草地早熟禾根茎发育不同阶段及不同部位糖代谢差异
2024-01-06刘敏洁刘文辉李军乔魏小星马晖玲
杨 伟,刘敏洁*,刘文辉,李军乔,魏小星,马晖玲
(1.青海民族大学生态环境与资源学院,青海 西宁 810007; 2.青海省特色经济植物高值化利用重点实验室,青海 西宁 810007;3.青海省畜牧兽医科学院/青海省青藏高原优良牧草种质资源利用重点实验室,青海 西宁 810016;4.甘肃农业大学草业学院,甘肃 兰州 730070)
根茎是一种地下变态茎,不仅可以储存养分,还可以不断形成新的根茎占据地下空间。根茎中的一部分在条件适宜的情况可形成新的植株,从而达到繁殖目的[1-2]。这种繁殖方式使根茎类植物具有很强的种群繁衍能力,通常这类植物都具有较强的适应性[3]。
碳水化合物是植物的主要有机成分之一,包括结构性碳水化合物和非结构性碳水化合物。其中,葡萄糖、果糖、蔗糖和淀粉等是参与植物生命活动的主要非结构性碳水化合物[4],它们还可以促进部分结构碳水化合物的合成,例如木质素和纤维素等[5-6]。光合作用产生的蔗糖通过韧皮部运输到根茎后,一部分在转化酶的作用下裂解为葡萄糖和果糖,从而为根茎的扩展提供能量[7-8];另一部分在蔗糖合酶的作用下产生果糖和腺苷二磷酸葡萄糖(ADP-Glucose,ADPG),然后在腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPase)和淀粉合酶的催化下合成淀粉,最后将淀粉储存在根茎中作为储备养分[9-11]。当根茎快速繁殖时,淀粉又会被降解为蔗糖,为植物生长提供能量[12]。近年来,关于根茎发育的相关研究愈来愈多,水稻(Oryzasativa)根茎蔗糖和淀粉主要分布在靠近植株的根茎基部,而葡萄糖和果糖在根茎尖端含量更高[13]。此外,蔗糖可以增加水稻根茎的生长速度,但在发育后期,过高的蔗糖浓度则会抑制新植株的形成[13-14]。花秆实心竹(Fargesiayunnanensis)根茎在形成新笋的过程中,其蔗糖和淀粉含量大幅减少[7],而高羊茅(Festucaarundinacea)根茎中的蔗糖含量和鸢尾(Iris tectorum)根茎中的淀粉含量同样随发育时间逐渐增加[12,15]。因此,根茎中糖的积累程度是影响根茎发育的主要因素,并且糖很可能以信号通路的方式参与调节根茎的发育。另有研究表明Poapratensis根茎中的SPS活性与蔗糖含量呈显著正相关,而分解酶与蔗糖含量呈显著负相关[4]。大豆(Glycinemax)果实成果过程中,SS和SPS活性与蔗糖含量呈显著正相关,而SAI活性与蔗糖含量呈显著负相关[16]。这些研究表明蔗糖和淀粉是根茎类植物储存物质的主要形式。目前对草地早熟禾的研究大多集中在地上部分,虽然关于其根茎糖代谢的研究也已有报道,但这些研究把初级根茎当作一个整体进行研究,研究初级根茎碳水化合物不同生育期之间差异,其根茎发育与碳水化合物代谢之间的关系尚不清楚[17-20]。
青海扁茎早熟禾是针对青藏高原高海拔、高寒的恶劣生长环境选育出的抗寒、耐旱、根茎繁殖快、固土保水能力强的为数不多的优良牧草品种之一。它在-35℃的低温下能安全越冬,适合在海拔高度2 000~4 100 m、年降水量400 mm左右的高寒牧区旱作种植。此外,在我们多年田间驯化过程中发现,有1份野生草地早熟禾种质(SN)生长比较稳定,并且其根茎繁殖能力较弱。因此,本研究将强根茎型品种青海扁茎早熟禾(QH)和弱根茎型材料野生草地早熟禾的根茎发育期糖代谢差异进行比较,研究初级根茎发育过程中碳水化合物含量及相关酶活性动态变化,以及初级根茎即将形成根茎子株时期碳水化合物从根茎基部到顶端的分布情况,从而更好地了解糖的储存和分解与根茎发育的关系。
1 材料与方法
1.1 供试材料
青海扁茎早熟禾和野生草地早熟禾均由青海省畜牧兽医科学院提供,详情见表1。

表1 供试材料信息表Table 1 Information sheet of the materials in experiment
1.2 试验地概况
试验地位于青海省海北州西海镇(101°45′E,36°49′N;海拔3 156 m)的国家牧草种质资源苗圃(青藏高原)。试验站气候类型为高原大陆性气候,2019年年平均气温0.5℃,无绝对无霜期,降雨量集中在7-9月,并且年平均降水量仅为369 mm,年蒸发量1 400 mm[17]。土壤为板栗土(表2),2019年雨热同期(图1)。

图1 降雨量及平均温度变化Fig.1 Monthly precipitation and mean temperature changes

表2 试验地土壤特征Table 2 Soil characteristics of the experimental site
1.3 试验设计
2019年6月10日,单株种植两份材料,株距为1 m,试验地面积为1 200 m2(长30 m,宽40 m),每份材料占地面积为600 m2。定期进行除草,并于9月份进行间苗,每穴只保留一株幼苗。整个试验期间不施肥、不灌溉。
在前期试验中发现QH和SN形成新植株时,具有7~9个节间的初级根茎分别有76%和75%(图2)。这表明QH和SN根茎向新植物的转化发生在7~9节间的阶段。因此,在2020年拔节期采样时分为两种方式:
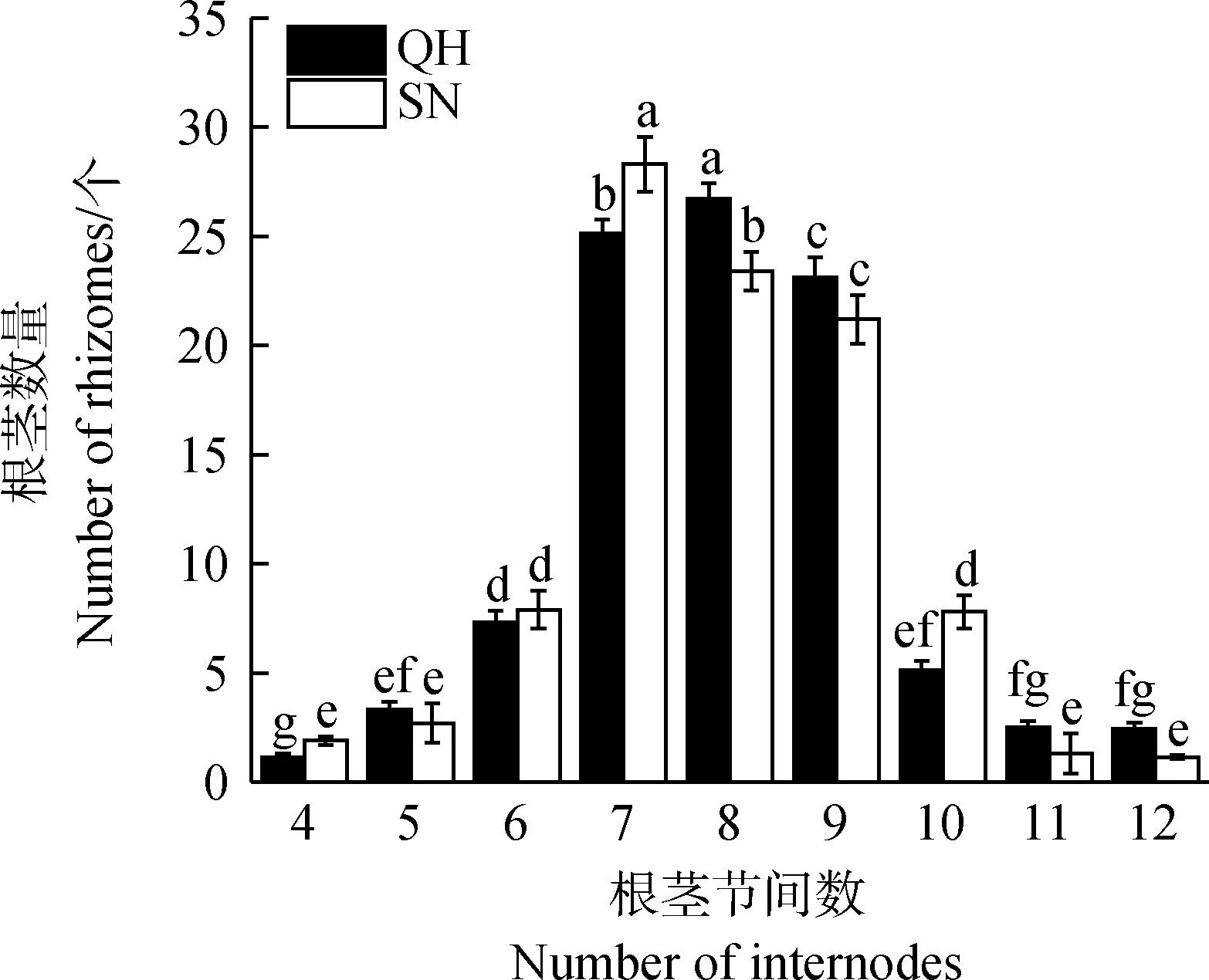
图2 弯曲生长阶段根茎具有的节间数Fig.2 Number of internodes of rhizomes during bending stage注:不同小写字母表示同一材料不同根茎节间数之间差异显著(P<0.05)Note:Different lowercase letters indicate a significant difference among different number of rhizomes internodes of the same test material (P<0.05)
(1)收集不同发育时期的初级根茎:T1,根茎芽形成初期;T2,具有1个根茎节间的初级根状茎;T3,具有3个根茎节间的初级根状茎;T4,具有5个根茎节间的初级根状茎;T5,根茎向上弯曲即将形成根茎子株阶段。具体采样方式见图3。

图3 采样示意图Fig.3 Sampling diagram of the development of primary rhizome
(2)为进一步明确根茎发育过程中碳水化合物的变化,收集T5阶段的初级根茎,分为以下四个部分:P1,从基部开始第1节间;P2,从基础开始第3节间;P3,从基础开始第5节间;P4,根茎向上弯曲的部分。具体采样方式见图4。

图4 采样示意图Fig.4 Diagram for the sampling sections of primary rhizome
1.4 测定指标
1.4.1非结构性碳水化合物含量 蔗糖、果糖、葡萄糖和淀粉含量测定方法参考牛奎举等[6,21-22]的方法。
1.4.2蔗糖代谢酶活性 蔗糖磷酸合酶(Sucrose Phosphate Synthase,SPS)、蔗糖合酶-合成方向(Sucrose Synthase at Synthesis Direction,SS)、可溶性酸性转化酶(Soluble Acid Invertase,SAI)和中性转化酶(Neutral Invertase,NI)活性测定方法参考牛奎举等的方法[6,23]。
1.4.3淀粉代谢酶活性 腺苷二磷酸葡萄糖焦磷酸化酶(ADP-Glucose Pyrophosphorylase,AGPase)、可溶性淀粉合酶(Soluble Starch Synthase,SSS)、淀粉磷酸化酶(Starch Phosphorylase,SP)和颗粒结合淀粉合酶(Granule-Bound Starch Synthase,GBSS)活性测定参考Wang等[7]的方法。
1.5 数据分析
使用Microsoft Excel 2019对数据进行整理,采用SPSS 21.0对于同一材料不同发育时期初级根茎的碳水化合物变化和同一材料根茎弯曲阶段不同根茎节间碳水化合物变化进行单因素方差分析,采用Duncan法在0.05水平上进行多重比较;对于同一发育时期不同材料间碳水化合物变化和同一根茎节间不同材料间碳水化合物变化进行独立样本t检验,最后用Origin 2021绘图。
2 结果分析
2.1 初级根茎不同发育时期/部位碳水化合物含量变化
不同发育时期初级根茎的碳水化合物含量差异显著(图5,P<0.05)。其中,两份材料初级根茎蔗糖和淀粉含量随着生长阶段的变化持续升高(T1-T5)(P<0.05)。T1阶段,QH和SN初级根茎蔗糖含量为5.36 mg·g-1和4.93 mg·g-1,两者间差异不显著,但从T2阶段开始,QH蔗糖含量显著高于SN(P<0.05)。T5阶段,QH初级根茎蔗糖含量达11.67 mg·g-1,较T1阶段增加了117.78%;SN初级根茎蔗糖含量为9.68 mg·g-1,较T1阶段增加了96.55%;并且此时QH蔗糖含量比SN高出20.48%(P<0.01)。此外,T1阶段,QH和SN初级根茎淀粉含量分别仅为5.56 mg·g-1和4.34 mg·g-1,T5阶段分别增加到8.75 mg·g-1和7.70 mg·g-1,分别较T1增加了57.37%和77.50%,并且整个发育阶段QH淀粉含量显著高于SN(P<0.05)(图5A,B)。

图5 初级根茎不同发育时期碳水化合物含量变化Fig.5 Changes of the carbohydrate content of primary rhizome at different development stages注:不同小写字母表示同一材料不同发育阶段间差异显著(P<0.05),*,**和***分别代表同一处理下不同材料间在P<0.05、P<0.01和P<0.001水平上差异显著。下同Note:Different lowercase letters indicate a significant difference among different developmental stages of the primary rhizome of the same tested species (P<0.05). *,** and *** represent significant differences between different tested species under the same developmental stage at the P<0.05,P<0.01,and P<0.001 levels,respectively. The same as below
QH果糖含量在T1-T3阶段逐渐降低,在T4期突然增加(P<0.05),之后在T5阶段又呈现断崖式下降(P<0.05)(图5 C),而SN果糖含量呈现持续降低的趋势(图5 D)。并且整个生长阶段内,QH果糖含量显著高于SN(P<0.05),其中T4阶段QH果糖含量达10.83 mg·g-1,较SN高出61.27%。QH和SN根茎葡萄糖含量在T1阶段分别为6.89 mg·g-1和6.06 mg·g-1,T5阶段与T1相比分别降低了40.76%和31.85%,并且T1-T3阶段QH葡萄糖含量显著高于SN(P<0.05)。
由图6可知,初级根茎向上弯曲阶段不同部位间碳水化合物含量差异显著(P<0.05)。其中,初级根茎向上弯曲阶段不同部位间蔗糖和淀粉含量具体表现为初级根茎基部节间(P1)含量最高,越靠近初级根茎弯曲部位越低(P4)(图6 A,B)。P1节间,QH和SN蔗糖含量分别为14.92 mg·g-1和11.34 mg·g-1,P4分别较P1降低了72.64%和69.06%;淀粉含量分别为11.32 mg·g-1和10.30 mg·g-1,P4分别较P1降低了85.95%和85.92%。同时,在P1-P3节间,QH蔗糖含量均显著高于SN(P<0.05),而淀粉含量QH仅在P1节间显著高于SN(P<0.05)。
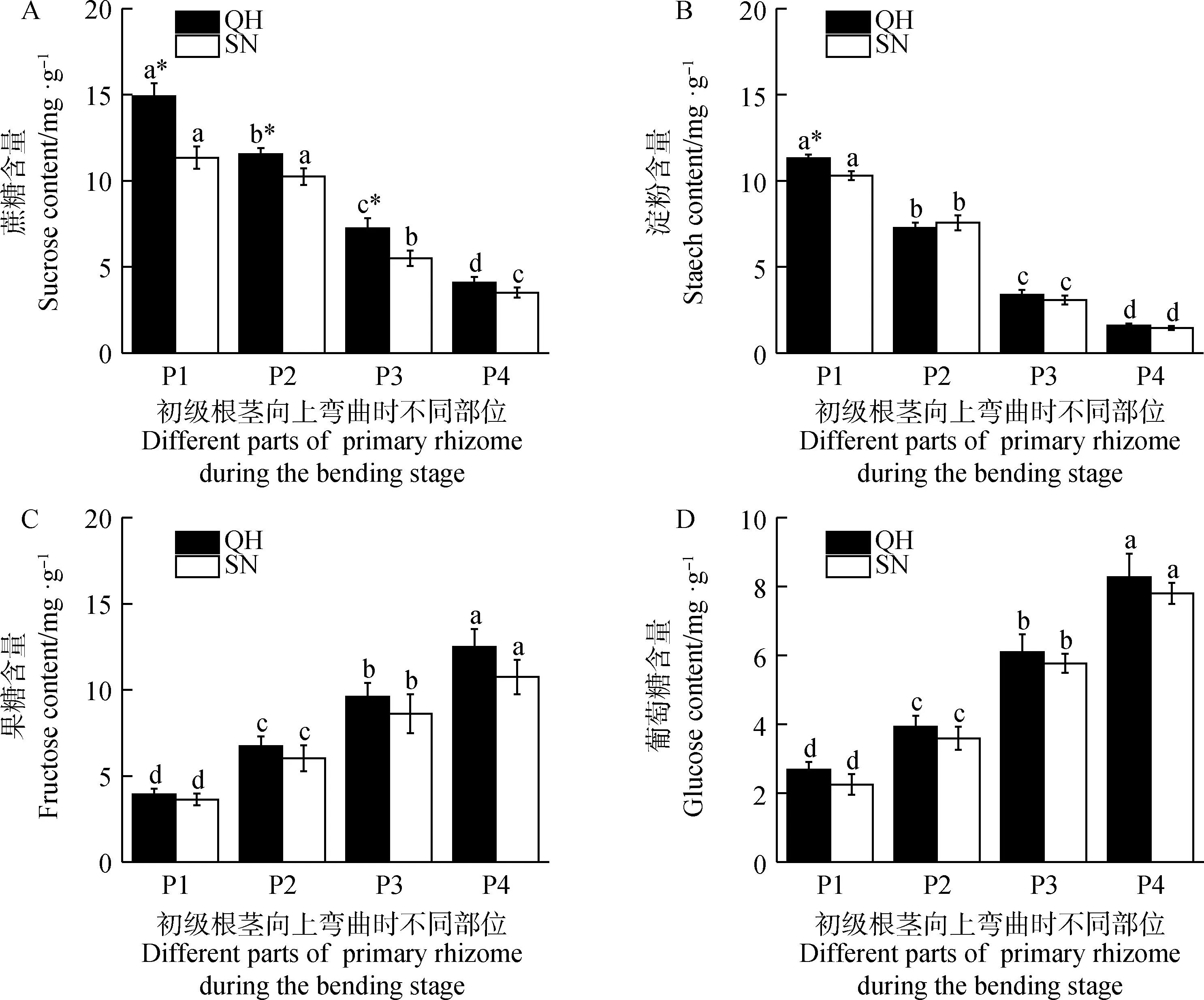
图6 初级根茎向上弯曲阶段不同部位碳水化合物含量变化Fig.6 Changes of carbohydrate content of different sections of primary rhizome during its bending stage
初级根茎向上弯曲阶段不同部位间果糖和葡萄糖含量表现为初级根茎基部节间(P1)含量最低,越靠近初级根茎弯曲部位越高(P4)(图6 C,D)。P4节间,QH和SN的果糖含量高达12.50 mg·g-1和10.75 mg·g-1,分别较P1增加了217.08%和195.33%;葡萄糖含量达到8.27 mg·g-1和7.80 mg·g-1,分别较P1增加了207.81%和246.31%。同时,QH和SN各个部位节间果糖和葡萄糖含量无显著差异。
2.2 根茎不同发育时期/部位蔗糖代谢酶活性变化
由图7可知,不同发育时期初级根茎的蔗糖代谢酶活性差异显著(P<0.05)。其中,T1-T5时期,QH初级根茎SPS活性先升高后降低,在T3时期达到最大值1 248.22 μg·min-1·g-1(图7 A)。而SN初级根茎SPS活性仅在T2时期较T1降低了15.12%,其他时期间无显著差异。同时,在T1-T4时期,QH初级根茎SPS活性显著高于SN(P<0.05),尤其在T3时期,较SN高出61.40%。
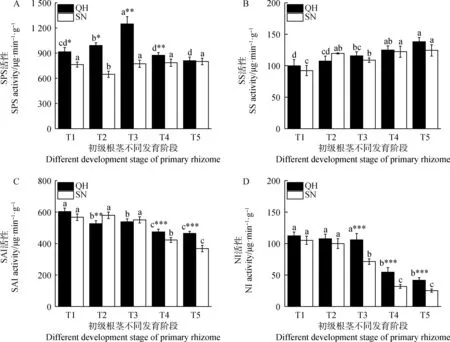
图7 初级根茎不同发育时期蔗糖代谢相关酶活性变化Fig.7 Changes of sucrose metabolizing-related enzyme activities in primary rhizome at different development stages
SS活性在T1-T5时期呈现逐渐升高的趋势(图7B),T5时期QH和SN根茎SS活性分别为138.04 μg·min-1·g-1和124.40 μg·min-1·g-1,较T1时期升高了38.39%和34.93%,并且各个时期两份材料间SS活性无显著差异(P<0.05)。随着初级根茎整体发育,蔗糖合成相关酶活性均有所升高,并且,在此过程中以SPS活性占主导地位(QH和SN初级根茎各发育时期SPS活性均值为860.84 μg·min-1·g-1,而SS活性均值仅为115.29 μg·min·g-1)。
T1-T5时期,QH和SN初级根茎SAI和NI活性呈现降低趋势(P<0.05)(图7 C,D),并且均在T4时期活性下降幅度较大。T5时期QH和SN根茎SAI活性分别为464.55和367.99 μg·min-1·g-1,与T1相比降低了23.07%和35.17%;而NI活性为41.79和25.34 μg·min-1·g-1,与T1时期相比,分别下降了62.81%和75.91%。此外,T2时期QH初级根茎SAI活性显著低于SN(P<0.05),T4和T5时期,QH根茎SAI活性和NI活性显著高于SN(P<0.05)。总体来看,初级根茎发育后期,QH蔗糖分解酶活性高于SN(P<0.05),并且蔗糖分解相关酶活性在初级根茎发育后期均有不同程度的降低。
由图8可知,初级根茎向上弯曲阶段不同部位间蔗糖代谢酶活性同样差异显著(P<0.05)。其中,初级根茎向上弯曲阶段不同部位间SPS活性和SS活性变化和蔗糖含量十分相似(图8 A,B),具体表现为初级根茎基部节间(P1)活性最高,越靠近初级根茎弯曲部位越低(P4)。P1节间,QH和SN初级根茎SPS活性分别为714.52和621.63 μg·min-1·g-1,P4节间分别较P1降低了72.79%和72.00%;QH和SN初级根茎SS活性分别为90.98和100.93 μg·min-1·g-1,P4节间分别较P1降低了64.51%和64.49%。同时,在P1和P2节间,QH初级根茎SPS活性均显著高于SN(P<0.05),但SS活性无显著差异。
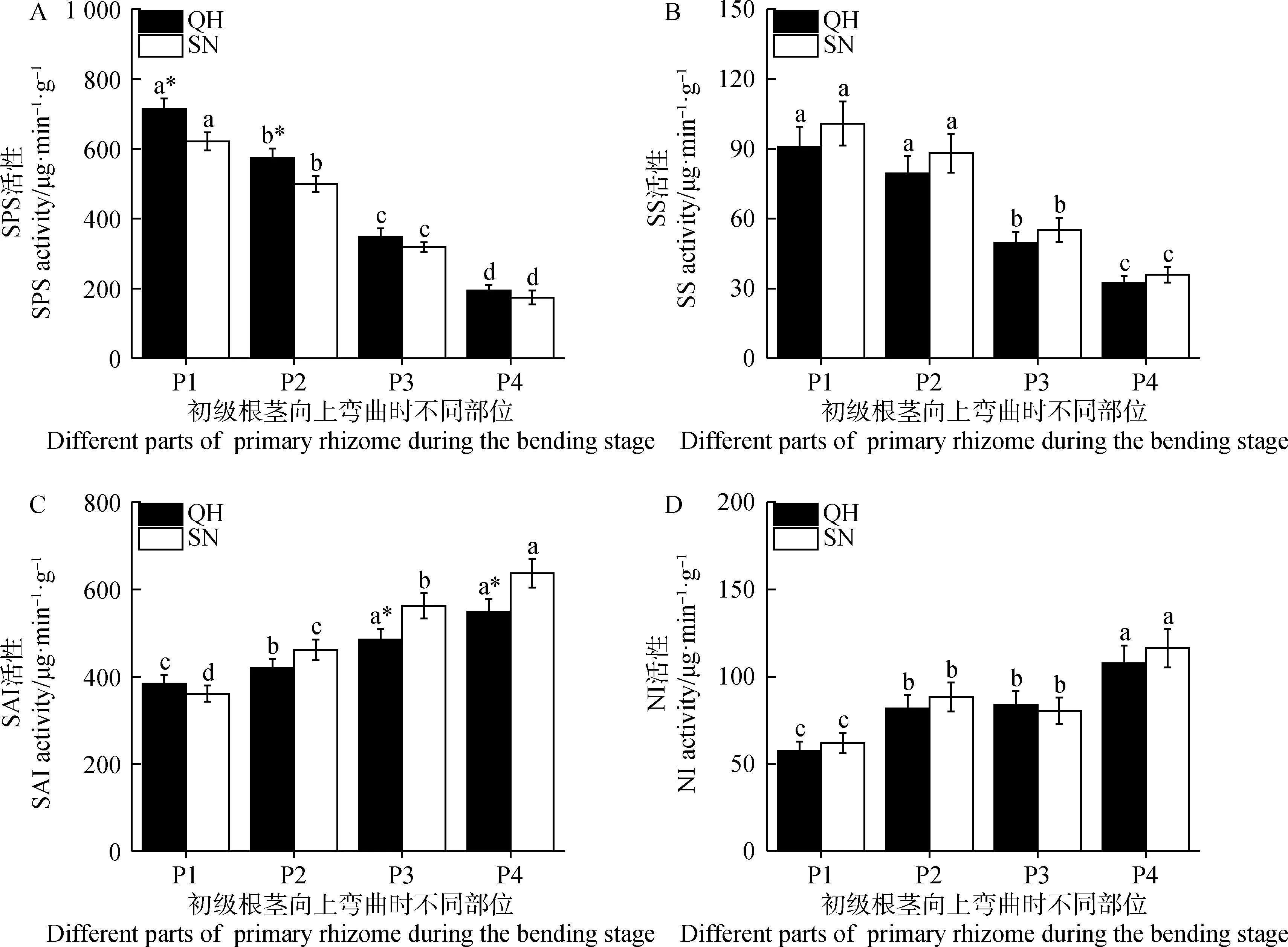
图8 初级根茎向上弯曲阶段蔗糖代谢相关酶活性变化Fig.8 Changes of sucrose metabolizing-related enzyme activities in different sections of primary rhizome during its bending stage
QH和SN初级根茎弯曲阶段不同部位SAI和NI活性分布与SPS和SS活性完全相反,P1节间活性最低,P4节间活性最高(图8C,D)。P4节间QH和SN初级根茎SAI活性分别为549.03和636.88 μg·min-1·g-1,较P1增加了42.90%和76.34%;NI活性分别为107.71和116.33 μg·min-1·g-1,较P1增加了87.72%和87.66%。此外,初级根茎弯曲阶段各部位间,QH和SN根茎SAI和NI活性基本无显著差异。
2.3 根茎不同发育时期/部位淀粉代谢酶活性变化
由图9可知,不同发育时期初级根茎的淀粉代谢酶活性差异显著(P<0.05)。其中,T1-T5时期,QH初级根茎AGPase活性先升高后降低,在T3时期达到最大值188.63 μg·min-1·g-1(图9 A),较T1时期升高了27.05%。SN初级根茎AGPase活性则表现为持续升高的趋势,T5时期其活性为167.57 μg·min-1·g-1,较T1时期升高了28.56%。同时,在T2和T3时期,QH初级根茎AGPase活性显著高于SN(P<0.05)。

图9 初级根茎不同发育时期淀粉代谢相关酶活性变化Fig.9 Changes of starch metabolizing-related enzyme activities in primary rhizome at different development stages
SSS活性在T1-T5时期呈现逐渐升高的趋势(图9 B)。其中QH初级根茎SSS活性在T3时期开始显著增加,T5时期SSS活性为62.54 μg·min-1·g-1,较T1时期升高了78.05%。而SN初级根茎SSS活性在整个发育过程中变化幅度较小,T5时期较T1时期仅增加了25.76%。同时T3-T5时期,QH初级根茎SSS活性显著高于SN(P<0.001),尤其T4时期,QH初级根茎SSS活性比SN高75.58%。
T1-T5期间,QH和SN初级根茎GBSS活性无显著变化,并且其活性水平相对较低(图9 C),但T4和T5时期,QH初级根茎GBSS活性显著低于SN(P<0.05)。此外,QH和SN初级根茎SP活性呈现逐渐降低的趋势(图9 C),在T5时期SP活性分别为63.09和57.40 μg·min-1·g-1,较T1时期分别降低了37.50%和34.74%。总体来看,T1-T5阶段,淀粉代谢相关酶活性变化趋势和蔗糖代谢相关酶活性相似,合成酶活性均有所升高,分解酶活性均有不同程度的降低。
由图10可知,初级根茎向上弯曲阶段不同部位间淀粉代谢酶活性同样差异显著(P<0.05)。其中,AGPase、SSS和GBSS活性,均表现为P1节间活性高,P4节间活性低(图10 A,B,C)。P1节间,QH初级根茎AGPase、SSS和GBSS的活性分别为125.57,27.22和4.67 μg·min-1·g-1,P4节间较P1阶段分别降低了52.09%、78.78%和74.18%;SN初级根茎AGPase、SSS和GBSS活性分别为为114.69,24.50和5.61 μg·min-1·g-1,P4节间分别较P1阶段降低了61.45%,75.54%和74.13%。此外,初级根茎弯曲阶段各个部位QH和SN根茎AGPase、SSS和GBSS活性无显著差异。

图10 初级根茎向上弯曲阶段淀粉代谢相关酶活性变化Fig.10 Changes of starch metabolizing-related enzyme activities in different sections of primary rhizome during its bending
QH和SN初级根茎弯曲阶段不同部位SP活性分布与合成酶活性变化相反,P1节间活性最低,P4节间活性最高(图10 D)。P4节间QH和SN初级根茎SP活性分别为82.81和71.88 μg·min-1·g-1,较P1增加了54.64%和52.51%。同时,初级根茎弯曲阶段各部位间,QH和SN初级根茎SP活性无显著差异。
3 讨论
3.1 初级根茎不同发育时期/部位碳水化合物含量变化
根茎尚未突破土壤形成新的植物前,主要借助于母株提供的营养物质,或者说存贮于根茎中的营养物质[22],蔗糖在此过程中扮演重要角色[17,24]。当地上部分运输到根茎的蔗糖超过当时发育要求时,蔗糖就会在一系列酶的作用下形成淀粉,作为贮备物质;而当地上部分运输的蔗糖满足不了生长需求时,存贮的淀粉将重新被分解蔗糖,从而为植物发育提供能量[7]。有研究发现随着发育时间增加,鸢尾(Iristectorum)根茎中的淀粉含量同样和其生长时间成正比[12]。这些研究表明蔗糖和淀粉是根茎类植物储存物质的主要形式。本研究发现,初级根茎中蔗糖和淀粉的含量在发育过程中显著增加(P<0.05),尤其是蔗糖含量的积累更快,其积累量高于淀粉。这个研究结果既印证了其他学者的研究,又表明草地早熟禾初级根茎发育过程中营养储存方式兼顾蔗糖和淀粉两种形式,并且以蔗糖为主。此外,在初级根茎弯曲阶段,初级根茎基部节间蔗糖和淀粉含量显著高于弯曲部位(P<0.05),而葡萄糖和果糖含量显著低于弯曲部位(P<0.05)。初级根茎弯曲阶段碳水化合物的变化趋势和已有研究相似[13],水稻根茎基部蔗糖和淀粉含量同样显著高于根茎尖端(P<0.05)。同时,本研究发现,强根茎材料QH初级根茎蔗糖和淀粉含量显著高于弱根茎材料SN,这可能是两者初级根茎扩展或发育能力存在差异的因素之一。可见,在初级根茎的发育过程中,物质积累与自身生长之间存在一定的取舍。根茎发育一定阶段后,碳水化合物以蔗糖和淀粉的形式暂时储存在初级根茎基部,而初级根茎顶端的大部分蔗糖和淀粉分解成单糖,为初级根茎的扩张提供能量。这也证实了之前的研究,几乎所有的蔗糖和淀粉,在旺盛的生长部位,都被分解成葡萄糖和果糖,以支持植物的快速生长[7,24]。
3.2 初级根茎不同发育时期/部位蔗糖代谢酶活性变化
SPS、SS和INV是蔗糖代谢通路中的几个主要酶[25],SPS活性的大小决定了蔗糖合成的快慢,在蔗糖代谢中发挥了巨大的作用[26]。SS既可以促进蔗糖的合成,又可以促进蔗糖的分解[27]。INV包括SAI和NI,它们促进蔗糖分解为葡萄糖和果糖,从而促进植物生长发育,但通常SAI的活性远高于NI[28]。研究发现,Poapratensis根茎中的SPS活性与蔗糖含量呈显著正相关,而分解酶与蔗糖含量呈显著负相关[4]。此外,大豆(Glycinemax)果实成果过程中,SS和SPS活性同样与蔗糖含量呈显著正相关,而SAI活性与蔗糖含量呈显著负相关[28]。在本研究中,在初级根茎的发育过程中,蔗糖含量逐渐升高,QH初级根茎SPS活性先升高后降低,SN初级根茎SPS活性先降低后升高,并且蔗糖分解酶活性整体虽然均有所降低,但其降低幅度较小。这可能是由于初级根茎虽然是一个整体,但在发育过程中从初级根茎基部到顶端处于不同的活跃状态,因此导致蔗糖含量变化和酶活性变化整体规律性不强。但是SPS、SS活性和蔗糖含量在初级根茎的基部到顶端的分布呈现逐渐降低的趋势,而SAI和NI活性逐渐增加,这也再次表明初级根茎基部是蔗糖积累的主要部位,而初级根茎尖端的蔗糖在大量蔗糖分解酶的作用下被分解,从而促使初级根茎尖端生长发育。这与马铃薯、莲藕等的块茎植物的研究结果相似,其块茎未膨大前和根茎的尖端一样,大量蔗糖在分解酶的催化下被分解,从而促进其生长发育;而当块茎膨大时,则和根茎基部一样,在合成酶作用下迅速积累大量的蔗糖[29]。上述结果不仅证实了之前的观点,也提出了一些分歧。这也说明Poapratensis初级根茎发育是一个复杂的过程,初级根茎不同部位或不同发育阶段的碳水化合物代谢存在显著差异。
刘溢健等[30]认为INV参与细胞扩增过程,更高的INV活性可以促进植物快速生长。因此,通常在快速生长的组织中可以检测到更高的SAI活性,例如芽和根尖[31]。在本研究中,初级根茎尖端弯曲部分(P4)SAI活性和NI活性均高于其他部位,这一结果也证实了植物活跃部位SAI活性更高的观点。此外,无论那个发育时期或部位,NI的活性也远低于SAI活性,同样印证了前人的观点,在新梢和快速伸长的节间NI活性均远低于SAI[7]。
3.3 初级根茎不同发育时期/部位淀粉代谢酶活性变化
淀粉的生物合成和降解是一个高度调控的代谢过程,需要多种酶的协同作用。其中AGPase是淀粉生物合成中的关键酶,其活性高低决定了淀粉合成底物是否充足[32]。GBSS和SSS是主要的淀粉合成酶,其中SSS参与支链淀粉的合成,而GBSS参与直链淀粉的合成[33]。SP可以催化淀粉的分解或合成,但由于SP分解方向的底物浓度比合成方向的底物浓度高出近两个数量级,因此认为淀粉磷酸化酶只催化淀粉的分解[34]。在莲藕膨大过程中,AGPase和SSS的活性急剧增加,并且它们均与总淀粉含量呈极显著正相关[29]。此外,马铃薯块茎中的GBSS酶活性水平显著高于叶子[34]。在本研究中,从初级根茎基部到根茎弯曲部位,AGPase、SSS、GBSS活性和淀粉含量的分布呈现逐渐降低的趋势,而SP活性逐渐增加,这也再次表明初级根茎基部是淀粉积累的主要部位,而初级根茎尖端的淀粉在SP的作用下被分解,从而促使初级根茎尖端生长发育。此外,AGPase和SSS活性在初级根茎发育过程中同样呈现先升高后降低或持续升高的趋势,初级根茎淀粉酶活性变化的结果说明Poapratensis根茎发育是一个复杂的动态变化,淀粉酶活性变化与以往研究结果不同的原因可能是莲藕和马铃薯块茎的主要功能是储存淀粉供食用,QH和SN初级根茎中储存的淀粉主要是为其自身的无性繁殖做准备,在初级根茎发育过程中根茎尖端或根茎芽中的蔗糖和淀粉均被分解,以促进初级根茎扩张或新植物的形成。
4 结论
草地早熟禾根茎整体发育是一个十分复杂的过程,既是物质积累的过程,又是物质消耗的过程。其中,初级根茎发育的过程也是淀粉和蔗糖积累的过程,并且在越靠近根茎基部的节间,两者含量越高。此外,与弱根茎材料SN相比,强根茎材料QH蔗糖和淀粉的积累更为迅速。
