禁牧对西藏亏祖山温性草原主要植物生态位及种间联结的影响
2024-03-08杨金涛张晓晗尹正辉桑旦群培古桑群宗
杨金涛,张晓晗,尹正辉,桑旦群培,古桑群宗*
(1 西藏大学 生态环境学院,拉萨 850000;2 拉萨市第一中等职业技术学校,拉萨 850000)
优势种是对群落结构的形成有明显控制作用的物种,是研究生态过程的关键[1]。对优势种生态位的分析,能够了解植物群落的结构特征及植物在其生境中的空间分布格局,并对群落稳定性做出解释[2]。关于植物群落内种间关系的研究主要集中在种间联结和种间相关两方面[3],种间联结和种间关联相结合,能够较客观地解释研究区域植物群落内物种间的关系[4-5],也有助于进一步研究禁牧条件下的种群分布格局。草原作为青藏高原生态系统重要的组成部分,生态功能价值巨大,其在生物多样性保护方面发挥关键作用[6]。放牧不仅能够改变植物群落的数量结构特征,还能改变主要植物种之间的相互关系[7]。过度放牧会导致植物群落多样性下降,草地生产力降低,进而会造成草地面积退化[8-9]。相关数据表明放牧活动对草地生态系统造成巨大影响,尤其是对于高海拔区域,其变化的程度要高于全球平均水平[10],而禁牧是恢复退化草地的主要途径,能改善草地植物群落结构,降低杂类草比例,且在草地生态系统健康维持中起重要作用[11]。拉萨河流域林周县白朗村亏祖山温性草原位于西藏自治区中南部,属于较为脆弱的高寒草地区[12],山体海拔跨度大,植被类型多样,是研究禁牧对该区域主要植物生态位和种间联结影响的优良试验场所。目前,关于亏祖山温性草原的研究主要集中在种质资源调查、植物物种丰富度及群落结构特征研究和小型土壤动物多样性研究等方面[12-13],而有关禁牧如何改变植物生态位和种间关系的研究则较少。本文从生态位和种间联结的角度阐释了亏祖山温性草原8个主要植物物种对禁牧的响应,以期为制定科学的管理措施提供依据。
1 材料和方法
1.1 研究区概况
研究区域选于西藏拉萨林周县卡孜乡亏组山(91°07′33″—91°07′19″E,29°52′23″—29°49′50″N),属于拉萨河支流澎波藏曲流域西南部,距林周县10 km 左右,年日照时间大于3 000 h,年均气温7.5~7.6 ℃,年均降水量在440 mm 左右,主要集中在6—9月[14]。研究区域为亏祖山温性草原地段,主要植物包括草沙蚕(Tripogonbromoides)、白草(Pennisetumflaccidum)、高山嵩草(Kobresiapygmaea)、丝颖针茅(Stipacapillacea)、矮生嵩草(Kobresiahumilis)、笔直黄耆(Astragalusstrictus)、藏橐吾(Ligulariarumicifolia)、肉果草(Lanceatibetica)等。
1.2 研究方法
1.2.1 试验设计
试验地位于该山体温性草原区域内,于2012年在山体海拔3 900 m 和4 000 m 各设置1个禁牧处理样地,样地面积为20 m×20 m,禁牧外为自然放牧区域。本研究组于2019年8月(禁牧第7年)采用典型取样法在2个样地的禁牧处理和自然放牧区域各设置5个1 m×1 m 草本样方,共计20个样方开展植被调查。记录的植物物种指标包括:植物种名、植物生长高度(3个重复)、盖度、多度、地上生物量等。在本试验中,禁牧处理用CK 表示,自然放牧处理用GRZ表示。
1.2.2 物种选取
根据2019年8月植被调查结果,选取自然放牧区域和禁牧区域2个群落中均出现的8个主要植物种作为研究对象。亏祖山温性草原8 个主要植物种:草沙蚕(S1)、高山嵩草(S2)、白草(S3)、丝颖针茅(S4)、矮生嵩草(S5)、笔直黄芪(S6)、藏橐吾(S7)、肉果草(S8)。
1.2.3 指标计算方法
生态位宽度(Bi)和生态位重叠值(NO)用Levins方法[4]计算:
式中:r为资源状态数;j为资源;Pij为物种i利用资源j的个体占该种所有个体数的比例;Pkj为物种k利用资源j的个体数占所有个体数的比例。
采用方差比率法(variance ratio,Vr)对群落的总体联结性进行[15]计算:
式中:Pi为物种i的频度;N为总样方数;S为总物种数,当Vr>1表示种间为正的关联,反之为负;W为检验Vr值显著程度的统计量。
种对间联结性——χ2检验统计量[16]。该方法基于2×2列联表检验种对之间的联结性和显著程度。由于取样的非连续性会造成偏差,可用Yates连续校正系数纠正[4],公式为:
式中:当χ2<3.814时,说明种对之间联结性不显著(P>0.05);当3.814<χ2<6.635时,说明种对之间联结性显著(0.01<P<0.05);当χ2>6.635时,说明种对之间联结性极显著(P<0.01)。
种对种间联结性——关联程度。在χ2检验的基础上若种对间存在显著联结性,可用种间联结系数(Ac)测算[4]种间联结程度的大小。
式中:Ac的取值区间为[-1,1],当Ac值越接近1时,表示正联结性越强,Ac值越接近-1时,表明负联结性越强,当Ac值为0时,则表明物种间相互独立[16]。
此外,也可用Ochiai(Oi)表示物种间的联结程度和相伴出现的几率[17]:
式中:Oi的取值区间在[0,1],当值越接近1时,表明种对之间有较大的相伴出现机率;当Oi值为0时,表明种间无联结。
以上数据分析运用R4.3.1 的Spaa程序包进行计算。
2 结果与分析
2.1 主要植物生态位在不同处理下的响应
2.1.1 生态位宽度(Bi)
在禁牧群落和自然放牧群落中,8个优势物种的生态位宽度差异明显(表1)。自然放牧处理下的矮生嵩草(生态位宽为6.9)和禁牧处理下的草沙蚕(生态位宽为7.9)具有最大的生态位宽度,表明它们的资源利用能力极强。而2种处理下生态位宽度最小的物种为肉果草(生态位宽为2.1)和藏橐吾(生态位宽为1.0),表明它们的分布范围较窄。
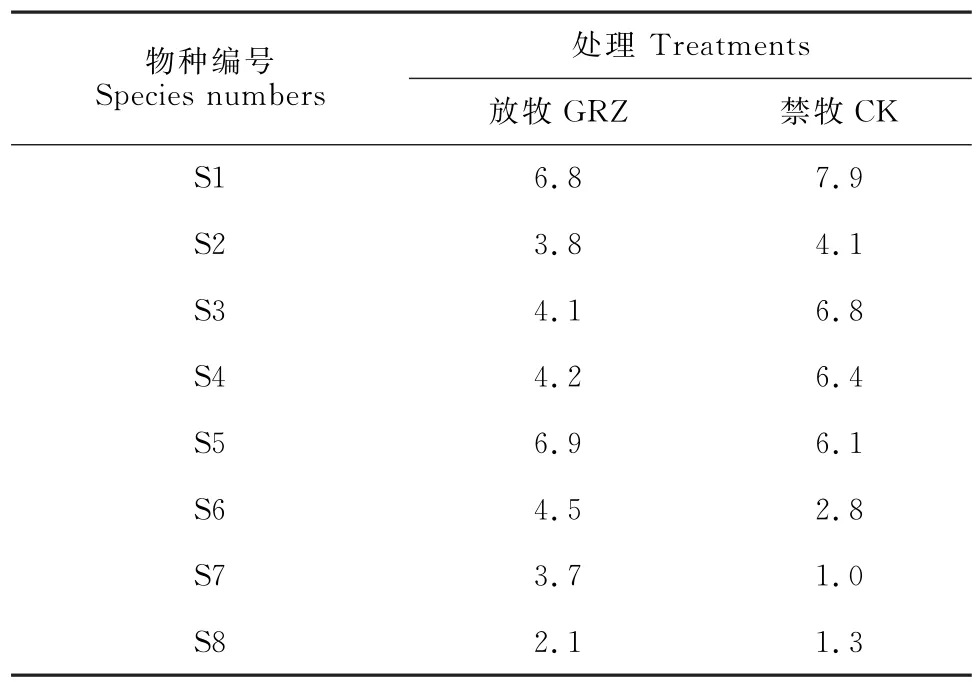
表1 亏祖山地优势物种在不同处理下生态位宽度Table 1 Niche width of the dominant species in the Kuizu Mountains under different treatments
除此之外,禁牧处理下的草沙蚕、高山嵩草、白草和丝颖针茅的生态位宽度却高于自然放牧处理,而该条件下的矮生嵩草、笔直黄芪、藏橐吾和肉果草的生态位宽度却更低,表明禁牧改变了群落中各物种的生态位宽度。
2.1.2 生态位重叠(No)
在禁牧群落和自然放牧群落中,28个优势植物种对的NO值(生态位重叠值)在0~0.83之间(图1)。自然放牧处理下,白草和肉果草的NO值最大(0.77),高山嵩草和肉果草的NO值最小(0.04)。所有种对中,NO值在0.60以上的种对有5个,NO值介于0.20~0.60之间的种对有17个,NO值在0.20以下的种对有6个。
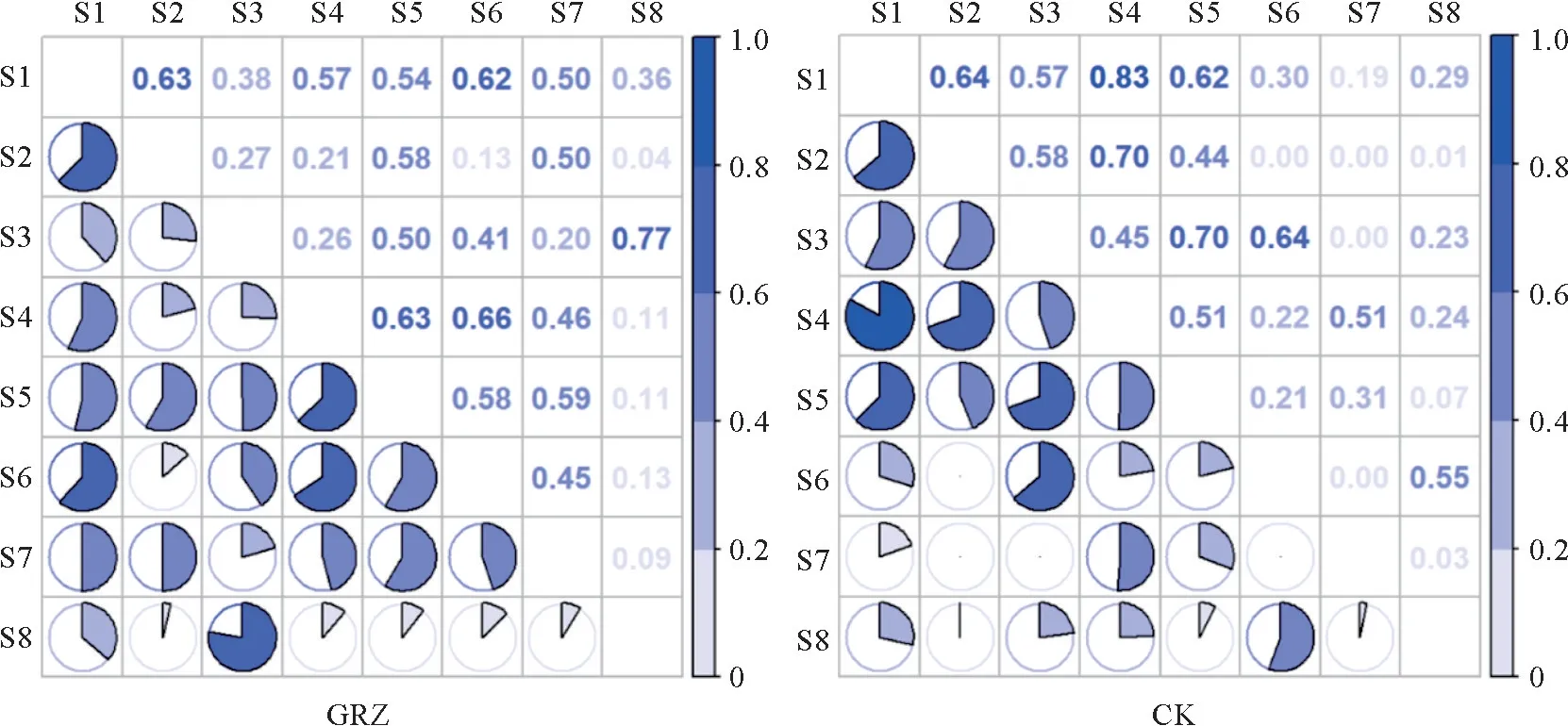
图1 亏祖山地优势物种在不同处理下的生态位重叠Fig.1 Niche overlap of the dominant species in the Kuizu Mountains under different treatments
禁牧处理下,草沙蚕和丝颖针茅的NO值最大(0.83),高山嵩草和笔直黄芪、高山嵩草和藏橐吾、白草和藏橐吾及笔直黄芪和藏橐吾的NO值最小(0)。所有种对中,NO值在0.60 以上的种对有6个,NO值介于0.20~0.60 之间的种对有14 个,NO值在0.20以下的种对有8个。
相比自然放牧,禁牧处理下的部分物种对NO值出现一定幅度的增加,其中高山嵩草和草沙蚕、草沙蚕和白草、草沙蚕和丝颖针茅及高山嵩草和丝颖针茅等种对之间的NO值升高,且高山嵩草和丝颖针茅之间的NO值增加量最大(增加量为0.49)。NO值降低最多的是白草和肉果草(减小量为0.54)。
2.2 主要植物种间联结性在不同处理下的响应
2.2.1 总体联结性
在禁牧群落和自然放牧群落中种间总体联结性Vr表明:自然放牧处理Vr(1.21)大于1,8个优势物种总体关联性呈正关联,表明自然放牧处理下各优势物种之间逐渐趋于稳定共存状态;禁牧处理下Vr(0.43)小于1,总体联结性表现出负关联,表明围封群落中各主要物种之间对环境资源的竞争程度较强。
2.2.2 χ2检验统计量
自然放牧和禁牧处理下,28个优势植物种对中正联结种对分别有13个(46.4%)和9个(32.1%);负联结种对分别有15个(53.6%)和19个(占67.9%)。由显著性可知(图2),自然放牧处理下联结性不显著的种对有18个(64.3%),如草沙蚕和高山嵩草、高山嵩草和笔直黄芪、白草和丝颖针茅及丝颖针茅和矮生嵩草之间相互独立;禁牧处理下联结性不显著的种对有22个(78.6%),如草沙蚕和矮生嵩草、高山嵩草和白草之间相互独立。
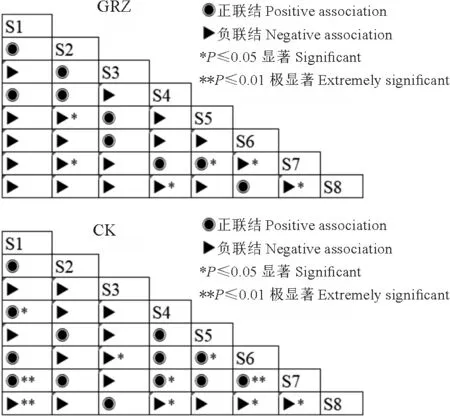
图2 亏祖山地优势物种在不同处理下的χ2检验半矩阵图Fig.2 Thesemi-matrixdiagramof χ2 testofthe dominant species in the Kuizu Mountains under different treatments
2.2.3 Ac 指数
自然放牧处理下,Ac值在0.5以上的种对有5个,Ac值介于0~0.5之间的种对有11个,Ac值介于(-0.5)~0之间的种对有6个,Ac值在-0.5以下的种对有6个(图3)。其中呈正关联与负关联的种对均占总数的42.9%。进一步分析可知,高山嵩草和丝颖针茅、白草和丝颖针茅、高山嵩草和矮生嵩草之间呈正关联,而草沙蚕和高山嵩草、草沙蚕和丝颖针茅、草沙蚕和白草、草沙蚕和矮生嵩草与高山嵩草和白草之间呈负关联,丝颖针茅和矮生嵩草、丝颖针茅和藏橐吾、矮生嵩草和笔直黄芪及笔直黄芪和藏橐吾之间无联结性。说明自然放牧条件下,草沙蚕和高山嵩草、草沙蚕和丝颖针茅、草沙蚕和白草、草沙蚕和矮生嵩草及高山嵩草和白草等物种之间存在较为激烈的资源竞争。
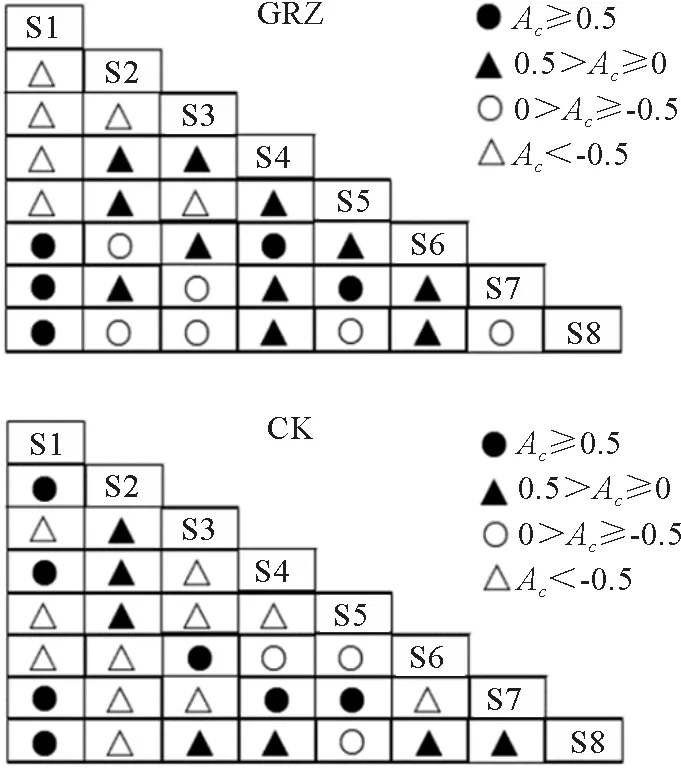
图3 亏祖山地优势物种在不同处理下的联结系数半矩阵图Fig.3 The semi-matrix diagram of association coefficient of the dominant species in the Kuizu Mountains under different treatments
禁牧处理下,Ac值在0.5以上的种对有7个,Ac值介于0~0.5之间的种对有7个,Ac值介于(-0.5)~0之间的种对有3个,Ac值在-0.5以下的种对有11个(图3)。其中呈负关联的种对占总数的50%。进一步分析可知,草沙蚕和高山嵩草、草沙蚕和丝颖针茅、高山嵩草和丝颖针茅及丝颖针茅和藏橐吾呈正关联,而草沙蚕和白草、草沙蚕和矮生嵩草、白草和丝颖针茅、丝颖针茅和矮生嵩草、矮生嵩草和笔直黄芪及笔直黄芪和藏橐吾之间则呈负关联,高山嵩草和白草、高山嵩草和矮生嵩草之间不存在联结性。禁牧改变了草沙蚕和高山嵩草、草沙蚕和丝颖针茅之间的关联性(由自然放牧中的负关联变为禁牧中的正关联),也改变了白草和丝颖针茅之间的关联性(由自然放牧中的正关联变为禁牧中的负关联),表明禁牧封育改变了部分物种之间的种间关系。
2.2.4 Oi 指数
高寒草地植物优势物种间的Oi关联指数分析结果(图4)表明,在禁牧和自然放牧处理下,Oi≥0.8 的种对数分别是4 个(占14.3%)、5 个(占17.9%);0.5≤Oi<0.8的种对数分别为12个(占42.9%)、16个(占57.1%);0.2≤Oi<0.5的种对数分别为7个(25%)、7个(25%);Oi<0.2的种对数分别为5个(占17.9%)、0个,说明上述条件下大多数物种联结程度均比较高,而禁牧相较于自然放牧处理,其优势物种种对间的关联程度增加。
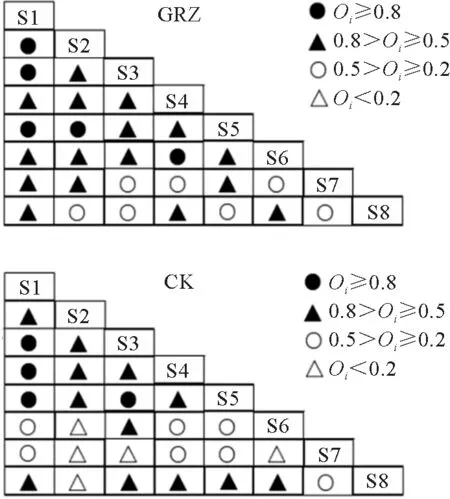
图4 亏祖山地优势物种在不同处理下的Oi 指数半矩阵图Fig.4 The semi-matrix diagram of Ochiai indices of the dominant species in the Kuizu Mountains under different treatments
3 讨论
禁牧会改变温性草原植物物种的生态位宽度[17]。本研究中,禁牧处理下的草沙蚕与自然放牧处理下的矮生嵩草生态位宽度最大(表1),表明这些物种在群落中占优势,且对于维持群落内部稳定发挥着重要作用[19]。而禁牧处理下的藏橐吾和自然放牧处理下的肉果草生态位宽度最小,说明这些物种分布范围相对较小[19]。在自然放牧处理下,白草的生态位宽度降低程度最大,而藏橐吾的生态位宽度增加程度最大,在自然放牧处理下草沙蚕的优势地位被矮生嵩草所取代。这种变化情况可能与物种的生态习性相关。白草作为高寒草地的优势种,同时也是牲畜最喜食的牧草之一,放牧会对草地植物群落产生抑制作用,即通过采食、践踏等行为抑制植物的生长,还可能导致表层土壤养分胁迫,进一步减少地上生物量[20],所以放牧活动会降低白草生态位宽度。同时,禁牧会使笔直黄芪等退化指示植物的生态位宽度降低,而使草沙蚕、丝颖针茅等适口性禾草类植物的生态位宽度增加,这是由于封育后,因家畜采食和践踏等干扰程度降低,植株高大的禾草类植物因根系较深具有较强的资源竞争优势[11],根据梯度胁迫假说,外界压力大则植物种间竞争较弱,反之竞争较强,禁牧会导致草地生态系统的植物种群生态位发生分化。禁牧处理下,限制种群生长的因素主要是自然资源,因此家畜的游走、践踏等行为不作为限制植物种群生长的关键因素时,禾草类植物的生存空间会进一步增大。
禁牧会改变温性草原植物物种的生态位重叠值[20]。本研究发现,禁牧处理和自然放牧处理下草沙蚕、白草和矮生嵩草与其他物种所组成种对的生态位均有重叠(图1),说明此类种对的资源利用状态相似[20]。在禁牧处理中,笔直黄芪和肉果草之间的生态位重叠值较高,但是笔直黄芪和肉果草的生态位宽度较窄,可能是因为种对具有相似的生物学特性,导致其在对生态环境的需求上产生互补[8],与高寒草地的养分含量等资源匮乏相关。进一步分析发现禁牧处理下的植物群落优势物种生态位重叠程度增加,表明种间竞争激烈,植物群落结构不稳定[21]。
禁牧会改变群落的总体联结性[22]。本研究中,禁牧群落总体表现为显著负联结(Vr=0.43),与χ2检验结果相符合,进一步验证了种间竞争的存在,这与吴姣姣等[9]在拉萨河谷温性草原禁牧群落主要物种间的总体联结性为负的结果相一致。表明无放牧干扰情况下,限制群落内植物种群生长的关键因素为环境资源,且主要物种对环境的适应性存在差异,主要物种之间对光照、水分及土壤养分等资源存在一定竞争关系,同时个别优势物种对其他物种的生长具有抑制作用[23]。本研究所有种对中,19 个种对表现出负联结性,9个种对表现出正联结性,进一步验证了群落中种间竞争的存在,此外白草和矮生嵩草、丝颖针茅和藏橐吾均表现出明显的负关联性,表明白草在禁牧群落中占据优势地位,并且对矮生嵩草、丝颖针茅和藏橐吾的生长具有明显的抑制作用。自然放牧处理下群落总体表现为显著正联结,与χ2检验结果相符合,这与刘菊红等[8]在荒漠草原重度放牧群落主要物种间的总体联结性为正相一致。可能是家畜的采食、游走等行为活动限制植物群落的发展,降低高大植物的竞争强度,增加下层低矮植物的物种数[11],且减弱优势物种对环境资源的竞争,植物种群在长期进化过程中形成相互促进的种间关系以抵御放牧干扰,形成矮生植物和中生植物共存的分布格局,故群落的种间关系由互相竞争转变为互利共存[21]。由分析联结系数Ac可以得知,禁牧使得丝颖针茅和白草的联结性转变为负联结,表明禁牧增强了此类物种之间对资源的竞争。由此证明,禁牧增强物种之间的竞争排斥力,使得植物群落的种间关系由互利共生甚至可能转变为相互强竞争[24]。
禁牧会改变种间的关联程度[22]。本研究发现,禁牧处理下优势种(草沙蚕、高山嵩草、白草、丝颖针茅)之间均有生态位重叠(图1),且优势种之间的Oi指数较大(图4),表明优势种利用资源的关联程度提高,共同出现的几率较大[24]。进一步分析发现,禁牧处理下,高山嵩草和笔直黄芪、高山嵩草和藏橐吾、白草和藏橐吾、笔直黄芪和藏橐吾等种对之间无重叠(Oi=0),表明禁牧条件下这些物种共同出现的几率小。放牧条件下优势种之间的Oi值降低且生态位重叠程度也下降,表明放牧导致优势种之间的资源竞争情况减弱,但伴生种与其他物种之间的资源竞争情况增强,说明各优势种在群落中共同出现的几率降低,而伴生种与其他物种在群落中出现的几率增大[9]。禁牧处理下,Oi值在0.5以上的种对数有16个,占总数的57.14%,相对较高,表明禁牧群落中,各植物物种间关系较为紧密。可能是由于禁牧封育会形成以高大禾草和杂草为主的群落,上层植物对光照和水分等环境资源的竞争加剧[25],而对自然资源的争夺成为限制种群发展的关键因素,且禁牧进一步导致土壤温湿度及其主要成分发生改变,有效改善土壤成分和通气状况,也会影响腐殖质的降解速度,生态位类似的物种会对环境资源存在激烈竞争关系[26],群落的结构也会发生变化[27],因此禁牧群落中物种多样性增加的同时种间竞争也更为激烈。
4 结论
禁牧使得草沙蚕、高山嵩草、白草和丝颖针茅生态位宽度增加,而矮生嵩草、笔直黄芪、藏橐吾和肉果草的生态位宽度降低,根据生态位互补效应,禾草类生存空间增大。同时使主要物种生态位重叠值增加,加剧种间竞争程度,草地植物群落的恢复向正方向发展。禁牧改变种间关联程度,使植物群落整体表现为负联结,进一步使种间关系更加紧密。
