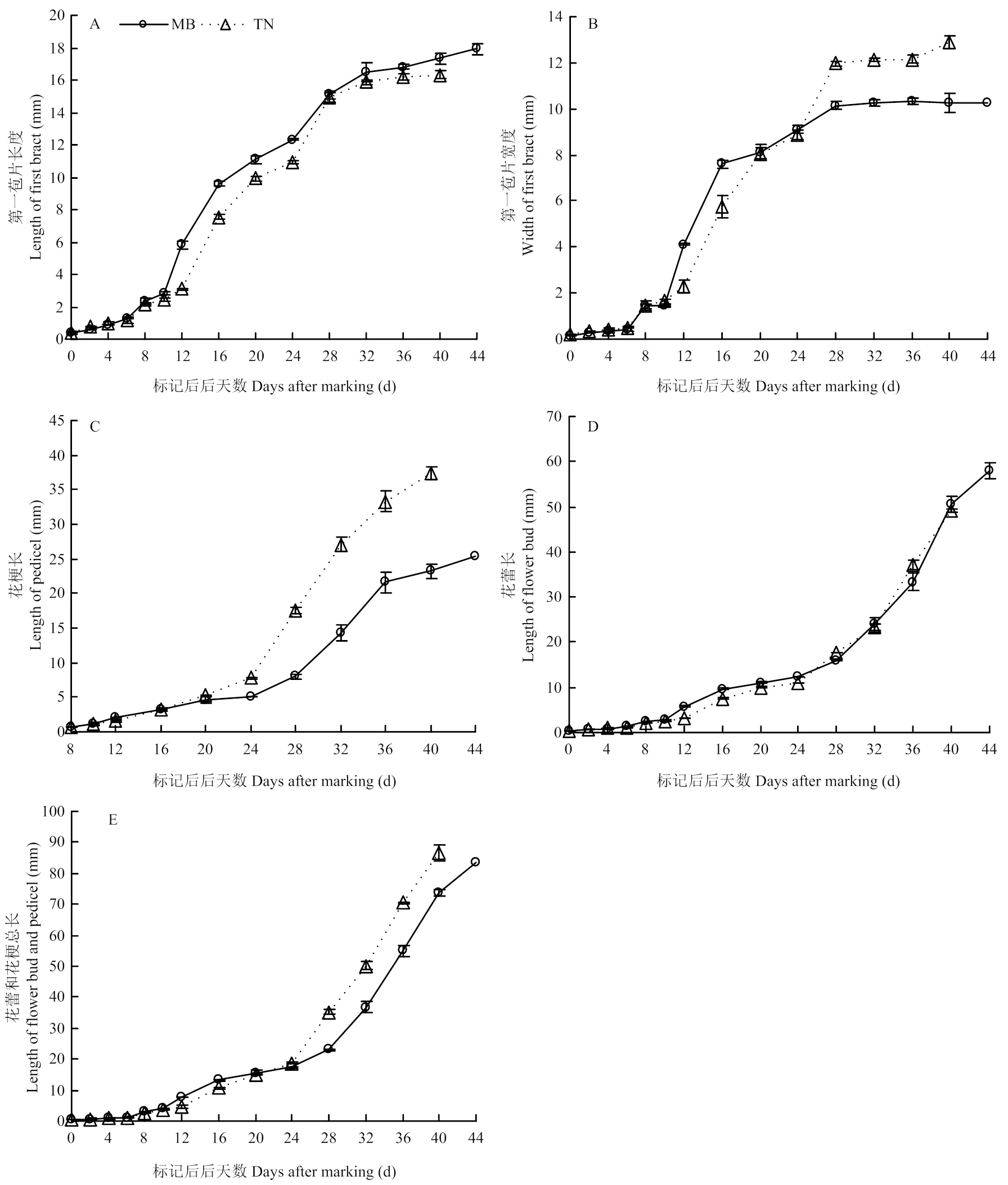
图7 ‘壮蜜05’和‘台农1号’的花生长曲线
Fig. 7 Flower growth curves of Zhuangmi 05 and Tainong No. 1
2.2.3 花生长模型拟合及模型验证 利用Origin软件对MB和TN的第一苞片长度、第一苞片宽度、花梗长、花蕾长、花蕾和花梗总长共5个花形态指标和标记后天数进行Logistic模型的非线性回归拟合。由表2可以看出,各花器官生长模型的决定系数2在0.9524—0.9988,且各方程均达到极显著水平,说明模型方程的拟合效果较好。将试验中MB和TN的花生长指标实测值分别代入生长模型进行验证,由表2可知,各处理间实测值和模拟值的标准化均方根误差()为8.54%—19.62%,其中,TN的第一苞片长度、花蕾和花梗总长均低于10%,模型表现极好,其他生长模型10%<<20%,表现良好。说明本试验条件下建立的西番莲花生长Logistic模型可以较好地拟合花器官实际生长动态。
2.2.4 花生长模型特征参数分析 由表3可知,MB和TN的第一苞片长度分别在标记后23.64 d和26.36 d进入生长缓增期,之后萼片长度超过第一苞片长度,第一苞片宽度生长趋势与第一苞片长度基本一致。其次,MB和TN花梗长的快增期分别为标记后28.44—50.46 d和标记后26.90—43.28 d,分别在标记后39.45 d和35.09 d达到最大生长速率,TN的花梗最大生长速率较MB高87.27%,与其花梗长较长有关。MB花蕾长在标记后的29.32 d之前生长较慢,这之后进入生长快增期,快增期平均生长速率为0.0037 mm∙d-1,在标记后39.43 d时花蕾长达到最大生长速率,为2.926 mm∙d-1,按照实际观察,MB在标记后39—44 d花蕾长达到最大值后开花;TN呈现类似趋势,在标记后37.09 d达到最大生长速率,实际观察其在标记后36—41 d花蕾长达最大值后开花。MB和TN的花蕾和花梗总长则分别在标记后24.30 d和25.11 d之后进入快速增长期,直至开花一直处于快速生长状态。

表2 西番莲花器官Logistic生长模型的拟合
V1为最大生长速率,t1为最大生长速率对应的标记后天数,t2、t3为生长曲线上的拐点,V2为快增期的平均生长速率。0—t2为生长渐增期、t2—t3为生长快增期、t3至开花为生长缓增期
V1is the maximum growth rate, t1is the number of days after marking corresponding to the maximum growth rate, t2and t3are the inflection points on the growth curve, and V2is the average growth rate during the rapid growth period. 0-t2represents the gradual growth period, t2-t3represents the rapid growth period, and t3to flowering represents the slow growth period
3 讨论
3.1 西番莲花芽分化的过程及特点
当西番莲植株生长达到一定节位以上时,花原体在外界环境适合时迅速发育形成具有花器的花芽[6]。西番莲茎顶端分生组织先后经历叶片和托叶的形成、卷须和花芽的形成,西番莲的花芽分化过程主要包括:第一苞片形成、额外的苞片形成、萼片形成、花瓣形成、雄蕊形成、雌蕊形成、副花冠形成,之后雌蕊分化出子房和胚珠。本研究中西番莲从第一苞片形成到雌蕊形成历时10—12 d,从第一苞片形成到开花历时36—44 d,其中TN花芽分化进程稍快于MB,但远低于前人报道的紫果西番莲从花芽分化至开花需60 d[8],这可能与品种差异和气候差异有关;若以雌蕊形成作为花芽形成的标志,则本研究中花芽形成到开花所需时间为26—32 d,与张育森等[6]报道的TN从具有花器的花芽到开花约一个月接近。本研究中西番莲花芽分化观察取材时间为8月,据观察,不同季节花芽分化进程存在差异,但波动幅度不大,将另文报道。从西番莲花芽分化过程中苞片、卷须、叶片的形态变化观察中发现,当叶片长度在0.7—1.0 mm时,该节位花原基形成,当叶片伸长到1.2 mm时第一苞片分化,此时肉眼可见该节位叶片,因此可设为标记的初始节位;当叶片长6.8 mm、卷须长3 mm、第一苞片长1.35—1.41 mm时,萼片分化或萼片形成;当第一苞片长度为3.23 mm(TN)和3.64 mm(MB)时雌蕊原基分化。Nave等[14]的研究也将花芽长度3 mm作为西番莲花是否发育成功的节点指标,因此,雌蕊形成是花芽分化成功的标志,从表观形态上可根据苞片长度、叶片长度等指标推测西番莲的花芽分化进程。
西番莲的花芽形成有其自身的生物学特性和规律,不同于葡萄[15-17]、荔枝[18-20]等果树的花芽由休眠后的营养芽转化发育而来,西番莲的花芽从当年新生枝条的叶腋发出,且合适条件下各级枝蔓可多批次开花[2]。由电镜图可清晰看到花芽与营养芽分开独立,在茎顶端下第9节可观察到腋生营养分生组织出现在卷须和花芽内侧,此时花芽处在额外苞片形成期,之后腋生营养分生组织发育形成腋芽。由此可见,西番莲花芽分化过程中,花芽与营养芽同步生长。本研究还观察到不同西番莲品种间营养芽发育成侧枝的数量差异很大,一部分品种侧枝很少;而一部分品种侧枝茂盛,需要及时抹除,以保证花芽的正常发育和开花。笔者前期研究发现,西番莲花芽一般从新生枝蔓基部以下第4节位及之后开始分化出现,如TN夏秋季平均花芽分化率为77.2%,且分化的花芽有一部分黄化凋落,花芽退化率高达41.4%[2],因此,西番莲花芽与营养芽发育体现的生殖生长和营养生长的竞争与平衡关系有待进一步研究。
3.2 西番莲花生长模型的拟合
基于Logistic的生长模型主要应用在作物株高[10,21-22]、果实生长[11,23-24]、叶面积指数[25-26]、干物质积累量[27-28]、耐热性[29-30]等性状的拟合和模型预测。本研究主要关注的是西番莲花形态生长(因变量)与发育时间进程(自变量)的模型拟合,并对花器官生长阶段进行划分。对两个西番莲品种的第一苞片长度、第一苞片宽度、花梗长、花蕾长、花蕾和花梗总长共5个花形态指标和标记后天数进行Logistic模型的非线性回归拟合,模型拟合效果较好[13,31],可用来预测两品种花形态指标的生长动态。本研究发现西番莲花形态生长有两个主要阶段,一是前期的苞片生长期,二是后期的花梗和花蕾生长期。如苞片的长度在标记后即第一苞片形成期后11—12 d进入快增期,第一苞片形成期后24—26 d进入缓增期;之后花梗和萼片迅速生长,萼片长度超过苞片,花蕾和花梗总长在第一苞片形成期后24—25 d进入快增期,第一苞片形成期后41—42 d进入缓增期到最大值即开花。因此,可以根据西番莲田间的花形态指标预测开花时间和判断其生长阶段处于渐增期、快增期还是缓增期。
综上,可将西番莲花芽分化和形态分化分为3个阶段,第一阶段:第一苞片形成期到雌蕊形成期,历时10—12 d,第一苞片长度从1.2 mm伸长到3—4 mm;第二阶段:苞片生长期,历时12—14 d,第一苞片长度从约3—4 mm伸长到约12 mm达最大值;第三阶段:花梗和花蕾生长期,历时15—17 d,花梗长从5—8 mm伸长到25—37 mm,花蕾长从约12 mm伸长到约50 mm,达最大值后开花。在西番莲实际生产中,可通过花生长模型预测开花时间及花芽分化进程,为促花保花方案提供参考依据。
4 结论
西番莲花芽分化过程主要包括第一苞片形成期、额外苞片形成期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期和副花冠形成期等,从第一苞片形成到开花历时36—44 d。本研究中拟合的花生长模型可较好地模拟花生长动态,生产中可通过花形态指标预测开花时间;同时结合花芽分化进程与花形态指标对应关系,根据各节位花苞片、叶片、卷须形态指标大致判断花芽分化进程。
[1] 中国科学院中国植物志编辑委员会. 中国植物志-第六卷, 第一分册-蕨类植物门. 北京: 科学出版社, 1999.
Sinicae Agendae Academiae Sinicae Edita. Flora reipublicae popularis sinicae. Beijing: Science Press, 1999. (in Chinese)
[2] 田青兰, 吴艳艳, 黄伟华, 刘洁云, 韦绍龙, 牟海飞, 韦弟, 黄永才, 熊晓兰, 张英俊. ‘台农1号’西番莲的成花坐果特性及与气象因子的关系. 果树学报, 2020, 37(9): 1358-1370.
TIAN Q L, WU Y Y, HUANG W H, LIU J Y, WEI S L, MOU H F, WEI D, HUANG Y C, XIONG X L, ZHANG Y J. Flower formation and fruit setting in ‘Tainong No.1’ passion fruit and its relationship with meteorological factors. Journal of Fruit Science, 2020, 37(9): 1358-1370. (in Chinese)
[3] 秦志聪, 朱继生, 冯钊, 陈永孝. 桂林不良气候对百香果坐果的影响及其应对措施. 南方园艺, 2017, 28(2): 28-30.
QIN Z C, ZHU J S, FENG Z, CHEN Y X. Influence of bad climate in Guilin on fruit setting of passion fruit and its countermeasures. Southern Horticulture, 2017, 28(2): 28-30. (in Chinese)
[4] 林秋金, 苏金强, 王美盛, 史国强, 武英, 李丽蓉, 牛先前. 气象因子对百香果坐果率的影响研究初报. 福建热作科技, 2019, 44(3): 18-21.
LIN Q J, SU J Q, WANG M S, SHI G Q, WU Y, LI L R, NIU X Q. A preliminery report on passion fruit fertile percentage influenced by meteorology factor. Fujian Science & Technology of Tropical Crops, 2019, 44(3): 18-21. (in Chinese)
[5] 马月萍, 戴思兰. 植物花芽分化机理研究进展. 分子植物育种, 2003, 1(4): 539-545.
MA Y P, DAI S L. Flower bud differentiation mechanism of anthophyta. Molecular Plant Breeding, 2003, 1(4): 539-545. (in Chinese)
[6] 张育森, 郑正勇. 百香果开花习性之研究. 中国园艺, 1988, 34(4): 271-282.
ZHANG Y S, ZHENG Z Y. On the flowering habits of passionfruits. Journal of Chinese Society for Horticutural Science, 1988, 34(4): 271-282.(in Chinese)
[7] 武春昊, 王强, 卢明艳, 闫兴凯, 刘明鹤, 张茂君. ‘单花’梨花芽特性及其形态分化过程研究. 园艺学报, 2019, 46(7): 1373-1378.
WU C H, WANG Q, LU M Y, YAN X K, LIU M H, ZHANG M J. Study on characteristics and floral bud development of flower bud in ‘Danhua’ pear. Acta Horticulturae Sinica, 2019, 46(7): 1373-1378. (in Chinese)
[8] ISHIHATA K, SHINDO T, IWAHORI S. Flower-bud differentiation and development in purple passion fruit,Sims. Bulletin of the Faculty of Agriculture, Kagoshima University, 1989(39): 103-119.
[9] 徐苏. 作物生长模型的研究进展. 安徽农学通报, 2023, 29(4): 26-32.
XU S. Research progress of crop growth model. Anhui Agricultural Science Bulletin, 2023, 29(4): 26-32. (in Chinese)
[10] 陈杨, 王磊, 白由路, 卢艳丽, 倪露, 王玉红, 徐孟泽. 有效积温与不同氮磷钾处理夏玉米株高和叶面积指数定量化关系. 中国农业科学, 2021, 54(22): 4761-4777. doi: 10.3864/j.issn.0578-1752.2021. 22.005.
CHEN Y, WANG L, BAI Y L, LU Y L, NI L, WANG Y H, XU M Z. Quantitative relationship between effective accumulated temperature and plant height & leaf area index of summer maize under different nitrogen, phosphorus and potassium levels. Scientia Agricultura Sinica, 2021, 54(22): 4761-4777. doi: 10.3864/j.issn.0578-1752.2021.22.005. (in Chinese)
[11] 马二磊, 黄芸萍, 臧全宇, 丁伟红, 王毓洪. 甜瓜植株和果实生长模型的拟合与分析. 南方农业学报, 2018, 49(7): 1358-1363.
MA E L, HUANG Y P, ZANG Q Y, DING W H, WANG Y H. Growth model fitting and analysis of melon plant and fruit. Journal of Southern Agriculture, 2018, 49(7): 1358-1363. (in Chinese)
[12] 石国朝. 冬枣花生长发育模型研究[D]. 杨凌: 西北农林科技大学, 2022.
SHI G Z. Development and growth model of flowers in‘Dongzao’ [D]. Yangling: Northwest A & F University, 2022. (in Chinese)
[13] BANNAYAN M, HOOGENBOOM G. Using pattern recognition for estimating cultivar coefficients of a crop simulation model. Field Crops Research, 2009, 111(3): 290-302.
[14] NAVE N, KATZ E, CHAYUT N, GAZIT S, SAMACH A. Flower development in the passion fruitrequires a photoperiod-induced systemic graft-transmissible signal. Plant, Cell & Environment, 2010, 33(12): 2065-2083.
[15] 罗惠格, 朱维, 黄泳碧, 陈潇, 林玲, 白扬, 曹雄军, 白先进, 张唯, 王博. 阳光玫瑰葡萄生长期花芽分化形态进程及相关生理分子水平变化研究. 果树学报, 2023, 40(1): 74-87.
LUO H G, ZHU W, HUANG Y B, CHEN X, LIN L, BAI Y, CAO X J, BAI X J, ZHANG W, WANG B. A study on the morphological process and physiological and molecular changes of flower bud differentiation in Shine Muscat grape during fruit growing season. Journal of Fruit Science, 2023, 40(1): 74-87. (in Chinese)
[16] 王博, 罗惠格, 覃富强, 陈祥飞, 朱维, 谢太理, 曹雄军, 白先进. 葡萄花芽分化研究进展. 南方农业学报, 2023, 54(3): 957-968.
WANG B, LUO H G, QIN F Q, CHEN X F, ZHU W, XIE T L, CAO X J, BAI X J. Research progress of grape flower bud differentiation. Journal of Southern Agriculture, 2023, 54(3): 957-968. (in Chinese)
[17] 贾楠. 葡萄花芽分化及其主要影响因素的研究进展. 河北果树, 2020(1): 1-3.
JIA N. Review on grape floral bud differentiation and its major influencing factors. Hebei Fruits, 2020(1): 1-3. (in Chinese)
[18] 陈厚彬, 苏钻贤, 张荣, 张红娜, 丁峰, 周碧燕. 荔枝花芽分化研究进展. 中国农业科学, 2014, 47(9): 1774-1783.doi: 10.3864/j.issn. 0578-1752.2014.09.012.
CHEN H B, SU Z X, ZHANG R, ZHANG H N, DING F, ZHOU B Y. Progresses in research offloral differentiation. Scientia Agricultura Sinica, 2014, 47(9): 1774-1783. doi: 10.3864/j.issn.0578- 1752.2014.09.012. (in Chinese)
[19] 陈厚彬, 欧良喜, 李建国, 苏钻贤, 杨胜男, 吴振先, 胡卓炎. 新中国果树科学研究70年: 荔枝. 果树学报, 2019, 36(10): 1399-1413.
CHEN H B, OU L X, LI J G, SU Z X, YANG S N, WU Z X, HU Z Y. Fruit scientific research in New China in the past 70 years:. Journal of Fruit Science, 2019, 36(10): 1399-1413. (in Chinese)
[20] 黄辉白, 陈厚彬. 以阶段观剖视荔枝的花芽分化. 果树学报, 2003, 20(6): 487-492.
HUANG H B, CHEN H B. A phasic approach towards the floral formation insonn. Journal of Fruit Science, 2003, 20(6): 487-492. (in Chinese)
[21] 肖哲元, 雷宏军, 张振华, 张倩, 金翠翠, 孙克平. 基于Logistic模型的加气灌溉辣椒生长特性和产量研究. 灌溉排水学报, 2022, 41(7): 16-23.
XIAO Z Y, LEI H J, ZHANG Z H, ZHANG Q, JIN C C, SUN K P. Modelling growth and yield of aerated pepper by the Logistic model. Journal of Irrigation and Drainage, 2022, 41(7): 16-23. (in Chinese)
[22] 雷涛, 郭向红, 毕远杰, 吕棚棚, 马娟娟, 孙西欢, 张勇, 雷震. 基于Logistic模型的番茄生长特性研究. 节水灌溉, 2020(10): 10-14.
LEI T, GUO X H, BI Y J, LÜ P P, MA J J, SUN X H, ZHANG Y, LEI Z. Study on tomato growth characteristics based on logistic model. Water Saving Irrigation, 2020(10): 10-14. (in Chinese)
[23] 朱海军, 生静雅, 刘广勤, 陈亚辉, 曹福亮. 基于Logistic模型的薄壳山核桃果实生长发育研究. 西南农业学报, 2015, 28(3): 1231-1235.
ZHU H J, SHENG J Y, LIU G Q, CHEN Y H, CAO F L. Study on growth and development of pecan fruit based on logistic model. Southwest China Journal of Agricultural Sciences, 2015, 28(3): 1231-1235. (in Chinese)
[24] 徐臣善, 徐爱红, 高东升, 程述汉. 苹果果实生长的数学模型及各生长指标间的相关性分析. 植物科学学报, 2015, 33(1): 72-80.
XU C S, XU A H, GAO D S, CHENG S H. Mathematical model of apple fruit growth and correlation analysis among growth indices. Plant Science Journal, 2015, 33(1): 72-80. (in Chinese)
[25] 王全九, 刘云鹤, 苏李君. 基于单参数Logistic的典型作物相对叶面积指数模型研究. 农业机械学报, 2020, 51(7): 210-219.
WANG Q J, LIU Y H, SU L J. Relative leaf area index of typical crops based on single parameter logistic model. Transactions of the Chinese Society for Agricultural Machinery, 2020, 51(7): 210-219. (in Chinese)
[26] 赵坤, 柳平增, 张泽, 张艳, 马峰. 基于Logistic模型的设施番茄生长过程数字化研究. 中国农机化学报, 2023, 44(9): 72-78.
ZHAO K, LIU P Z, ZHANG Z, ZHANG Y, MA F. Digital research on growth process of facility tomato based on Logistic model. Journal of Chinese Agricultural Mechanization, 2023, 44(9): 72-78. (in Chinese)
[27] 张继红, 刘云鹤, 王全九, 苏李君, 郭毅, 王康. 典型作物Logistic模型生长参数空间分布及其地区水热相关性. 干旱地区农业研究, 2021, 39(5): 199-209.
ZHANG J H, LIU Y H, WANG Q J, SU L J, GUO Y, WANG K. Typical crops’ spatial variability of Logistic model parameters and its correlation with regional water and heat. Agricultural Research in the Arid Areas, 2021, 39(5): 199-209. (in Chinese)
[28] 刘振洋, 吴鑫雨, 汤利, 郑毅, 李海叶, 潘浩男, 朱东宇, 王静静, 黄少欣, 覃潇敏, 肖靖秀. 小麦蚕豆间作体系氮素吸收累积动态及其种间氮素竞争关系. 植物营养与肥料学报, 2020, 26(7): 1284-1294.
LIU Z Y, WU X Y, TANG L, ZHENG Y, LI H Y, PAN H N, ZHU D Y, WANG J J, HUANG S X, QIN X M, XIAO J X. Dynamics of N acquisition and accumulation and its interspecific N competition in a wheat-faba bean intercropping system. Journal of Plant Nutrition and Fertilizers, 2020, 26(7): 1284-1294. (in Chinese)
[29] 凌瑞, 董钠, 刘惠斌, 郑泽新, 翟俊文, 吴沙沙. 应用Logistic方程测定八个绣球品种的耐热性. 北方园艺, 2021(11): 63-69.
LING R, DONG N, LIU H B, ZHENG Z X, ZHAI J W, WU S S. Heat resistance of eight cultivars oflogistic equation. Northern Horticulture, 2021(11): 63-69. (in Chinese)
[30] 李进, 顾绘, 殷琳毅. 6个草莓品种高温半致死温度与耐热性评价. 中国果树, 2021(1): 56-58.
LI J, GU H, YIN L Y. Evaluation of heat tolerance and high semi-lethal temperature of six strawberry cultivars. China Fruits, 2021(1): 56-58. (in Chinese)
[31] 程陈, 冯利平, 薛庆禹, 李春, 宫志宏, 董朝阳, 伍露, 王春雷, 刘淑梅, 李奕卓, 黎贞发. 日光温室黄瓜生长发育模拟模型. 应用生态学报, 2019, 30(10): 3491-3500.
CHENG C, FENG L P, XUE Q Y, LI C, GONG Z H, DONG C Y, WU L, WANG C L, LIU S M, LI Y Z, LI Z F. Simulation model for cucumber growth and development in sunlight greenhouse. Chinese Journal of Applied Ecology, 2019, 30(10): 3491-3500. (in Chinese)
Observation of Flower Bud Differentiation Process and Fitting of Flower Growth Model of Passion Fruit
Biotechnology Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530007
【Objective】This study aimed to elucidate the flower bud differentiation process in the primary passion fruit () varieties within Southern China’s ecological region. Additionally, it sought to establish a flower growth model depicting the morphological differentiation of passion fruit flower buds, so as to serve as a reference for promoting and retenting passion fruit flowers.【Method】Tainong No. 1 (TN) as the predominant cultivar of the purple fruit variety, and Zhuangmi 05 (also known as Zhuangxiangmibao, MB) as the main cultivar of the yellow fruit variety in Guangxi, China, were utilized as experimental materials. The scanning electron microscopy and paraffin section dissection were employed to observe flower bud differentiation process in these varieties. The differentiation process of nodes below the shoot apical meristem, and measured the first bract length of flower buds, leaf length, and tendril length at each node were observed too. The 7th node below the shoot apical meristem was marked as the initial marker node, and this marked the onset of the first bract differentiation phase of the flower bud. The flower bud differentiation from 0 to 16 days after marking were investigated, as well as the dynamic changes in five flower morphological indicators, such as the length of the first bract and bud from marking to flowering were tracked, for facilitating the construction of a flower growth model.【Result】The primary process of flower bud differentiation in the tested passion fruit varieties encompassed first bract formation stage, additional bract formation stage, sepal formation stage, petal formation stage, stamen formation stage, pistil formation stage, corona formation stage, etc. The duration from first bract formation to pistil formation spanned 10-12 days, with TN exhibiting 1-2 day advancement over MB. Pistil formation emerged as a vital marker for successful passion fruit flower bud development, characterized by a 3-4 mm first bract length. The duration from first bract formation to flowering ranged between 36-44 days, with TN progressing 3-4 days earlier than MB. Tendril differentiation was visible at the 4th-5th nodes below the shoot apical meristem, flower primordium differentiation at the 6th node, appearing alongside the tendril, first bract differentiation at the 7th node, vegetative meristem appearing at the 9th node, which were positioned separately on the inner side of the tendril and flower bud, axillary vegetative shoot formation at the 10th-11th nodes, and pistil primordium differentiation at the 14th-15th nodes. The non-linear regression fitting of a Logistic model in Origin software was performed for the first bract length, width, pedicle length, flower bud length, length of flower bud and pedicle along with days after making for both MB and TN, and the results showed the determination coefficient2ranged from 0.9524 to 0.9988, and the normalized root mean square error () was between 8.54% to 19.62%, indicating a good fit of the model equation. Based on the model parameters and actual observations, the length of bract entered a rapid growth period 11-12 days after marking (i.e. after the first bract formation stage) and stabilized 24-26 days after marking. Subsequently, the pedicels and sepals grew rapidly, with the sepals surpassing the bracts length, and the total length of the flower bud and pedicel entered a rapid growth phase 24-25 days after marking, blooming after reaching maximum growth at 41-42 days after marking.【Conclusion】The flower buds of passion fruit were closely associated with tendrils, and independent of nutritional buds. The flower bud differentiation and morphological differentiation of passion fruit could be divided into three stages: the first bract formation period to the pistil formation period, lasting 10-12 days; the bract growth period, lasting 12-14 days; and the flower buds and pedicels growth period, lasting 15-17 days. In actual production of passion fruit, the flowering time could be predicted using the flower growth model, and the flower bud differentiation process could be assessed using morphological indicators, such as bract length, which provided a reference basis for promoting flowering and promoting flowers.
passion fruit; flower bud differentiation; flower growth model; morphological differentiation

2023-07-07;
2023-11-30
广西自然科学基金(2021GXNSFBA075044)、广西科技重大专项(桂科AA22068091-1)、国家自然科学基金(32260740,32060660)、广西自然科学基金(2023GXNSFAA026301)、广西农业科学院基本科研业务专项项目(桂农科2021YT089)、广西浦北百香果试验站项目(桂TS2022010)
田青兰,E-mail:tianqinglan1991@163.com。通信作者牟海飞,E-mail:mhf@gxaas.net
(责任编辑 赵伶俐)
