补料策略优化促进乳酸乳球菌HB03发酵合成Nisin
2023-12-14熊华仪刘月锋
熊华仪,陈 曦,刘月锋,陈 雄,李 沛,王 志,*
(1.湖北工业大学 发酵工程教育部重点实验室,工业发酵省部共建协同创新中心,湖北 武汉 430068;2.安琪酵母股份有限公司,酵母功能湖北省重点实验室,湖北 宜昌 443003)
乳酸乳球菌同型乳酸发酵过程中合成的乳酸链球菌素(Nisin)由34 个氨基酸组成[1],其能有效抑制革兰氏阳性菌(G+)的生长,是一种安全、绿色的活性抗菌肽,被广泛应用于食品发酵领域[2]。运用基因工程改造菌株有效提高了Nisin合成效率,Papagianni等[3]将黑曲霉A60的磷酸果糖激酶基因pfk13、编码cAMP依赖型蛋白激酶pkaC和选择性氧化酶aox1基因克隆至乳酸乳球菌ATCC11454,提高了其糖酵解活性并促进了菌株的氧化呼吸能力,Nisin效价达到14 000 IU/mL。Hao Panlong等[4]发现过表达天冬氨酸合成酶基因(asnH)可以使乳酸乳球菌F44在pH 3条件下的存活率提高7 倍,同时Nisin产量提高57%,达到5 346 IU/mL。然而基因工程菌应用于食品发酵存在一定的安全风险,运用发酵工程策略提高Nisin合成效率仍然具有一定的积极意义。
Nisin合成效率与氨基酸前体或多肽的供给有关[5],并与乳酸乳球菌生长存在氮源竞争关系[6]。Nisin含有5 个脱水氨基酸残基丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和半胱氨酸(cysteine,Cys)合成的硫醚键[2],因而Cys的合理供应是满足高效Nisin发酵的基础条件之一。另外,乳酸乳球菌厌氧发酵过程依然存在氧化胁迫效应[7],Gaudu等[8]发现Cys作为还原剂能显著增加乳酸乳球菌的存活率并减少氧化胁迫损伤而促进细胞生长[7]。而有关Cys对Nisin发酵合成和氧化应激的影响鲜见报道。
乳酸乳球菌的蛋白水解体系[9]包括壁膜蛋白酶PrtP、多肽转运蛋白DtpP、胞内肽酶PepV[10]。该系统的表达受氮代谢全局调控子CodY负控制且主要受异亮氨酸的调节[11]。另外,Vido等[12]发现了不依赖于CodY的蛋白水解体系调控机制,乳酸乳球菌由发酵转为呼吸代谢可强烈诱导蛋白水解体系中性内肽酶(PepO1)和氨肽酶C(PepC)的表达。
精氨酸(arginine,Arg)在乳酸乳球菌生长代谢过程中也发挥了重要作用,Arg经脱亚氨酶(arginine deiminase,ADI)途径[13]分解,产生铵根和氨甲酰磷酸以及后续的分解产物CO2、铵根和ATP,这在细胞应对酸胁迫中发挥了重要作用[14]。乳酸乳球菌Arg代谢与嘧啶合成代谢也密切关联[15-16],调控蛋白ArgR阻遏arg操纵子表达,ΔargR菌株中Arg合成基因上调,同时尿嘧啶核苷酸(uridine monophosphate,UMP)从头生物合成基因(pyrRPB、carA、pyrEC及pyrKDbF)表达也上调[15]。由于肽聚糖合成始于尿苷三磷酸(uridine triphosphate,UTP)和N-乙酰葡萄糖胺-1-P[9],因而UTP的不足也可能导致细胞肽聚糖合成及生长受限[17],甚至引起细胞在胁迫条件下自溶[18],因此Arg在细胞能量供应、酸胁迫应激及UTP与肽聚糖合成方面发挥着重要作用,而有关Arg对乳酸乳球菌Nisin发酵合成效率的影响鲜见报道。
为了提高Nisin的合成效率,在10 L发酵罐水平研究碳氮源(糖、Cys、蛋白胨)的补料策略对Nisin合成的影响,分析Nisin合成关键时间节点前后转录组表达差异,并结合胞外氨基酸浓度变化规律,确定Arg为限制氨基酸,并优化补料策略,显著提高Nisin发酵合成效率。
1 材料与方法
1.1 材料与试剂
乳酸乳球菌乳酸亚种(Lactococcus lactissubsp.lactisHB03),本实验室保藏。
蛋白胨、酵母浸粉 安琪酵母股份有限公司;蔗糖(食用级)市售;K2HPO4·3H2O、NaCl、MgSO4·7H2O、MnSO4·H2O、CaCO3、H2SO4、HCl、蒽酮、柠檬酸铵(均为分析纯)国药集团化学试剂有限公司;Nisin标准品(1 000 IU/mg)美国Sigma公司。
斜面种子培养基:15 g/L胰蛋白胨,15 g/L酵母浸粉,15 g/L牛肉膏,10 g/L葡萄糖,10 g/L CH3COONa,2 g/L Na2HPO4,2 g/L柠檬酸铵,10 g/L CaCO3,20 g/L琼脂,pH 6.8。
摇瓶培养基:70 g/L蛋白胨,16 g/L蔗糖,10 g/L酵母浸粉,2 g/L K2HPO4,0.2 g/L MgSO4·7H2O,0.05 g/L MnSO4·H2O,5 g/L柠檬酸铵,6 g/L CaCO3。
10 L发酵罐培养基:70 g/L蛋白胨,16 g/L蔗糖,10 g/L酵母浸粉,2 g/L K2HPO4,0.2 g/L MgSO4·7H2O,0.05 g/L MnSO4·H2O,5 g/L柠檬酸铵。
1.2 仪器与设备
SPX-150D恒温培养箱 上海博讯实业有限公司医疗设备厂;HNV-211B摇床 天津市欧诺仪器仪表有限公司;YXQ-75SII立式蒸汽灭菌锅 上海博讯医疗生物仪器股份有限公司;Ultimate 3000高效液相色谱仪 美国Agilent公司;S-10生物传感器仪 西尔曼科技有限公司;C J-2 D 净化操作台 天津泰斯特仪器有限公司;10 L机械搅拌通风发酵罐 上海保兴生物设备有限公司;V-1300分光光度计 上海美析仪器有限公司。
1.3 方法
1.3.1 菌株活化
将-80 ℃冰箱保存的乳酸乳球菌HB03甘油管接种于斜面种子培养基的平板,于30 ℃培养箱24 h后转接茄子瓶,30 ℃培养24 h,无菌操作加入60 mL无菌水,并用无菌竹签将菌苔挑起混合于无菌水中,得到种子液,备用。
1.3.2 摇瓶培养及10 L发酵罐实验
摇瓶培养:装液量为50 mL/250 mL发酵培养基,调节初始pH 6.8~7.0,121 ℃灭菌20 min,每瓶按OD600nm=0.1接种。放置于30 ℃恒温摇床间歇振荡培养12 h(每小时工作30 s,转速200 r/min,随后调整转速为0 r/min)。
10 L罐分批发酵:10 L发酵罐中装量7 L发酵培养基,调整初始pH 6.8~7.0,121~123 ℃灭菌25 min;接种60 mL种子液,30 ℃、100 r/min不通气培养20~24 h。
1.3.3 分析检测
1.3.3.1 生物量(以细胞数计)测定
稀释涂布平板法[19]。
1.3.3.2 总糖测定
参考硫酸-蒽酮法[20]。
1.3.3.3 乳酸测定
生物传感仪[21]测定。
按式(1)计算:
1.3.3.5 Nisin效价测定
采用高效液相色谱法[22]测定。样品处理:发酵液用盐酸稀释至pH 2.0~2.7[23],12 000 r/min离心20 min,经0.45 μm滤膜过滤后备用。Nisin标准方程绘制:将0.25 g Nisin标准品称取至50 mL容量瓶,使用0.01 mol/L盐酸溶液(pH 2.0)定容,得到标准液(5 000 IU/mL),经0.45 μm滤膜过滤备用。色谱条件:流动相:水-乙腈(81∶19,V/V)混溶后加入0.05%三氟乙酸;流速1 mL/min;检测波长200 nm;柱温40 ℃;进样量20 μL。
1.3.3.6 H2O2浓度的测定
采用索莱宝公司试剂盒,按说明书方法检测H2O2浓度。
1.3.3.7 游离氨基酸的测定
参考文献[24]的方法进行测定。
1.3.4 转录组数据分析
取发酵15、18 h的发酵液20 mL于50 mL离心管中,4 ℃、12 000 r/min离心5 min,无菌操作留取菌体沉淀并-80 ℃保存。转录组测序由美吉生物科技有限公司完成。转录表达定量分析每百万读取次数中某个转录本占比(transcripts per million,TPM)值按照式(2)计算[25]:
式中:R、L为需计算基因的read counts和基因长度/kb;Ri、Li(i=1,2,…,n)为样品中第i个基因的read counts和基因长度/kb。
1.4 数据分析
2 结果与分析
2.1 摇瓶水平Cys添加促进Nisin合成
Nisin是由34 个氨基酸组成的多肽,含有5 个硫醚桥(由Cys、Thr、Ser生成)[2]。因而,Cys是Nisin的重要前体氨基酸。另外,其作为含硫氨基酸还具有抵御氧化胁迫的作用[8]。因此,摇瓶水平考察了对数中期(16 h左右)添加不同浓度Cys对菌株HB03合成Nisin的影响,如图1所示。16 h添加0.15~0.6 mmol/L的Cys在24 h的Nisin效价较对照(未添加Cys)分别提升7.9%~20%。显然,Cys的添加促进了Nisin的生成。Cys添加量为0.6 mmol/L时Nisin效价为6 457 IU/mL。但随着Cys的添加量增多Nisin效价呈下降趋势,当添加量为0.75 mmol/L和0.9 mmol/L时,Nisin效价分别为6 140 IU/mL和5 862 IU/mL,也较对照提高14.8%和9%。综上Cys添加量0.6 mmol/L为最优,提高效价幅度达到20%。
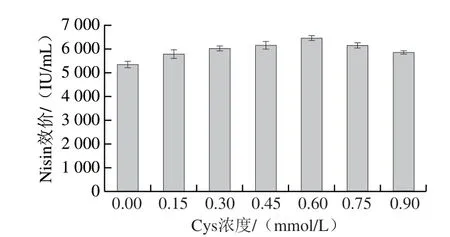
图1 摇瓶中Cys浓度对Nisin合成的影响Fig.1 Effect of Cys concentration on nsin production in shake flask culture
2.2 10 L发酵罐水平Cys添加促进Nisin合成
虽然发酵过程是静置(摇瓶)和未通空气的搅拌状态(10 L罐),但是发酵体系存在空气接触过程,乳酸乳球菌通过乳酸氧化酶LOX途径[26-27]、NADH氧化途径[28]等把氧气消耗,但同时产生了H2O2,而乳酸乳球菌是接触酶阴性菌株,因而细胞生长过程伴随着氧化胁迫发生[29-30]。由于Nisin合成与生长偶联[7],为了降低氧化胁迫效应以及维持细胞生长效率,在罐上将Cys的添加策略调整为对数期及其稳定期(11、13.5、18、23.5 h每次添加0.15 mmol/L,总计0.6 mmol/L)。10 L罐水平Cys添加策略(终浓度0.6 mmol/L,策略1#)对乳酸乳球菌HB03生长代谢的影响如图2所示。

图2 Cys补料策略1#对Nisin合成的影响Fig.2 Effect of Cys supplementation strategy 1# on nisin synthesis
细胞生长至18 h达到峰值(1.43×1010CFU/mL),而后自溶至21 h的1.16×1010CFU/mL并维持稳定。总糖由15.8 g/L缓慢消耗至14.62 g/L(9 h)后迅速消耗至1.75 g/L(21 h)。Nisin合成于15 h达到峰值(6 993 IU/mL)后迅速分解。同时,细胞对数早期(9 h)H2O2为0.87 mmol/L,这与文献[30-31]氧化胁迫伴随着细胞生长过程的报道一致。前9 h细胞生长启动较慢(糖耗也较低)可能也与细胞调整代谢以适应氧化胁迫过程有关。在细胞对数生长早期(11 h)添加Cys(0.15 mmol/L)使H2O2迅速降至0.18 mmol/L,在15 h降至0 mmol/L并维持至发酵结束。说明Cys少量多次添加方式对消除H2O2有效。
Cys消除H2O2使细胞生长加速,18 h细胞数为1.43×1010CFU/mL,平均生长速率为7.9×108CFU/(mL·h),是前9 h生长速率(3×108CFU/(mL·h))的2.6 倍。Nisin合成偶联细胞生长至15 h达到峰值(6 993 IU/mL),15~18 h细胞继续生长,但Nisin处于被降解的状态,也印证了Nisin合成与细胞生长对氮源存在竞争关系的报道[10]。此时,Nisin可能作为一种胞外多肽的储备分子,其被降解可能也是细胞维持活性的一种生存策略。由于15 h后H2O2浓度为0 mmol/L,因而Nisin的降解可能与其他未知因素有关,并调整Cys补加策略为:11 h和13.5 h分别加入0.15 mmol/L Cys。
2.3 Nisin对数后期降解前后转录组水平基因表达差异分析
1#策略发酵15~18 h期间,Nisin以221.6 IU/(mL·h)的速率迅速降解,但细胞数却提高了30%,达到了1.43×1010CFU/mL。即15~18 h是Nisin合成和细胞生长竞争的关键节点,因此,在转录组水平研究了15~18 h相关基因转录差异,包括肽酶相关基因、UMP从头合成途径基因、肽聚糖合成相关基因等,其显著差异基因如图3所示。
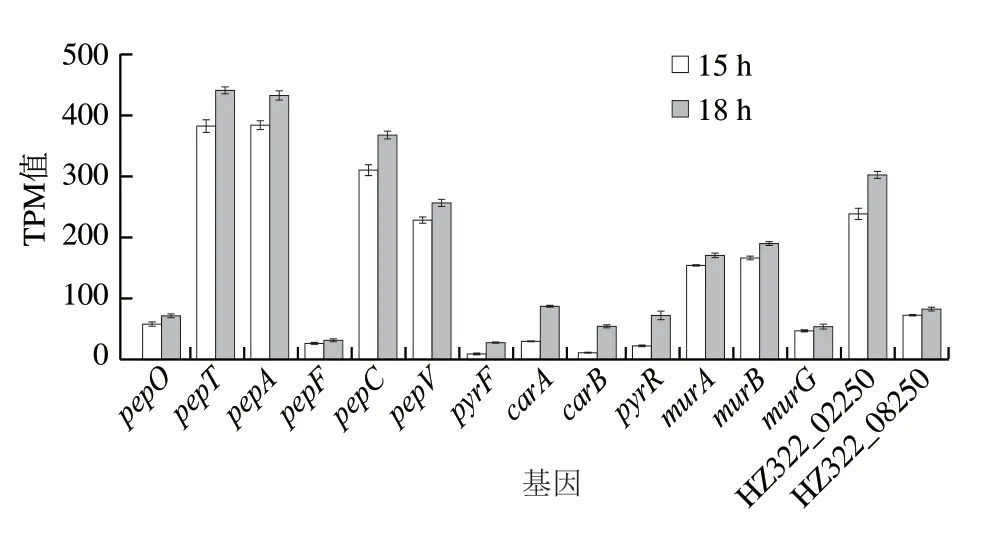
图3 15 h与18 h相关基因表达差异Fig.3 Differential expression of related genes at 15 and 18 h
pepO、pepT、pepA、pepF、pepC、pepV等胞内肽酶表达相关基因分别上调12%~24%,说明发酵15~18 h肽酶表达上升,强化了蛋白、多肽(含Nisin)的分解以用于生长。同时,18 h细胞UMP从头合成途径合成基因(pyrF、carA、carB、pyrR等)显著上调至2~4 倍,而18 h后细胞大量自溶,这可能与UMP合成(或合成前体)不足有关。而肽聚糖合成相关基因(murA、murB、murG等)以及与肽聚糖聚合的LysM蛋白相关基因(HZ322_08250、HZ322_02250)表达也均上调10%~27%。这与文献[15,32]报道一致。说明UMP合成前体氨甲酰磷酸可能处于限制状态,使得UMP、UTP供应不足,进而影响肽聚糖前体二磷酸尿苷葡萄糖的合成。细胞通过大幅上调UMP合成途径与肽聚糖合成相关基因以期满足细胞需求。
氨甲酰磷酸在Arg ADI途径中产生[13],说明18 h不仅氨甲酰磷酸可能缺乏,同时Arg也处于限制状态。因而,细胞为寻求Arg的及时补充,肽酶相关表达也上调12%~24%。这与含硫氨基酸或Arg的缺乏导致肽酶表达增强[9]的报道一致,细胞需要大量分解多肽以响应含硫氨基酸的缺乏,以及通过ADI途径降解Arg缓解UMP合成压力和应对酸胁迫压力。Nisin作为一种胞外的多肽“储备分子”,在细胞存在生存压力而加强肽酶表达时必然会被大量的降解。因此15~18 h细胞仍能持续生长,但Nisin效价下降。
2.4 基于转录组分析的Nisin补料策略优化
Nisin合成与细胞生长对氮源存在竞争关系[10]说明氮源的供应不足,因而,发酵过程中补加氮源(蛋白胨等)能缓解细胞对氮源的需求,也可作为前体直接用于Nisin合成;而Arg具有氮源和能源分子的双功能,也是需要考虑的因素。另外,蔗糖作为发酵过程唯一的碳源,适宜的流加也是必要的。补加碳氮源时间以及氨基酸对Nisin合成影响的设计如表1所示。

表1 10 L发酵罐上补料策略设计Table 1 Design of feeding strategy in a 10 L bioreactor
在补加Cys(11、13.5 h分别加入0.15 mmol/L Cys)的前提下,考虑到细胞自溶和Nisin降解同时发生,以及策略1# 9~18 h的平均糖耗速率为1.42 g/(L·h),确定策略2#为15 h补入蛋白胨(15 g/L)(1.5 g/(L·h)),其对乳酸乳球菌HB03代谢以及Nisin合成的影响如图4所示。
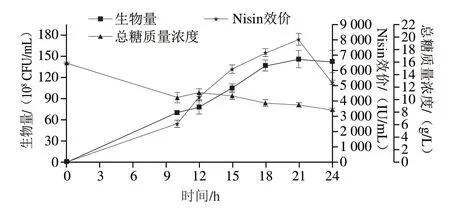
图4 策略2#对细胞生长代谢的影响Fig.4 Effect of strategy 2# on cell growth and nisin production in a 10 L bioreactor
Nisin 迅速合成至21 h 效价达到峰值,为8 042 IU/mL,较1#策略提高15%,后迅速分解。其_vq为383 IU/(mL·h)。10 h开始补糖以及15 h补加的蛋白胨使生物量在18~24 h稳定,总糖由15.8 g/L降至10.33 g/L(10 h),10~24 h残糖稳定维持在8~11 g/L之间。
以上结果说明,碳氮源的持续补加为乳酸乳球菌HB03生长以及Nisin合成起着积极作用,15 h补入蛋白胨使Nisin持续合成,但21 h Nisin合成达到峰值后依然以较快的速率降解。考虑到蛋白胨体系中可能存在氨基酸比例不平衡的问题,因此对策略2#细胞生长和Nisin主合成期(10~24 h)胞外氨基酸进行了测定,如表2所示。

表2 策略2#中胞外14 种氨基酸随时间的变化Table 2 Changes in concentrations of 14 extracellular amino acids in strategy 2#
Arg发酵过程均未检出,说明Arg是一种严重缺少的限制氨基酸。Arg参与ADI途径产能、缓解胞内pH值下降引起的酸胁迫、满足细胞生长、参与嘧啶合成[33]。因而,Arg的严重不足导致嘧啶途径表达的大量上调,与转录组基因差异结果一致,需要适量补充。
另外,胱氨酸发酵全过程质量分数稳定维持0.015%~0.025%,氧化条件下Cys会转化为胱氨酸,考虑添加的Cys可能不足,所以在策略2#的基础上17、19.5 h分别加入0.15 mmol/L Cys。12~15 h为614 IU/(mL·h),15~18 h为356 IU/(mL·h),从Nisin合成速率看12~15 h速率更快,所以将蛋白胨(15 g/L)补充时间提前至12 h,形成策略3#(11、13.5、17、19.5 h分别加入0.15 mmol/L Cys,10 h补蔗糖(1.5 g/(L·h))以及12 h补充蛋白胨(15 g/L)。其对乳酸乳球菌HB03代谢以及Nisin合成的影响如图5所示。
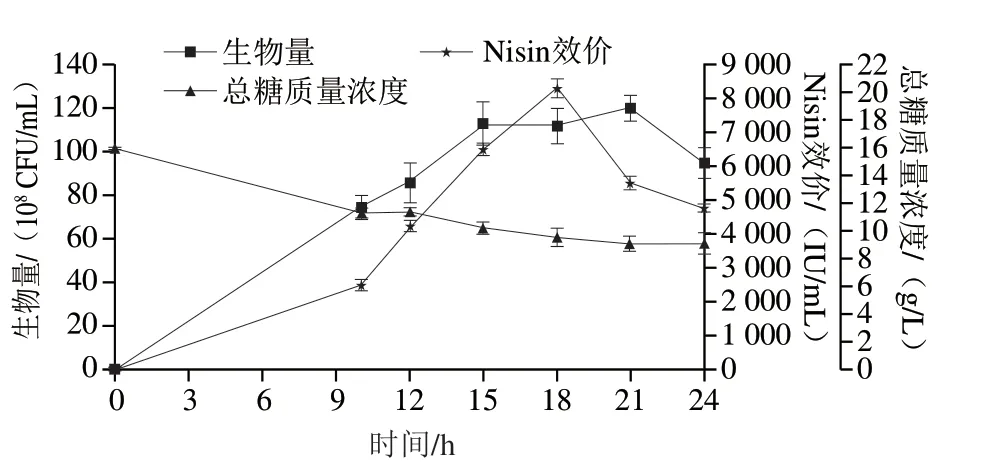
图5 策略3#对细胞生长代谢的影响Fig.5 Effect of strategy 3# on cell growth and nisin production in a 10 L bioreactor
细胞在10 h已经进入对数期,持续生长至15 h稳定后,于21 h生物量达到峰值,为1.20×1010CFU/mL,为1#策略的84%,后自溶。残糖由0 h的15.8 g/L迅速下降至10 h的11.32 g/L,10~24 h缓慢利用至9.18 g/L。Nisin合成在18 h达到峰值,为8 311 IU/mL,较1#策略提高18.8%,后迅速降解。其为461.7 IU/(mL·h),较2#策略提高20%。3#策略与2#策略相比,Nisin峰值效价差别较小,但达到Nisin效价峰值的时间点有所偏移,提前加入蛋白胨提前了Nisin的合成峰值时间。显然,加大含硫氨基酸的浓度以及提前蛋白胨的添加时间对于细胞来说虽然效价无明显提升,却提高了Nisin合成的速率。然而Nisin效价达到最高后仍然迅速分解,18~24 h Nisin以589.3 IU/(mL·h)的速率迅速降解。
细胞通过ADI途径获得能量、升高胞内pH值。ADI中间产物-氨甲酰磷酸可以作为前体进入嘧啶代谢,进而可能缓解细胞肽聚糖合成不足导致的细胞自溶以及肽酶大量表达等影响。因此需要确定Arg对Nisin合成的影响,如图6所示。

图6 摇瓶中Arg质量浓度对Nisin合成的影响Fig.6 Effect of different concentrations of Arg on nisin production in shake flask culture
摇瓶水平21 h添加1.5、1.8、2.5、3 g/L的Arg,在24 h较对照(未添加Arg)Nisin效价分别提升16%、23%、35%、29%。显然,添加Arg显著促进了Nisin的生成,以2.5 g/L最佳。在策略3#的基础上12~21 h加入Arg 0.25 g/(L·h),形成策略4#。其对乳酸乳球菌HB03代谢以及Nisin合成的影响如图7所示。
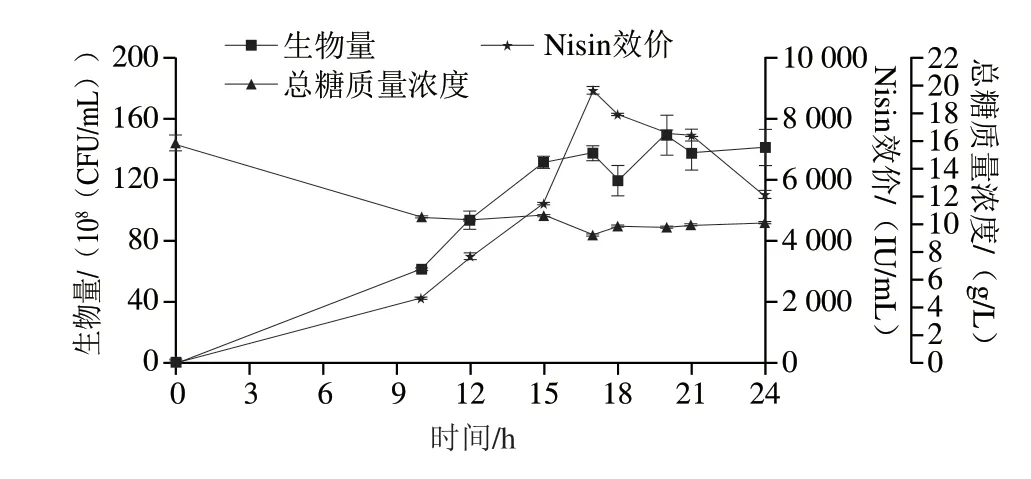
图7 策略4#对细胞生长代谢的影响Fig.7 Effect of strategy 4# on cell growth and nisin production in a 10 L bioreactor
细胞在10 h已经进入对数期,持续生长至15 h达到1.32×1010CFU/mL,后15~24 h维持稳定,其中生物量峰值为1.50×1010CFU/mL(20 h),较策略3#提高25%。残糖由0 h 15.8 g/L迅速下降至10 h 10.48 g/L,补糖后10~24 h保持在9~10 g/L之间。Nisin合成在17 h达到峰值,为8 963 IU/mL,较策略3#提高8%,17~21 h的Nisin以342.5 IU/(mL·h)速率降解,为策略3#的58%。而21 h后Arg不再流加,21~24 h Nisin降解速率恢复至686 IU/(mL·h),较17~21 h提高100%。说明Arg可大幅度延缓Nisin的降解速率,后续可增大添加量以持续促进Nisin合成和细胞生长。
3 结论
乳酸乳球菌HB03发酵合成Nisin过程存在氧化胁迫(9 h的H2O2为0.87 mmol/L),10 L罐发酵过程分4 次添加Cys(11、13.5、18、23.5 h分别加入0.15 mmol/L)降低了H2O2浓度,加速了细胞的生长,促进了Nisin合成,达到6 993 IU/mL。通过分析胞外氨基酸消长规律和肽水解酶系、UMP从头合成、肽聚糖合成代谢在转录组水平的表达差异,确定了对数中后期Nisin合成限制因素为碳氮源及Arg供应不足,并通过氨基酸单因素和碳氮源补加优化,确定了10 L发酵罐水平优化的补料策略。在此条件下Nisin效价达到8 963 IU/mL,比只补加Cys发酵效价(6 993 IU/mL)提高了28.2%,也比优化前效价(5 349 IU/mL,资料未显示)提高了67%。
