芝麻蛋白Ses i 3的重组表达及致敏性鉴定
2023-12-14姜松松汤鑫磊黄雨彤虞丹丹孟祥忍
姜松松,王 涛,汤鑫磊,李 倩,黄雨彤,虞丹丹,孟祥忍,2,*
(1.扬州大学旅游烹饪学院,江苏 扬州 225127;2.扬州大学中餐繁荣基地,江苏 扬州 225127)
芝麻过敏是食物过敏的一种,是机体摄入芝麻或芝麻制品引起的一种异常免疫反应。近年来,随着芝麻及其制品销量增多,芝麻过敏患者也逐渐增多,在世界范围内引起广泛关注。流行病学调查结果显示,芝麻是中东地区的主要食物过敏原之一,是以色列的第三大食物过敏原[1]。目前,芝麻已经被许多国家列入主要食物过敏原清单,如法国(芝麻过敏发病率为3%)[2]、澳大利亚(芝麻过敏发病率为0.8%)[3]和美国(芝麻过敏发病率为0.49%)[4]等,加拿大明确要求在食品包装上必须标注芝麻过敏原信息[5]。芝麻是我国常见的油料作物和加工食品原料,在我国食品和餐饮行业中广泛应用,消费量巨大。目前,我国虽然还没有准确的芝麻过敏流行病学调查报告,但一项对食物过敏患者过敏原调查结果显示,芝麻过敏发病率位列第三,仅次于小麦和鸡蛋,芝麻是7~17 岁年龄段食物过敏患者最主要的食物过敏原,且发病率逐年升高[6]。此外,研究发现过敏患者摄入极少量的芝麻即可诱发严重的过敏临床症状,如荨麻疹、皮肤水肿瘙痒、腹泻等,严重的甚至可能诱发过敏性休克,导致死亡[7]。2022年,综合芝麻过敏的发病率、临床症状严重程度和潜在危害考虑,联合国粮食及农业组织和世界卫生组织将其列入新“8大类”食物过敏原之一[8]。由此可见,芝麻过敏已经成为一个不容忽视的公共健康问题。
目前,已经被鉴定出来的芝麻过敏原蛋白有7 种,包括2S贮藏白蛋白(Ses i 1和Ses i 2)、7S类豌豆球蛋白(Ses i 3)、油质蛋白(Ses i 4和Ses i 5)和11S球蛋白(Ses i 6和Ses i 7)[9]。其中,Ses i 3由585 个氨基酸组成,分子质量约为45 kDa,属于Cupin超家族,具有β-折叠桶的蛋白质超二级结构,由单个肽链的3 个单体结合成三角形扁平三聚体,能够抵抗热变性和酶消化,可被75%芝麻过敏患者的血清识别,是芝麻主要的过敏原之一[10]。
高纯度的芝麻蛋白Ses i 3在致敏性及致敏机理的相关研究中具有不可或缺的地位。然而,从芝麻中提取高纯度天然Ses i 3会遇到不可避免的问题。繁杂的提取步骤可能会使蛋白降解损失,同时活性下降。在提纯过程中,过敏原蛋白与非过敏原蛋白同时存在,制备高纯度单一过敏原蛋白的难度系数较大。此外,天然过敏原蛋白在保存过程中,部分酶可能会将其降解,导致其生物活性降低[11]。这些原因使提取天然过敏原蛋白难以标准化和量产化。相比之下,通过重组技术获得的食物过敏原蛋白可以较好地规避这些问题。近年来,因重组蛋白具有纯度高、产量大及理化和免疫学性质与天然过敏原蛋白相当等特点,研究学者进行了多种食物过敏原蛋白的重组表达研究,并取得较理想的结果[12-13]。
因此,本研究从芝麻中提取总RNA,用反转录聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR)得到芝麻蛋白Ses i 3基因,构建pET-22b(+)质粒表达载体,转入BL21(DE3)感受态细胞宿主表达菌中诱导表达,对芝麻过敏原Ses i 3蛋白进行重组表达和纯化,并进一步建立BALB/c过敏小鼠模型对其免疫学特性进行鉴定,以期为芝麻过敏蛋白Ses i 3的量产化提供理论依据,同时也为后续芝麻致敏机理和芝麻脱敏研究奠定基础。
1 材料与方法
1.1 材料与试剂
LB平板组成成分:氯化钠10 g/L、胰蛋白胨10 g/L、酵母提取物5 g/L,121 ℃灭菌20 min,固体培养基中另加入质量分数1.5%琼脂粉。
天然芝麻蛋白Ses i 3(纯度≥90%),实验室制备;辣根过氧化物酶(horseradish peroxidase,HRP)-羊抗鼠免疫球蛋白G1(immunoglobulin G1,IgG1)、HRP-羊抗鼠免疫球蛋白G2a(immunoglobulin G2a,IgG2a)、HRP-羊抗鼠免疫球蛋白E(immunoglobulin E,IgE)赛默飞世尔科技(中国)有限公司;白细胞介素-4(interleukin-4,IL-4)、白细胞介素-5(interleukin-5,IL-5)、干扰素-γ(interferon-γ,IFN-γ)、组胺酶联免疫吸附测试(enzyme-linked immunosorbent assay,ELISA)试剂盒 上海宝曼生物科技有限公司;PrimeScriptTMRT Master Mix、琼脂糖凝胶回收试剂盒、质粒小量提取试剂盒、限制性内切酶NdeI、XhoI、XbaI 通用生物(安徽)有限公司;HRP anti-6×His tag小鼠单克隆抗体英国Abcam公司。
1.2 仪器与设备
1600凝胶成像分析系统、EPS200蛋白垂直电泳槽上海天能科技有限公司;JY99-IIDN超声波细胞粉碎机 宁波新芝生物科技股份有限公司;GHP-9160培养箱 上海一恒科学仪器有限公司;TGL-16M台式高速冷冻离心机 上海卢湘仪离心机仪器有限公司;Multiskan FC酶标仪 赛默飞世尔科技(中国)有限公司。
1.3 方法
1.3.1 pET-22b(+)-Ses i 3表达载体的构建
根据GenBank数据库中芝麻过敏原Ses i 3蛋白的基因序列设计扩增引物。上游引物:5’-AATTTTGTTTAACTT TAAGAAGGAGATATACATATGGAAAGCAAAGATCCG GAACTGAAACAGTGCAAAC-3’;下游引物:3’-AGCC GGATCTCAGTGGTGGTGGTGGTGGTGCTCGAGGGCA TCTGCGCGACCCTGCTGCTGCTGGCGCGGGCC-5’。
用Trizol法提取芝麻总RNA[14],将芝麻在液氮中充分研磨后,取50 mg加入1 mL的Trizol混匀,随后转移至RNase-free离心管中室温放置5 min。加入0.2 mL氯仿后剧烈摇晃15 s,室温放置3 min。4 ℃、12 000 r/min离心15 min后,将上清液转移至新的RNase-free离心管,加入0.5 mL异丙醇,混匀后室温静置10 min。4 ℃、12 000 r/min离心10 min,弃上清液,加入1 mL的75%乙醇溶液洗涤沉淀。4 ℃、7 000 r/min离心5 min,弃上清液,放置5 min,加入50 μL无RNase水溶解,-80 ℃保存。
用PrimeScriptTMRT Master Mix试剂进行反转录反应,反应体系为5×PrimeScript RT Master Mix 2 μL,RNA≤500 ng,加入无RNase的双蒸水至10 μL。37 ℃孵育15 min,85 ℃、5 s使反转录酶失活。
反应体系:GPV8高保真DNA聚合酶0.5 μL,10×GPV8 buffer 5 μL,10 mmol/L dNTP Mix 1 μL,上下游引物2×2 μL,模板DNA 2 μL,双蒸水37.5 μL。扩增条件:96 ℃预变性3 min;95 ℃变性15 s,58 ℃退火15 s,72 ℃延伸20 s,23 个循环,72 ℃彻底延伸60 s。用1%琼脂凝胶糖电泳鉴定PCR产物,将目的条带割胶回收,用NdeI和XhoI酶切pET-22b(+)载体,用聚合酶连接酶切后的目的基因和载体片段构建重组表达质粒,转入BL21(DE3)感受态细胞,在含有卡那霉素的LB平板上筛选阳性质粒。将阳性菌落扩大培养,提取质粒进行XbaI和XhoI酶切鉴定,送至安徽通用生物公司进行测序后用BLAST工具对结果进行序列对比分析。
1.3.2 重组芝麻过敏原Ses i 3蛋白的原核表达
将测序正确的载体转染BL21(DE3)感受态细胞宿主表达菌中扩大培养,当菌液OD600nm值为0.6时,分别加入异丙基硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)至终浓度为0.2 mmol/L和1 mmol/L,分别在37 ℃和15 ℃以220 r/min培养4 h。将诱导完成的菌液3 000 r/min离心10 min,收集细菌沉淀,随后使用超声破菌缓冲液将沉淀破碎,超声破碎条件为功率300 W,超声3 s,停3 s,至菌液澄清透亮,此时为全菌蛋白。将超声破碎的菌液12 000 r/min离心15 min,上清液即为上清蛋白,沉淀为包涵体蛋白。将包涵体蛋白置于包涵体溶解液中4 ℃搅拌至无沉淀,再将溶解好的溶液置于透析袋中,放入尿素浓度依次为6、4、2、1、0 mol/L的磷酸盐缓冲液(phosphate buffered saline,PBS)中透析,逐步去除蛋白溶液中的尿素,完成包涵体蛋白的溶解复性,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)分析其表达情况。
1.3.3 重组芝麻过敏原Ses i 3蛋白的纯化及复性
将适量的镍离子树脂与溶解复性的包涵体蛋白混合[15],4 ℃混匀2 h后,将充分混合作用的溶液加入纯化柱中,用清洗缓冲液洗掉杂蛋白,清洗缓冲液为含有50 mmol/L咪唑和尿素、pH 7.4的PBS。随后再用洗脱缓冲液洗脱目的蛋白,洗脱缓冲液为含有500 mmol/L咪唑和尿素、pH 7.4的PBS,收集洗脱液,即为纯化的重组蛋白。
将纯化后蛋白复性至PBS中(含有300 mmol/L NaCl、10%丙三醇溶液,pH 7.4),过滤除菌。复性后蛋白进行SDS-PAGE和Western blot检测。电泳完成后,去除浓缩胶进行转印,30 V、45 min。使用1×PBST漂洗5 min共3 次,随后使用5%脱脂奶粉室温封闭2 h。经1×PBST漂洗3 次后,使用HRP anti-6X His tag小鼠单克隆抗体(1∶2 000)孵育1 h。最后加入增强型化学发光工作液,静置2 min后拍照显色。
1.3.4 芝麻过敏BALB/c小鼠模型
24 只BALB/c 小鼠按体质量随机分成3 组,分别为PBS 组(对照组),天然Ses i 3(native-Ses i 3,N-Ses i 3)组和重组芝麻蛋白Ses i 3(recombinant Ses i 3,R-Ses i 3)组,每组8 只。N-Ses i 3组与R-Ses i 3组小鼠分别在第0、7、14、21和28天经口灌胃1 mg/只N-Ses i 3和R-Ses i 3(含10 μg CT佐剂),PBS组小鼠则经口灌胃等体积PBS(含10 μg CT佐剂)。第42天,每只小鼠进行5 mg蛋白进行大剂量刺激。采样,进行相关过敏性指标检测,所有动物实验研究均经过扬州大学动物伦理委员会批准(伦理号:YXYLL-2023-091)。
1.3.4.1 过敏症状的测定
在大剂量刺激后测定小鼠的体温变化;同时,观察各组小鼠的临床症状表现并进行评分,具体如下:0 分=无症状;1 分=抓挠鼻子和头部;2 分=活力降低,活动减少;3 分=眼睛与嘴部周围浮肿,毛发竖立;4 分=刺激后抽搐或静止,呼吸困难;5 分=死亡[16]。
1.3.4.2 特异性抗体检测
对小鼠进行眼部内眦采血,1 200 r/min离心15 min取上清液得到血清。采用间接ELISA方法测定各组小鼠血清特异性Ses i 3的IgE、IgG1和IgG2a的水平[17]。
1.3.4.3 细胞因子及组胺检测
第42天对小鼠进行眼部内眦采血后置于含乙二胺四乙酸二钾抗凝剂的离心管中,1 200 r/min离心15 min取上清液得到血浆,用于测定小鼠组胺的含量。IL-4、IL-5、IFN-γ、组胺含量检测参照ELISA试剂盒说明书。
1.4 统计学方法
2 结果与分析
2.1 重组芝麻过敏原Ses i 3蛋白的原核表达
2.1.1 pET-22b(+)-Ses i 3表达载体的构建及测序
提取芝麻总RNA,检测其RT-PCR扩增产物,如图1A所示,在1 500~2 000 bp之间有一条明亮条带,大小与已公布的Ses i 3大小基本相符。将目的条带与pET-22b(+)酶切后连接,随后进行酶切鉴定。由图1B可知,泳道1中5 000~6 000 bp的条带为载体,1 500~2 000 bp之间条带为Ses i 3基因。测序和BLAST工具分析显示与已公布的Ses i 3基因序列相似度100%,表明Ses i 3基因已被成功克隆且已连接到pET-22b(+)表达载体上。图1C为pET-22b(+)-Ses i 3表达载体示意图,蛋白末端为6×His标签。
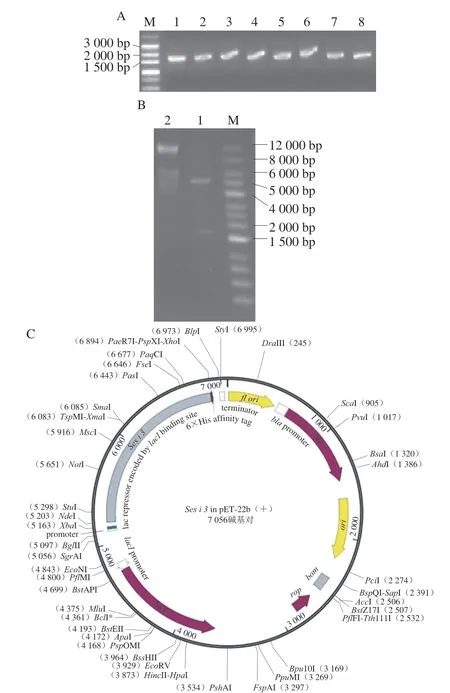
图1 Ses i 3基因的克隆(A)、pET-22b()-Ses i 3双酶切鉴定(B)和pET-22b()-Ses i 3图谱(C)Fig.1 Cloning of Ses i 3 gene (A),identification of pET-22b(+)-Ses i 3 (B) and profile of pET-22b(+)-Ses i 3 (C)
2.1.2 重组芝麻过敏原Ses i 3蛋白的表达及纯化
为了探究在不同诱导剂浓度和温度条件下重组表达载体目的蛋白的表达情况,分别在37 ℃和15 ℃条件下添加不同浓度的IPTG,进行重组蛋白的诱导表达,结果如图2A所示。泳道1~4表明,在相同温度下,0.2 mmol/L和1 mmol/L浓度的IPTG均可较好地诱导重组Ses i 3蛋白的表达,且表达量差异不大。鉴于IPTG具有一定的毒性,故选用0.2 mmol/L为IPTG诱导表达的浓度条件。泳道6~13表明,重组芝麻过敏原Ses i 3蛋白以包涵体形式存在,且分子质量约为66 kDa。此外,目的蛋白在37 ℃和15 ℃都是以包涵体的形式存在并且表达量差异不大,因此,选择大肠杆菌生长常规最适温度37 ℃作为诱导表达条件。包涵体蛋白溶解透析复性后,用镍离子吸附柱纯化带有His标签的重组芝麻过敏原Ses i 3蛋白。由图2B可见,经过纯化后,在66 kDa附近基本没有出现目的蛋白条带以外的其他蛋白,因此达到预期纯化的效果。包涵体蛋白通过纯化后进行复性,图2C表明,复性的芝麻过敏原Ses i 3蛋白条带单一,纯度较高,无杂蛋白,且复性后表达情况良好。Western blot结果(图2D)显示,蛋白与HRP anti-6×His tag小鼠单克隆抗体在大约66 kDa附近有明显结合,这表明重组Ses i 3蛋白具有一定的活性,可用于后续实验。
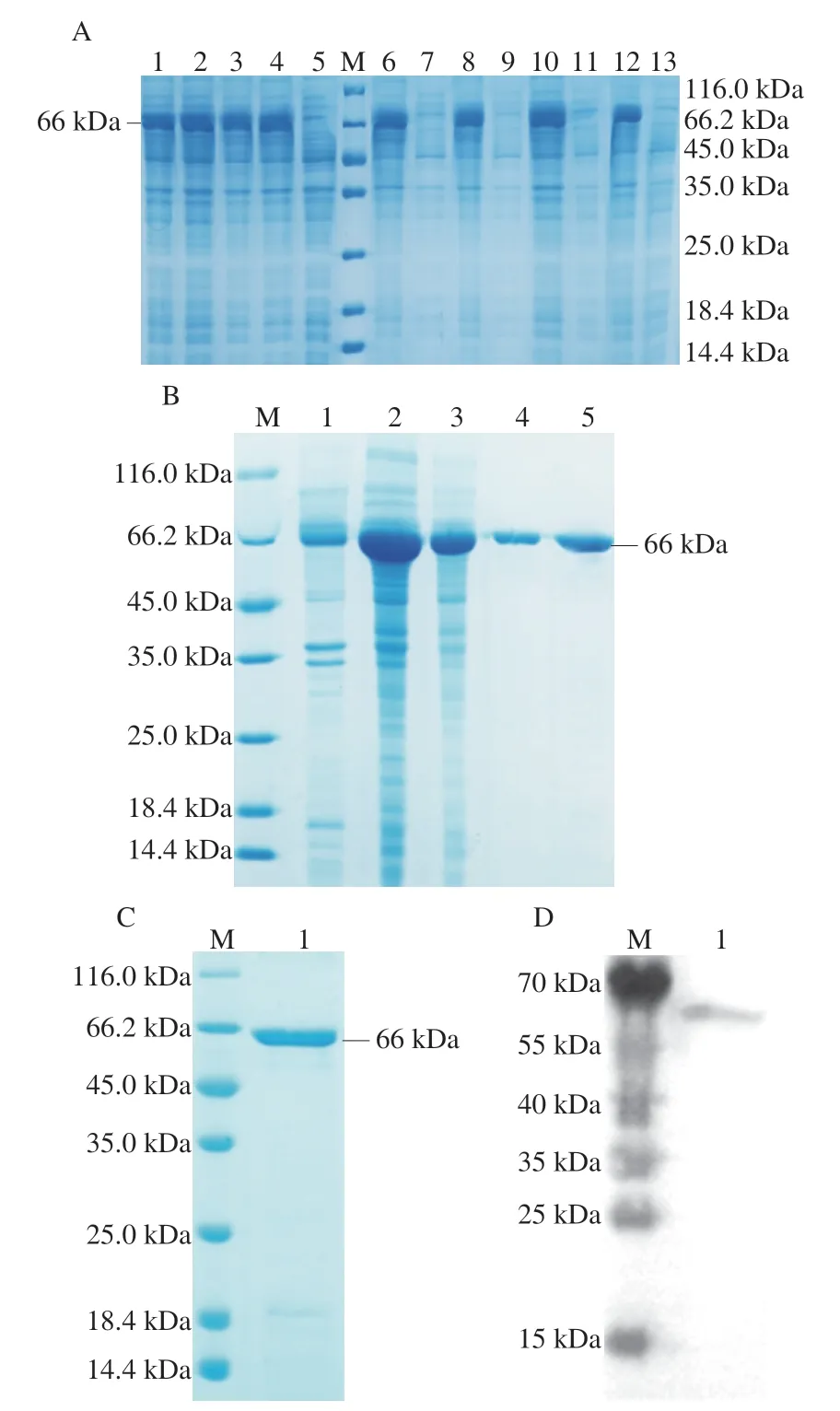
图2 重组芝麻过敏原Ses i 3蛋白的表达(A)、纯化(B)和复性(C)以及Western blot分析(D)Fig.2 Expression (A),purification (B),renaturation (C) and Western blot analysis (D) of recombinant Ses i 3
2.2 芝麻过敏原Ses i 3蛋白的致敏性鉴定
本研究采用BALB/c小鼠模型对R-Ses i 3的致敏性进行评估。如图3A所示,与PBS组相比,N-Ses i 3和R-Ses i 3组的小鼠经大剂量刺激后,体温均极显著下降(P<0.001),R-Ses i 3组的小鼠体温下降程度略强于N-Ses i 3,但无显著差异。小鼠过敏症状评分如图3B所示,N-Ses i 3和R-Ses i 3组绝大部分的小鼠表现出较为强烈的过敏症状,其中,1 只R-Ses i 3组的小鼠出现刺激后抽搐的症状。这表明,N-Ses i 3和R-Ses i 3都引起小鼠产生明显的食物过敏症状。
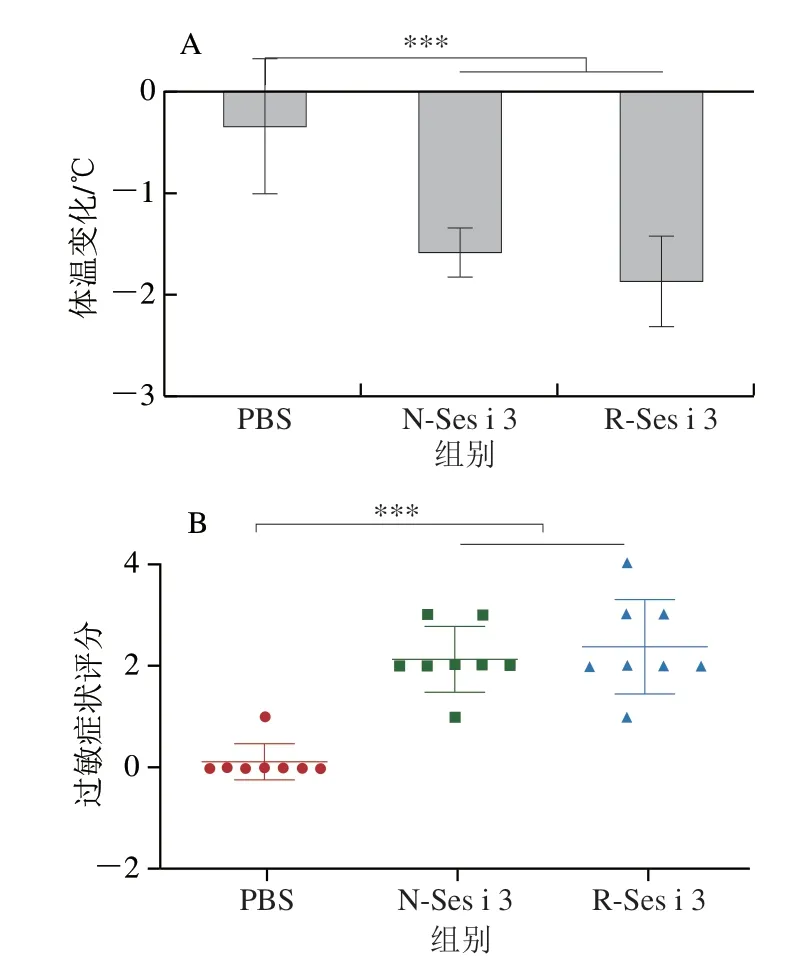
图3 小鼠体温变化(A)和过敏症状评分(B)Fig.3 Body temperature change (A) and hypersensitivity score (B) of mice
如图4所示,N-Ses i 3和R-Ses i 3组的小鼠血清中IgE和IgG含量均极显著高于PBS组(P<0.001),表明N-Ses i 3和R-Ses i 3都具有诱发小鼠芝麻过敏的能力,且在N-Ses i 3和R-Ses i 3组的小鼠血清中IgG1滴度明显大于IgG2a的滴度,表明在两组小鼠体内过敏反应均为Th2型。此外,N-Ses i 3和R-Ses i 3组的小鼠血清中特异性抗体的滴度基本相似,但IgG2a有轻微差别。
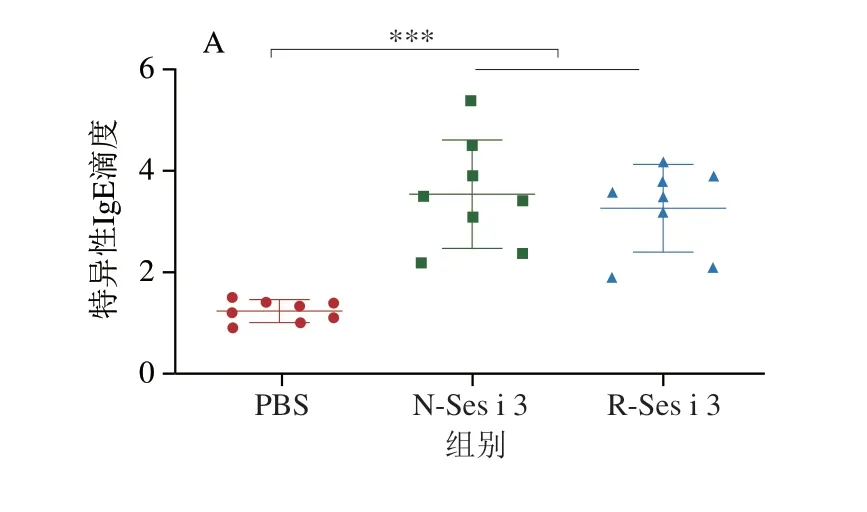
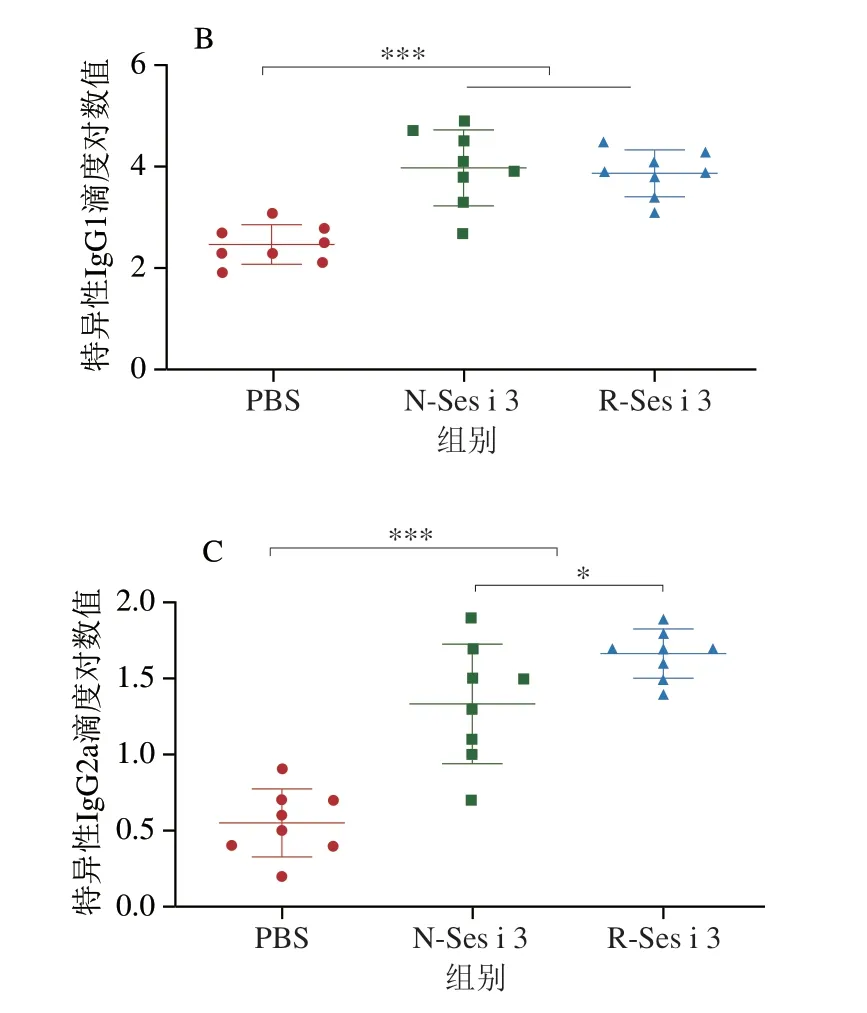
图4 小鼠血清中特异性IgE(A)、IgG1(B)和IgG2a(C)含量Fig.4 Specific IgE (A),IgG1 (B) and IgG2a (C) contents of mouse serum
如图5A~C所示,N-Ses i 3和R-Ses i 3组的小鼠血清中Th2型细胞因子IL-4和IL-5均显著高于PBS组(P<0.05),而Th1型细胞因子IFN-γ与对照组相比无明显差异,这表明N-Ses i 3和R-Ses i 3都可以通过上调Th2型细胞因子诱发小鼠Th2型过敏反应。此外,N-Ses i 3和R-Ses i 3组小鼠的血浆中组胺含量(图5D)较PBS组都极显著升高(P<0.001),这也证明N-Ses i 3和R-Ses i 3诱导小鼠脱颗粒的能力相似。
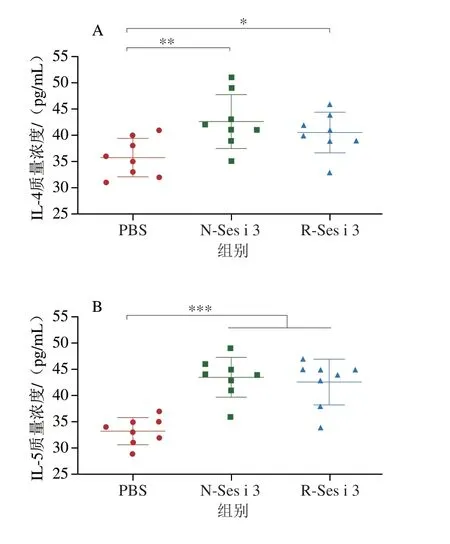
图5 小鼠血液中的细胞因子和组胺水平Fig.5 Cytokine and histamine levels in mouse serum
3 讨论
本研究对芝麻过敏原蛋白Ses i 3进行重组表达并发现其以包涵体形式存在,随后对包涵体蛋白进行变复性处理,最终得到纯度较高的R-Ses i 3。当重组蛋白在细胞中以包涵体形式表达时,虽然纯化步骤较可溶性表达繁琐,但也存在一定的优点,包涵体中的重组蛋白表达较高,变复性后可获得高纯度的蛋白[18-19]。重组表达系统主要有原核表达系统、真核表达系统和无细胞表达系统[20]。而大肠杆菌表达系统作为目前最成熟的表达系统[21],具有培养条件简单、生长繁殖快、基因操作容易、价格低廉以及快速生产目的蛋白等优点[22],该表达系统在生产花生、鱼虾等多种致敏性较强的食物过敏原重组蛋白中取得较大的进展。如程青丽等[23]利用BL21(DE3)大肠杆菌表达系统,重组牡蛎肌浆钙结合蛋白,并表明可用重组肌浆钙结合蛋白替代天然肌浆钙结合蛋白进行后续致敏性研究。此外,本研究中R-Ses i 3分子质量约为66 kDa,而N-Ses i 3分子质量约为45 kDa,推测可能与pET-22b(+)载体表达的融合标签及分子间和分子内的静电斥力有关[24]。本研究进一步采用BALB/c小鼠模型评估R-Ses i 3与N-Ses i 3的致敏性。小鼠过敏模型的建立是评估重组蛋白与天然蛋白致敏性差异的有效方法。史云凤[25]则采用Rosetta(DE3)大肠杆菌表达系统,成功重组了花生过敏原Ara h 1蛋白,通过建立BALB/c小鼠过敏模型证明重组Ara h 1与天然Ara h 1蛋白具有相似诱发BALB/c小鼠过敏反应的能力。这与本研究中利用BL21(DE3)大肠杆菌表达系统重组表达的R-Ses i 3与N-Ses i 3可以诱发小鼠芝麻过敏反应及脱颗粒的结果一致,可用于后续免疫学相关实验。
近年来,重组表达的食物过敏原蛋白在食物过敏机制研究及低致敏性食物开发研究上具有良好的应用前景。Sun Lirui等[26]的研究表明,与天然鱼类主要过敏原小清蛋白相比,重组小清蛋白具有批量表达灵活、纯度高、成本低、周期短等优点,且重组小清蛋白含有天然小清蛋白的大部分结合表位,可用于鱼类过敏的诊断及制备低致敏性鱼类产品的相关研究。Han Tianjiao等[27]研究表明,重组牡蛎过敏原肌浆钙结合蛋白具有和天然肌浆钙结合蛋白相似的理化性质及免疫活性。同时,云肖[28]的研究也表明,重组葡萄牙牡蛎原肌球蛋白与天然葡萄牙牡蛎原肌球蛋白具有相似的结构及免疫学性质。Tian Yang等[29]利用重组Ara h 1蛋白与葡萄糖胺进行糖化和糖基化反应,并证明可有效降低重组Ara h 1蛋白的抗原性,这可能与重组Ara h 1蛋白构象表位的丢失和游离氨基酸残基修饰其线性表位相关。此外,重组蛋白在定位过敏原蛋白的抗原表位上也具有较大的应用潜力。Fu Linglin等[30]通过重组中国对虾中的原肌球蛋白和精氨酸激酶,揭示了中国对虾原肌球蛋白的10 个抗原表位,精氨酸激酶的7 个抗原表位,以及3 个关键氨基酸(亮氨酸,天冬氨酸和谷氨酸)。Yang Yang等[31]采用BL21(DE3)大肠杆菌表达系统,重组克氏原螯虾肌球蛋白轻链亚型1蛋白,表明其具有完整的IgE结合活性和结构,可用于进一步的免疫学研究,此外,该研究还利用重组蛋白定义了3 个构象表位区域,并进行空间定位。
4 结论
本研究成功重组表达纯化了芝麻过敏原蛋白Ses i 3,并通过动物实验证实了其与天然Ses i 3蛋白具有相似的免疫学性质,可用于后续芝麻致敏性及致敏机制研究,为未来进一步制备低致敏性芝麻制品奠定了基础。
