潜在抗肝纤维化药物与靶点相关信号通路研究进展
2023-12-18何雪茹付裕豪荀雪姣董占军
周 鑫, 王 智, 何雪茹, 付裕豪, 荀雪姣, 李 颖, 董占军
1 河北医科大学研究生学院, 石家庄 050017; 2 河北省人民医院药学部, 石家庄 050057
肝纤维化是由非酒精性脂肪性肝炎、病毒性肝炎、过度饮酒和金属中毒等引起的慢性肝损伤的伤口愈合的过程。不加干预的肝纤维化将会进展为肝硬化、肝癌,最终导致死亡[1]。全球约75%的肝癌发生在亚洲,仅我国就承担了全球50%的肝癌负担[2]。大量研究[3]表明,肝纤维化是可控的,甚至是可逆的。因此,及早地干预肝纤维化将极大地减轻我国的社会负担和医疗压力,延缓、逆转肝纤维化已成为治疗慢性肝炎的关键选择。细胞外基质(ECM)在肝脏内的过度沉积是肝纤维化的一个标志特征,作为其主要贡献者的肝星状细胞(HSC)在肝纤维化进行性发展中有非常重要的作用。肝纤维化是一个动态的过程,HSC 的活化与其他细胞之间的作用受多种信号通路之间复杂的相互作用影响[4]。同时,细胞的自噬与凋亡在HSC 的活化中也有多方面的影响。本文介绍了多条信号通路在肝纤维化进程中的影响并综述了非编码RNA 在肝纤维化进程中的不同作用,以期发现抗肝纤维化相关潜在候选目标与新的治疗靶点,为延缓、逆转肝纤维化提供新的思路。
1 肝纤维化相关信号通路
1.1 TGF-β介导的信号通路 TGF-β是一种多功能细胞因子,调节巨噬细胞、未成熟造血细胞和树突状细胞等多种细胞的增殖、分化、凋亡、黏附和迁移。TGF-β通过经典(基于SMAD)和非经典(非基于SMAD)信号通路表达信号响应。TGF-β激活下游介质,进一步激活几种细胞内信号通路以调节各种细胞功能。
1.1.1 TGF-β1/SMAD 信号通路与相关活性成分及药物 TGF-β1 通过激活下游底物(包括SMAD2 和SMAD3)发挥其生物学作用,同时受到SMAD7表达的负反馈调节[5]。在肝纤维状态下,SMAD2和SMAD3表达增加起正向调节作用,而SMAD7表达下降进行负调节。TGF-β/SMAD的信号传导起始于一个异源传导复合物,当TGF-β1与TGF-β受体Ⅱ(TGFβRⅡ)相互作用时,异源传导复合物被激活,诱导TGF-βRⅡ对TGFβRⅠ进行磷酸化,接着磷酸化TGFβRⅠ反过来磷酸化细胞质介质中的SMAD2/SMAD3,并且与SMAD4 形成异源三聚体复合物,逐渐向细胞核内转移,识别结合序列发挥转录因子作用[6]。
柚木具有抗炎保肝解热镇痛作用。柚木叶取物以剂量依赖性降低Ⅰ型α1 胶原链(COL1A1)、α-平滑肌肌动蛋白(α-SMA)、血小板衍生生长因子和组织金属蛋白酶抑制剂1(TIMP1)的基因表达,升高基质金属蛋白酶3(MMP3)的表达。同时柚木提取物在mRNA水平上呈剂量和时间依赖性,在蛋白水平上呈剂量依赖性,显著下调TGF-β1 和SMAD2 表达,上调SMAD7表达,通过调节TGF-β1/SMAD2 通路和上调MMP3/TIMP1 比值逆转肝纤维化[7]。芝麻酚是一种从芝麻中分离出的脂溶性木酚素类化合物,被证明对硫代乙酰胺(TAA)诱导的肝纤维化大鼠具有保护作用。芝麻酚以剂量依赖性方式显著降低TGF-β1、SMAD3、磷酸化的SMAD3(p-SMAD3)和α-SMA mRNA的表达和蛋白质浓度,上调SMAD7 mRNA 的表达,同时以浓度依赖性方式显著降低溶血磷脂酸受体1(LPAR1),LPAR3和结缔组织生长因子mRNA 的表达,通过抑制TGF-β1/SMAD3 信号通路和下调LPAR1/3 的表达发挥抗肝纤维化作用[8]。阿司匹林在体内外以剂量依赖性方式下调α-SMA、Ⅰ型胶原蛋白、TGF-β1、p-SMAD2/3 的蛋白质表达水平和α-SMA、COL1A1、TGF-β1 mRNA 水平,通过抑制TGF-β1/SMAD 信号减轻肝纤维化[9]。柠檬素通过上调SMAD7,减少SMAD2/3 的c 端磷酸化(p-SMAD2/3c)和核转位,抑制TGF-β/SMAD 信号通路来减轻肝细胞EMT 和HSC 活化,从而抑制四氯化碳(CCl4)诱导的小鼠肝纤维化[10]。
1.1.2 TGF-β1/其他信号通路与相关活性成分及药物 除规范的TGF-β1/SMAD 信号通路外,还存在由其他信号通路介导的TGF-β信号传导。例如,丝裂原活化蛋白激酶(MAPK)通路,包括细胞外信号调节激酶、c-Jun氨基末端激酶(JNK)、p38 MAPK,以及NF-κB抑制剂(IκB)激酶(IKK)、磷脂酰肌醇-3激酶(PI3K)、蛋白激酶B(Akt)和Rho 家 族GTPases,这 些 受 体 激 活 的 非SMAD 转导器以独立或与SMAD 联合的方式介导信号反应,并汇聚到SMAD上以控制SMAD活动[11]。
咖啡因阻断了磷酸化的ERK、JNK、p38 和pSMAD3L 蛋白水平的显著增加,通过阻断MAPK 和TGF-β/SMAD3 信号通路,减轻实验性非酒精性脂肪性肝炎和TAA 诱导的肝纤维化进展[12]。米糠油(RBO)富含多种生物活性植物化学物质,具有降低胆固醇、抗炎抗氧化活性等作用。RBO 显著下调NF-κβ 信号通路,进而抑制一些炎症介质的表达,包括环氧化酶2、IL-1β和TNF-α。RBO通过抑制TGF-β1、α-SMA、Ⅰ型胶原、羟脯氨酸、结缔组织生长因子和黏着斑激酶(FAK)的生物学作用,通过抑制HSC 活化和调节TGF-β1 和FAK信号转导之间的相互作用来减少肝纤维化[13]。五味子酯甲具有抗氧化抗炎抗细胞凋亡和心肌保护等多种药理活性,降低了磷酸化的TGF-β-活化激酶1(p-TAK1)/TAK1、p-JNK/JNK、p-p38/p38、p-IκB/IκB 和NF-κB p65 蛋白表达水平,不同程度上抑制了α-SMA 和COL1A1 mRNA的表达,通过TGF-β1介导的TAK1/MAPK和NF-κB通路的激活,在体内外改善肝纤维化[14]。
1.2 Wnt/β-catenin 信号通路与相关活性成分及药物Wnt 信号通路是一种比较保守的信号转导通路,在调节细胞功能和生长发育方面发挥重要作用。Wnt蛋白首先与靶细胞膜上的卷曲蛋白和辅助受体低密度脂蛋白受体相关蛋白5/6结合,从而通过蓬乱蛋白的磷酸化将胞外信号传递到细胞质内。然后细胞质内的蓬松蛋白通过抑制糖原合成酶激酶-3β(GSK-3β)的激活,阻止β-catenin 的磷酸化或降解,导致细胞质内游离的β-catenin增加。当细胞质中游离的β-catenin达到一定水平时,进入细胞核与核内淋巴细胞增强因子/转录T淋巴细胞因子(LEF/TCF)结合,形成β-catenin-LEF/TCF复合体,导致经典Wnt信号通路下游靶基因转录[15]。
漆黄素显著抑制与β-catenin 下降和GSK-3β 水平升高相关的wnt3a基因表达。此外,漆黄素还能减缓肝组织纤维化的进程,减少α-SMA和特异性周期蛋白-D1(cyclin D1)的肝脏表达,通过抑制Wnt/β-catenin 信号通路减轻TAA 诱导的大鼠肝纤维化[16]。和厚朴酚(HNK)是从广玉兰中提取的一种生物活性化合物,具有潜在的抗肝纤维化作用。HNK 激活了CC-3 和GSK3β,下调了HSC 中的α-SMA、Wnt3a 和β-catenin 的表达,通过激活GSK3β 和抑制Wnt3a/β-catenin 信号通路诱导细胞凋亡和抑制肝纤维化[17]。
1.3 PI3K/Akt/哺乳动物雷帕霉素靶蛋白(mTOR)信号通路与相关活性成分及药物 PI3K/Akt/mTOR 信号通路是癌症的关键通路之一[18]。大量的研究表明PI3K/Akt/mTOR 信号通路在肝纤维化中发挥着重要作用。不同的生长因子对应不同的受体酪氨酸激酶(RTK)。首先生长因子介导的RTK 刺激PI3K 的活化。活化的PI3K 使磷脂酰肌醇4,5-二磷酸(PIP2)磷酸化并将其转化为磷脂酰肌醇3,4,5-三磷酸(PIP3)。第二个信使PIP3 是Akt 的对接位点,而Akt 是PI3K 通路的核心介质。随后磷脂酰肌醇依赖性激酶14 与Akt之间的相互作用,导致Akt在thr308位点上进行磷酸化[19]。磷酸化的Akt通过结节性硬化症1/2 复合体激活下游效应蛋白mTOR,并上调各种转录因子,从而发挥其生物学活性作用。
多沙唑嗪是一种选择性α1 肾上腺素能受体阻断剂,被发现有抗肝纤维化作用。多沙唑嗪在体内外以浓度依赖性方式抑制HSC 的活化,上调PI3K/Akt/mTOR 信号通路中的p-mTOR 的表达从而抑制自噬来减弱肝纤维化[20]。富马酸替诺福韦二吡呋酯主要用于慢性乙型肝炎患者的抗病毒治疗,通过持续显著降低PI3K/Akt/mTOR 信号通路的磷酸化在TAA 诱导的肝纤维化模型中诱导HSC凋亡[21]。委陵菜酸是从委陵菜中提取的一种活性成分,委陵菜酸显著降低了α-SMA、TGF-β、半胱天冬酶-3、Bax、IL-6 和TNF-α 的表达,升高了Bcl-2 和IL-10 的表达,通过抑制甘油磷脂代谢途径以及PI3K/Akt/mTOR 和NF-κB 信号通路,缓解体内肝纤维化进展[22]。
1.4 NF-κB信号通路与相关活性成分及药物 NF-κB信号通路是一种高度保守的进化途径,在调节免疫和炎症反应方面具有相当重要的作用。PI3K/Akt 可调控NF-κB 通路/转录因子家族,而NF-κB 通路/转录因子家族可调控炎症、细胞应激、先天和适应性免疫反应,进而调控肝细胞、肝巨噬细胞和HSC 的生存、增殖、迁移和侵袭[23]。
NF-κB 包括RelA(p65)、RelB、c-Rel、p105/p50 和p100/p52五个家族成员。Rel蛋白作为成熟蛋白合成,p50 和p52 分别由前体p105 和p100 的蛋白体降解产生。这些蛋白具有保守的Rel 同源结构域,可与目标DNA 序列结合,形成homo/异二聚体,在没有任何刺激的情况下,通过IκB 定位于细胞质。NF-κB 的活化通过不同的典型和非典型途径都可以进行,在规范途径中,抑制剂IκB被IKK复合物磷酸化,并受到泛素介导的蛋白酶体降解。游离的NF-κB 二聚体(主要是p50/p65 和p50/c-Rel)进入细胞核内、并激活靶基因转录。在非典型途径中,NF-κB 诱导激酶被激活,随后p100被翻译后加工为p52 亚基,与RelB 二聚,以及核定位,从而诱导基因转录[24]。
二氢杨梅素是腾茶提取物中的一种天然活性物质,二氢杨梅素抑制了促凋亡蛋白Bax、半胱天冬酶-3/9(CC-3/9)、磷酸化蛋白及其上游炎症调节因子p-IKKα/β和p-IκBα、TGF-β1 和α-SMA 的表达,促进抗凋亡蛋白Bcl-2 和Bcl-XL 的表达。通过抑制NF-κB 介导的炎症和TGF-β1 调节PI3K/Akt 信号通路来逆转TAA 诱导的肝纤维化[25]。甲氧基丁香酚(4-烯丙基-2,6-二甲氧基苯酚)是存在于人类饮食中的成分,被广泛用作食品工业中的添加剂,显著降低了Col-1、α-SMA、IL-6、TNF-α、一氧化氮合成酶和IL-8 的表达,通过NF-κB/过氧化物酶体增殖物激活受体-ɣ和NF-κB机制减弱肝纤维化和炎症[26]。獐牙菜被用来治疗包括肝脏疾病在内的各种疾病,獐牙菜提取物显著抑制CCl4诱导的大鼠肝纤维化模型中TGF-β、α-SMA、SMAD2/3、Col1A1、NF-κB、TNF-α和IL-1β mRNA和蛋白的表达,恢复了肝组织中改变的铁调素水平,通过抑制TGF-β/SMAD 和NF-κB信号通路减轻大鼠肝纤维化并恢复肝铁调素水平[27]。西他列汀以其抗糖尿病活性而闻名,提高了Nrf2 及其靶基因的mRNA 表达,显著降低了NF-κB 的水平和蛋白表达以及TNF-α 的蛋白表达,通过激活Nrf2和抑制NF-κB通路发挥抗肝纤维化作用[28]。
1.5 Hippo 信号通路与相关活性成分及药物 Hippo途径是一种进化保守的信号通路,在伤口愈合和免疫调节中发挥关键作用。河马通路由一个信号网络组成,这些信号最终指示转录调节因子YES 关联蛋白(YAP)和PDZ结合基序转录共激活因子(TAZ)的功能。核YAP/TAZ 直接进行基因表达,调控细胞增殖和存活等一系列行为[29]。YAP/TAZ的消耗会阻断TGF-β诱导的肌成纤维细胞转化和ECM 的产生,而活化的YAP/TAZ 的异位表达会促进这些情况。YAP 与TAZ 结合直接调节TGF-β 激活的SMAD 转录因子,为促纤维化信号传导串扰提供了一种机制。
白藜芦醇是一种存在于葡萄花生等作物中的生物活性成分,已被证明通过多种信号通路发挥抗肝纤维化作用。白藜芦醇显著降低了YAP和TAZ的mRNA和蛋白水平,增强Bax 蛋白表达并降低Bcl-2 蛋白表达,部分通过Hippo通路抑制HSC活化[30]。N-乙酰-L-色氨酸(L-NAT)是N-乙酰色氨酸(NAT)的三种异构体之一,以往研究发现其具有神经保护抗炎保肝等作用。L-NAT 显著降低α-SMA、Ⅰ胶原、TGF-β1、p-SMAD2/3、MMP9 和Bax 蛋白水平的表达。增加Bcl-2 和YAP1 的蛋白表达,通过调控TGF-β1/SMAD 和Hippo/YAP1 信号抑制CCl4诱导的肝纤维化[31]。
1.6 MAPK 信号通路与相关活性成分及药物MAPK 途径是最保守的信号转导途径之一,在细胞增殖、分化、死亡和胚胎发生中具有关键功能。MAPK信号传导由三种基于激酶的反应组成,通过这些反应,由丝裂原激活的蛋白激酶激酶(MAPKK 或MEK)催化的上游激酶反应激活,而上游激酶反应又被另一种称为MAPKKK(MAPKK 激酶)或Raf(以快速加速纤维肉瘤命名)的上游激酶磷酸化[32]。常规MAPK包括JNK、p38 MAPK 和ERK。已有研究表明MAPK 通路与肝纤维化之间有着密切关系。
香芹酚是一种单萜烯酚类的非选择性TRPM7抑制剂,具有抗肿瘤、抗炎和抗肝纤维化作用,香芹酚显著降低了CCl4诱导肝纤维化小鼠模型中α-SMA、Col1α1、p-ERK1/2、p-JNK1/2和p-p38的表达,通过阻断MAPK通路发挥抗肝纤维化作用[33]。HNK是从广玉兰中提取的一种生物活性化合物,具有潜在的抗肝纤维化作用。HNK降低了α-SMA、COL1A1、COL3A1和SMAD2/3的表达,增加了微管相关蛋白1轻链3-Ⅱ(LC3-Ⅱ)和p62的表达,通过抑制TGF-β1/SMAD 信号通路和激活p38 MAPK通路分别介导HSC的活化与自噬,起到抗肝纤维化作用[34]。所有肝纤维化信号通路与潜在抗肝纤维化药物及活性成分详见表1。
2 非编码RNA(ncRNA)介导的肝纤维化信号通路
人类基因组中约80%的基因被主动转录为各种ncRNA,在多种疾病发病机制的重要途径中起关键作用。ncRNA 包括长链RNA(lncRNA)、微小RNA(miRNA)、环状RNA(circRNA)、小核仁氨酸(snoRNA)和Piwi相互作用的RNA(piRNA)。大量研究[35]表明,ncRNA 可以通过调节多个下游基因靶点和相关途径的表达能力,调控肝纤维化进程(表2)。
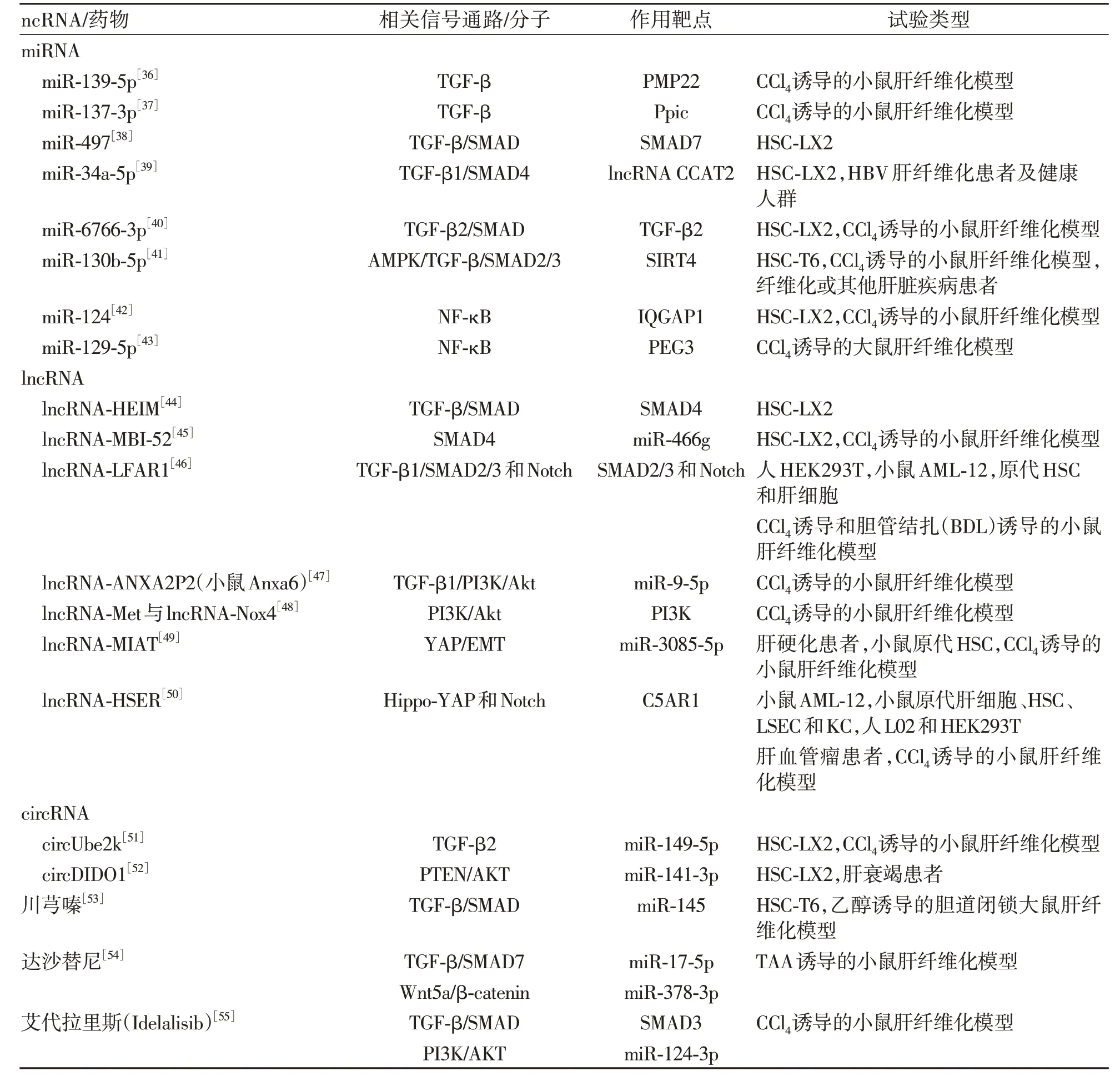
表2 ncRNA及药物与肝纤维化相关通路与试验类型总结Table 2 Summary of non-coding RNAs and drug and liver fibrosis-related pathways and trial types
2.1 miRNA 介导的肝纤维化信号通路及其靶点miRNA 是长度约22 个核苷酸的内源性ncRNA,在转录后水平调节基因表达。外周髓鞘蛋白22(PMP22)首先从小鼠NIH3T3 成纤维细胞中分离出来,主要表达在周围神经的髓鞘化雪旺细胞中,研究[36]表明PMP22 在CCl4诱导的小鼠肝纤维化模型和TGF-β 激活的HSC 中显著上调,miR-139-5p 通过靶向抑制PMP22 阻碍TGF-β 诱导的HSC 的激活。肽基脯氨酰异构酶C(Ppic)在骨髓中的造血祖细胞和胸腺中的T淋巴细胞前体细胞或胸腺细胞中表达,以往研究表明Ppic 在环孢素(CsA)诱导的肾损伤起重要作用。在CCl4诱导的小鼠肝纤维化模型中,Ppic 敲低降低了TGF-β 诱导的小鼠HSC 活化,miR-137-3p 直接靶向并负调节Ppic 的表达调控TGF-β 引起的小鼠HSC 活化[37]。miR-497 在多种疾病过程中起到一定作用,是潜在的生物标志物与治疗靶点。miR-497 可以通过靶向SMAD7 增强TGF-β/SMAD 信号通路以促进肝纤维化[38]。miR-34a-5p 靶向并负调节lncRNA CCAT2 的表达,通过TGF-β1/SMAD4 信号通路发挥抗肝纤维化作用[39]。三维人胚胎干细胞外泌体(3D-hESC-外泌体)中含有非编码小RNA与蛋白质等其他生物分子,已被证明可以增强组织再生并调节内源性修复。3D-hESC-外泌体中的miR-6766-3p 通过抑制TGFβRII 表达来灭活SMAD 信号传导,从而减弱HSC 活化并抑制肝纤维化[40]。miR-130b-5p 的表达在多种肝损伤模型中上调,miR-130b-5p通过AMPK/TGF-β/SMAD2/3信号通路调节SIRT4,从而促进肝纤维化[41]。含有IQ 基序的GTP 酶激活蛋白1(IQGAP1)是一种信号支架蛋白,通过促进细胞中的信号转导来调节一系列细胞活动。研究[42]发现miR-124的表达在肝纤维化组织和TNF-α诱导的LX-2 细胞中下调,同时伴随着IQGAP1 的上调表达,miR-124 的过表达可通过靶向IQGAP1 抑制NF-κB途径来降低HSC 炎性细胞因子的分泌。父系表达基因3(PEG3)是一种编码DNA结合蛋白的印记基因,是一种已知的转录抑制因子。miR-129-5p 可能调节纤维腺炎相关基因的表达,miR-129-5p 在CCl4诱导的大鼠模型肝纤维化组织中表达较低,PEG3 表达较高。研 究[43]表明miR-129-5p 可以通过降 低PEG3 抑制NF-κB信号通路发挥抗肝纤维化作用。
2.2 lncRNA 介导的肝纤维化信号通路及其靶点lncRNA 是一类长度大于200 个核苷酸且不具有蛋白质编码能力的ncRNA。lncRNA-ENST00000519726(HEIM)是一种新鉴定的lncRNA。lncRNA-HEIM 的表达在HBV 感染后上调,导致TGF-β 和SMAD4 的表达增加,并进一步促进HSC 的纤维化[44]。lnc-MBI-52在体内和体外肝纤维化过程中均增加,lnc-MBI-52 可以通过抑制miR-466g表达来抑制SMAD4 表达发挥抗肝纤维化作用[45]。lnc-LFAR1 在HSC 中特异性上调,通过激活TGFβR1/SMAD2/3 和Notch 信号通路促进肝纤维化[46]。小鼠Anxa6 敲低部分在TGF-β1 刺激的HSC 中 抑 制 了TGF-β1 诱 导 的HSC 活 化 并 阻 断 了PI3K/Akt 信号激活,小鼠Anxa6/miR-9-5p/Anxa2 轴和PI3K/Akt 通路可能参与了lncRNA ANXA2P2(mouse Anxa6)对 肝 纤 维 化 的 作 用[47]。 lncRNA-Met 与lncRNA-Nox4是两种新型lncRNA,共表达可以通过氧化还原过程和PI3K/Akt信号通路激活HSC,增加ECM的产生,并调节肝纤维化的过程[48]。lncRNA-MIAT的表达与miR-3085-5p 水平在肝硬化患者和活化的HSC 中呈负相关,lncRNA-MIAT 至少部分通过miR-3085-5p/YAP/EMT 信 号 通 路 增 强HSC 的 活 化[49]。lncRNA-HSER 在肝细胞中特异性表达,通过C5AR1-Hippo-YAP 和Notch 信号通路分别抑制肝细胞凋亡和肝细胞上皮间质转化[50]。
2.3 circRNA 介导的肝纤维化信号通路及其靶点circRNA 是由前体mRNA 反向剪接产生的一类内源性ncRNA。circUbe2k 在CCl4诱导的肝纤维化小鼠和TGF-β1 刺激的LX-2 细胞中表达上调,抑制其表达可以通过miR-149-5p/TGF-β2 轴抑制HSC 的活化和增殖[51]。circDIDO1 通 过 靶 向miR-141-3p 上 调 升 高PTEN 蛋白水平,降低LX2 细胞中p-AKT/AKT 的比例抑制HSC的活化[52]。
2.4 ncRNA 相关药物与活性成分及其靶点 中药生物碱单体川芎嗪在体内和体外上调miR-145表达并抑制TGF-β/SMAD 信号通路发挥抗肝纤维化作用[53]。达沙替尼是第二代口服多靶点RTK抑制剂,上调miR-378-3p的表达抑制Wnt-10a,下调miR-17-5p升高WiF-1和SMAD-7 的表达,通过抑制Wnt/β-catenin 和TGF-β/SMAD 通路抑制TAA 诱导的小鼠肝纤维化[54]。艾代拉里斯(Idelalisib)是首个高选择性口服磷脂酰肌醇3-激酶δ(PI3K-δ)抑制剂,用于治疗复发性慢性淋巴细胞白血病、复发性滤泡性淋巴瘤、复发性小淋巴细胞淋巴瘤。艾代拉里斯显著降低了COL1和α-SMA 基因及蛋白水平的表达,通过抑制TGF-β/SMAD3信号通路的活化抑制了HSC的活化,同时艾代拉里斯上调miR-124-3p的表达抑制PI3K/AKT通路促进细胞凋亡[55]。
3 小结
肝纤维化的进展涉及HSC、肝巨噬细胞在内的多种细胞,通过多通路相互交联作用,调节细胞活化、细胞自噬和细胞凋亡等多种形式。上文中提到的潜在活性成分及靶标大多数处于前瞻性研究阶段。近年来,随着人们对肝纤维化认识的逐步加深,已经有越来越多的药物及活性成分发现具有抗肝纤维化的潜力,新的肝纤维化调节方式也在逐渐被发现。目前,药物治疗仍是肝纤维化的主要治疗方式。ACC 抑制剂Aramchol Meglumine、THR-β 激动剂Resmetirom、小分子半乳糖凝集素-3 抑制剂Belapectin 和GB1211、脂质纳米颗粒BMS-986263 等一批新药已步入Ⅱ/Ⅲ期临床研究,相信随着对以往机制及新发现机制的深入剖析,一定能给药物抗肝纤维化治疗提供新的助力。
利益冲突声明:本文不存在任何利益冲突。
作者贡献声明:周鑫负责文献检索,撰写论文;王智、何雪茹、付裕豪、荀雪姣负责资料分析;李颖、董占军负责指导撰写文章并最终定稿。
