OmniLogTM微生物鉴定系统在青海高原野生型鼠疫噬菌体宿主谱检测中的应用研究
2024-02-23李存香祁芝珍张青雯赵海红游陪松杨建国吴海生冯建萍
李存香,祁芝珍,张青雯,赵海红,马 龙,游陪松,杨建国,吴海生,冯建萍
噬菌体是地球上最具生物多样性的生命体,通常超过细菌密度的一个数量级[1]。作为细菌的病毒性捕食者,噬菌体在微生物进化、生态学、细菌性疾病预防和治疗、细菌鉴定与疾病诊断等方面扮演着重要角色[2-3]。特定细菌的噬菌体总体上类似,即细菌能够被噬菌体特异性裂解,其特异性主要源于噬菌体与其宿主菌细胞壁上特异性受体的相互识别与吸附[4],而差异性主要在于宿主的特异性范围[5]。不同噬菌体的吸附方式和特异性分子生物学机制各异,但目的都是为了将其核酸注射到宿主菌中,持续循环侵染宿主菌[6]。鼠疫噬菌体作为鼠疫耶尔森菌的病毒,除了大多数在特定温度下可以裂解多数的鼠疫菌株,还有部分噬菌体能裂解某些大肠埃希氏菌、假结核耶尔森菌、痢疾志贺氏杆菌和猪霍乱沙门氏菌等[7-9]。噬菌体宿主的定义和结果因检测方法的不同而不同[10-11]。自然环境中分离噬菌体时,除了分离时所用的宿主菌,观察它对其他近属的菌株是否感染,都需要通过不同的培养方法来检测。国内外噬菌体敏感性试验常见的培养方法大致分为两类:半固体培养法和液体检测法。另外,各种微生物鉴定系统也已在微生物鉴定和代谢研究、微生物群落分析等方面被广泛应用[12-13],其中国内外研究者曾利用OmniLogTM微生物鉴定系统间接评估了炭疽芽孢杆菌及其噬菌体抗性突变体分别与6株炭疽芽孢杆菌噬菌体/噬菌体鸡尾酒/组合抗生素存在环境下的细菌生长动力学特点,利用Bioscreen C微生物生长分析仪测定了铜绿假单胞菌噬菌体的活性、蜡样芽孢杆菌噬菌体的宿主谱,都具有较好的应用价值[14]。本研究团队同样基于OmniLogTM微生物鉴定系统建立了鼠疫噬菌体在微量液体培养基中的生长方法[15],同时利用该方法成功检测了青海高原分离的3株野生型鼠疫噬菌体的宿主谱,为今后噬菌体生态学研究和基于宿主范围噬菌体分类研究提供科学依据。
1 材料与方法
1.1 材 料
1.1.1 实验菌株 鼠疫疫苗株EV76、614F和假结核耶尔森菌PTB3、PTB5、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2由青海省地方病预防控制所鼠疫菌专业实验室提供。
1.1.2 噬菌体 072204号鼠疫噬菌体分离自2019年青海省同德县鼠疫疫源地喜马拉雅旱獭盲肠样本,效价为10-6,087号鼠疫噬菌体分离自2019年青海省天峻县鼠疫疫源地喜马拉雅旱獭盲肠样本,效价为10-6,476号鼠疫噬菌体分离自2020年青海省天峻县鼠疫疫源地喜马拉雅旱獭盲肠样本,效价为10-7。诊断用鼠疫噬菌体由青海省地方病预防控制所鼠疫菌专业实验室提供。
1.1.3 仪器 恒温培养箱、空气浴振荡培养箱和OmniLogTM微生物鉴定系统由青海省地方病预防控制所鼠疫菌专业实验室提供。
1.1.4 培养基和试剂 LB液体培养基、LB固体培养基由青海省地方病预防控制所鼠疫菌专业实验室培养基室制备。四唑类染液购自于美国OmniLog公司。
1.2 方 法
1.2.1 点滴法 取6个LB固体培养基分别划分为6等份,每一等份接种1株复苏的试验菌株,共6株菌。分别取已制备的15 μL被检噬菌体点滴于已接种的试验菌株上,使其自然渗入培养基。每株噬菌体培养2个平皿,分别置于28 ℃和37 ℃恒温培养箱,20~24 h后观察噬菌斑。
1.2.2 基于OmniLogTM系统微量液体检测法 第2至6列孔加入 90 μL LB 液体培养基。第1列孔中加入实验用噬菌体,按照10倍梯度稀释至第5列孔,第5列孔弃去10 μL噬菌体稀释混合液,参照文献[15]检测3株野生型鼠疫噬菌体对2株鼠疫耶尔森菌(鼠疫疫苗株EV76、614F)和4株非鼠疫耶尔森菌(假结核耶尔森菌PTB3、PTB5、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2)的裂解情况。
2 结 果
2.1 点滴法培养结果 通过点滴法检测3株野生型鼠疫噬菌体的宿主谱,发现以鼠疫疫苗株EV76、614F为宿主菌,476号鼠疫噬菌体在28 ℃和37 ℃时琼脂培养基上产生清晰的噬菌斑,072204号鼠疫噬菌体在28 ℃和37 ℃时均不能在琼脂培养基上形成可见的斑块,这并不意味着缺乏生产性感染(通过双层琼脂平板法经37 ℃时培养24 h可见不透明的斑)。087号鼠疫噬菌体在28 ℃时不能在琼脂培养基上形成可见的斑块,但37 ℃时在琼脂培养基上形成可见的斑块。以4株非鼠疫耶尔森菌拟为宿主菌,476号噬菌体对假结核耶尔森菌PTB5有裂解作用,对其他3株非鼠疫耶尔森菌不裂解。087号和072204号噬菌体对4株非鼠疫耶尔森菌均未发现有裂解现象。如图1、图2。

图1 3株鼠疫噬菌体经点滴法28 ℃作用24 h的生长情况Fig.1 Plaque formation after interaction of three plague phages with six intestinal bacteria, determined with the micro spot method on semisolid medium at 28 ℃ for 24 h
2.2 OmniLogTM系统检测结果 3株噬菌体经OmniLogTM系统检测其宿主谱结果显示,以鼠疫疫苗株EV76、614F为试验菌,3个检测板实验组第1列生长曲线几乎呈一条横线,且细菌浓度对数值不超过100,后续每孔中根据不同噬菌体效价的降低,宿主菌依次呈现对应S型生长曲线;试验孔中四唑类染料的颜色随着噬菌体数量的减少和宿主菌数量的增多而逐渐加深,说明3株噬菌体均能够感染2株鼠疫疫苗株。以假结核耶尔森菌PTB5为试验菌,仅476号噬菌体实验组第1列生长曲线几乎呈一条横线,且细菌浓度对数值不超过100,后续每孔中根据不同噬菌体效价的降低,宿主菌依次呈现对应的S型生长曲线,试验孔中四唑类染料的颜色随着噬菌体数量的减少和宿主菌数量的增多而逐渐加深;而087号和072204号噬菌体实验组每一行的前5孔与细菌对照第6孔均呈现几乎对应的相同S型曲线,且试验孔中四唑类染料的颜色与对应对照孔列相近,说明476号噬菌体能感染假结核耶尔森菌PTB5,087号和072204号噬菌体不能感染假结核耶尔森菌PTB5。以假结核耶尔森菌PTB3、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2为试验菌,3个检测板实验组每一行前5孔与细菌对照第6孔均呈现几乎对应的、相近的S型曲线,说明3株噬菌体不能感染假结核耶尔森菌PTB3、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2。由此可见,上述检测方法可以作为生物学特性之效价监测和宿主谱检测的依据之一。如图3、图4、图5。
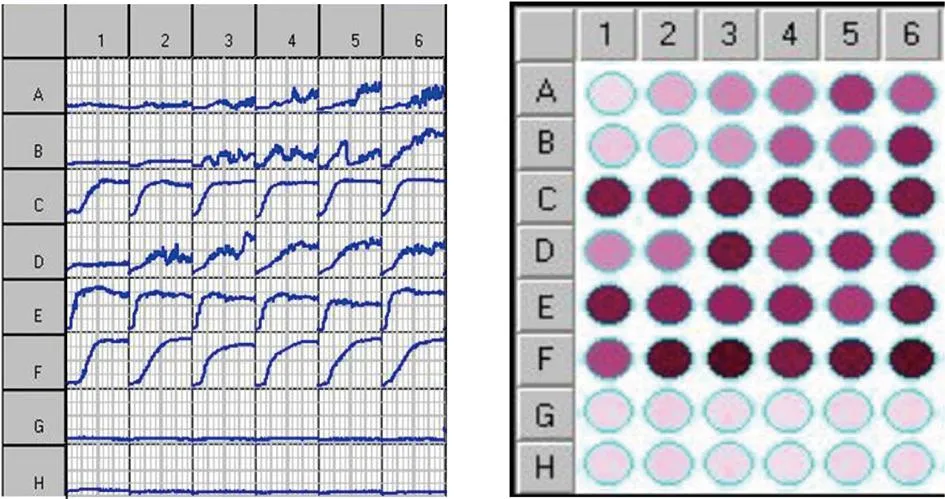
注: A、B、C、D、E、F、G、H行分别代表鼠疫弱毒菌EV76、鼠疫弱毒菌614F、假结核耶尔森菌PTB3、假结核耶尔森菌PTB5、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2、新鲜LB液体培养基、鼠疫噬菌体476。1、2、3、4、5列分别代表被测鼠疫噬菌体不同的稀释梯度,6列代表对应排的菌株对照组。图3 鼠疫噬菌体476与6株肠道菌株经OmniLogTM微生物鉴定系统33 ℃作用48 h后的生长情况Fig.3 Growth curves and chromatic graph for interactions of six intestinal bacteria with plague phage 476, determined with the OmniLogTM microbial identification system, at 33 ℃ for 48 h

注: A、B、C、D、E、F、G、H行分别代表鼠疫弱毒菌EV76、鼠疫弱毒菌614F、假结核耶尔森菌PTB3、假结核耶尔森菌PTB5、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2、新鲜LB液体培养基、鼠疫噬菌体087。1、2、3、4、5列分别代表被测鼠疫噬菌体不同的稀释梯度,6列代表对应排的菌株对照组。图4 鼠疫噬菌体087与6株肠道菌株经OmniLogTM微生物鉴定系统33 ℃作用48 h后的生长情况Fig.4 Growth curves and chromatic graph of interactions of six intestinal bacteria with plague phage 087, determined with the OmniLogTM microbial identification system, at 33 ℃ for 48 h

注: A、B、C、D、E、F、G、H行分别代表鼠疫弱毒菌EV76、鼠疫弱毒菌614F、假结核耶尔森菌PTB3、假结核耶尔森菌PTB5、大肠杆菌V517、小肠结肠炎耶尔森菌52302-2、新鲜LB液体培养基、鼠疫噬菌体072204。1、2、3、4、5列分别代表被测鼠疫噬菌体不同的稀释梯度,6列代表对应排的菌株对照组。图5 鼠疫噬菌体072204与6株肠道菌株经OmniLogTM微生物鉴定系统33 ℃作用48 h后的生长情况Fig.5 Growth curves and chromatic graph of interactions of six intestinal bacteria with plague phage 072204, determined with the OmniLogTM microbial identification system, at 33 ℃ for 48 h
3 讨 论
鼠疫耶尔森菌作为鼠疫的病原体,在生物进化处于有利的生活条件下表现为一般性适应,在不利的生活条件下表现为特异性适应[16]。鼠疫噬菌体作为鼠疫的细菌病毒,不同环境分离的噬菌体有相对于其分离地环境的生理特征。噬菌体的宿主范围作为噬菌体的核心特征之一,它是指能成功感染宿主的分类多样性[17],由整个感染周期中噬菌体和宿主之间的一系列分子相互作用决定的[18]。从微生态学方面来说,当鼠疫噬菌体有除鼠疫菌外的其它宿主菌存在时,鼠疫噬菌体可以不依赖鼠疫菌生长繁殖。噬菌体宿主谱的维持和改变促进了自然界微生物基因的多样性和基因的交换[19]。虽然许多经过充分研究的噬菌体模型似乎显示出较窄的宿主范围,但近期的生态学和宏基因组学研究表明,噬菌体可能具有从窄到宽的特异性[18-19]。噬菌体与宿主菌相互作用的机制及其进化对于理解噬菌体生态学和噬菌体在临床、工业和生物技术中的应用具有非常重要的意义[2]。
本研究采用点滴法检测087号和072204号鼠疫噬菌体的宿主谱,结果显示二者在28 ℃时不裂解鼠疫疫苗株EV76、614F,但在37 ℃时087号噬菌体点滴于固体培养基上显示噬菌斑,072204号噬菌体点滴于固体培养基上不显示噬菌斑,但在双层琼脂培养基上显示不透明噬菌斑。由此可见,087号和072204号噬菌体培养生长时具有温度依赖性。476号噬菌体的裂解谱结果显示,在28 ℃和37 ℃时对鼠疫疫苗株EV76、614F、假结核耶尔森菌PTB5均裂解,即有明显的噬菌斑。基于OmniLogTM微生物鉴定系统3株噬菌体经微量液体培养法33 ℃培养,生长曲线图及其显色图发现476号、087号和072204号噬菌体仅对鼠疫疫苗株EV76、614F均裂解,且476号噬菌体对假结核耶尔森菌PTB5也裂解。由此可见,基于点滴法在遇到如37 ℃不能产生可见噬菌斑的072204号噬菌体时,不能及时判断其是否存在裂解能力,而基于OmniLogTM检测系统的微效价宿主范围测定法不需要目视检查噬菌斑块,可以在单一试验中提供噬菌体宿主范围和毒力特性的信息。OmniLogTM检测系统在整个试验过程中实时、连续和自动化监测噬菌体感染细菌生长方面具有明显的优势。基于OmniLogTM检测系统的噬菌体宿主范围测定法可以作为一种替代传统检测宿主范围的测定方法,能够提供更多关于菌株间高分辨率的抑菌信息。另外,本研究利用OmniLogTM检测系统监测3株鼠疫噬菌体对6株细菌的裂解能力时,发现476号噬菌体具有宽宿主谱特性,这不仅在自然界微生态学研究中具有重要意义,而且可能为人类抗细菌感染提供另一种选择性治疗的手段。
利益冲突:无
