封育对昆仑山北坡温性草原植物、土壤及微生物的影响
2023-12-04马富龙冯秀花周志勇何镇南席琳乔
马富龙, 韩 路, 冯秀花, 周志勇, 何镇南, 席琳乔*
(1.塔里木大学, 新疆 阿拉尔 843300; 2.新疆生产建设兵团第十四师昆玉市一牧场农业发展服务中心, 新疆 和田 848399; 3.塔里木大学动物科学与技术学院, 新疆 阿拉尔 843300)
新疆天然草地面积5.7×107hm2,占新疆土地面积的34.4%,是我国重要的牧区之一[1]。其中,可利用草地面积4.8×107hm2,占全国可利用草地的14.5%,仅次于西藏和内蒙古,居全国第3位。其中温性草原是新疆主要的草地类型之一,面积达4.81×106hm2,占新疆草地总面积的9.2%,占全国温性草原总面积的10.4%,是新疆农牧业和经济发展的宝贵资源。然而,随着全球气候变暖,人类需求不断增加,长期不合理利用草地资源[2],以及新疆的草原生态系统对持续变化外部环境响应的脆弱性和敏感性,使草地类型发生了不同程度上的退化。其中,过度放牧导致的草原面积占我国草地总面积的33%,草地退化会使草地生产和生态服务能力大大降低、可食牧草种类和产量减少、生物多样性降低及生态环境恶化。因此,恢复退化草地植被势在必行。围栏封育是草地植被恢复重要措施之一,既可恢复植被,也可改善土壤理化性状[3]和植物养分土壤-植物系统是草地生态系统中重要的组分之一,其在营养素的生物地化循环过程中发挥重要作用。Pang等[4]研究表明,植被-土壤系统中的碳、氮和磷元素含量之间相关性显著。杨振安等[5]指出不同草地管理方式(放牧或禁牧)会使土壤与植被养分分配及其平衡关系发生变化,并且植被与表层土壤主要养分含量之间的关联性仅存在于部分营养元素与部分植物器官之间。近年来,许多学者在温性草原上开展了大量的工作,主要围绕封育下温性草原群落结构与物种多样性[6]、群落稳定性[7]、功能群结构与生物量[8]、土壤理化性质[9]和种群空间分布格局[10]等方面进行了研究。对于退化后草地恢复,围栏封育是恢复退化草地行之有效的措施,在一段时间内可提高物种丰富度,提高植物地上和地下生物量,改善土壤理化性质[11-12],而且围栏封育较其他恢复措施具有改良效果较好、投资少的特点[13],能可持续运用于草地生态系统改良。张伟华等[14]研究表明围封可以显著提高土壤全氮、有机质和含水量。生长良好的草地不仅能促进对生态环境保护,而且有利于经济可持续发展[15]。土壤微生物对环境变化较为敏感,其多样性和群落结构对围栏封育的响应决定了退化草地恢复的程度[16]。斯贵才等[17]发现,围栏封育6年土壤细菌丰富度显著高于放牧,围封对土壤真菌群落结构也有影响[18]。因此,围栏封育是恢复退化草地植被的重要措施。但是,围栏禁牧对新疆极端干旱区温性草原植被恢复与土壤微生物多样性的研究鲜见报道。因此,探讨围栏对新疆极端干旱区温性草原植被与土壤微生物多样性之间的关系对植被恢复显得尤为重要。
本研究以极端干旱区温性草原为研究对象,研究围栏封育对草地植物与土壤微生物多样性及土壤理化性质的影响,分析比较围栏内外植被与土壤特性的差异及二者之间的关联性,阐明围栏封育对草地植被与土壤的恢复作用,以期为极端干旱区退化草地科学管理与恢复对策提供科学指导。
1 材料与方法
1.1 研究区概况
研究区位于新疆和田地区策勒县域内第十四师一牧场(36°00′~36°20′ N,80°21′~81°22′ E),天然草地呈斑块状分布在昆仑山北坡中段山地。该区年平均气温7.7℃,≥10℃年积温2 715℃,年降水量176.5 mm。研究样地地势平坦、退化程度类似、植被均匀,为典型温性草原,以高山老鹳草(GeraniumdonianumSweet)为建群种,主要物种有火绒草(LeontopodiumLeontopodioides)、二裂委陵菜(PotentillabifurcaLinn.)、新疆龙胆(GentianakareliniiGriseb.)、棘豆(Oxytropisochrocephala)、新疆银穗草(LeucopoaGriseb)、早熟禾(PoaannuaL.)与平车前(PlantagodepressaWilld.)等,群落盖度68%~98%。围栏面积为5 hm2,形状为不规则四边形。围封前均为当地牧民的夏季牧场,至2023年围栏封育整2年。通过手持GPS (GPSmap 60CSx,Garmin)实地测量获得海拔和经纬度数据,气温和降水等气象资料来自于努尔乡水文站。
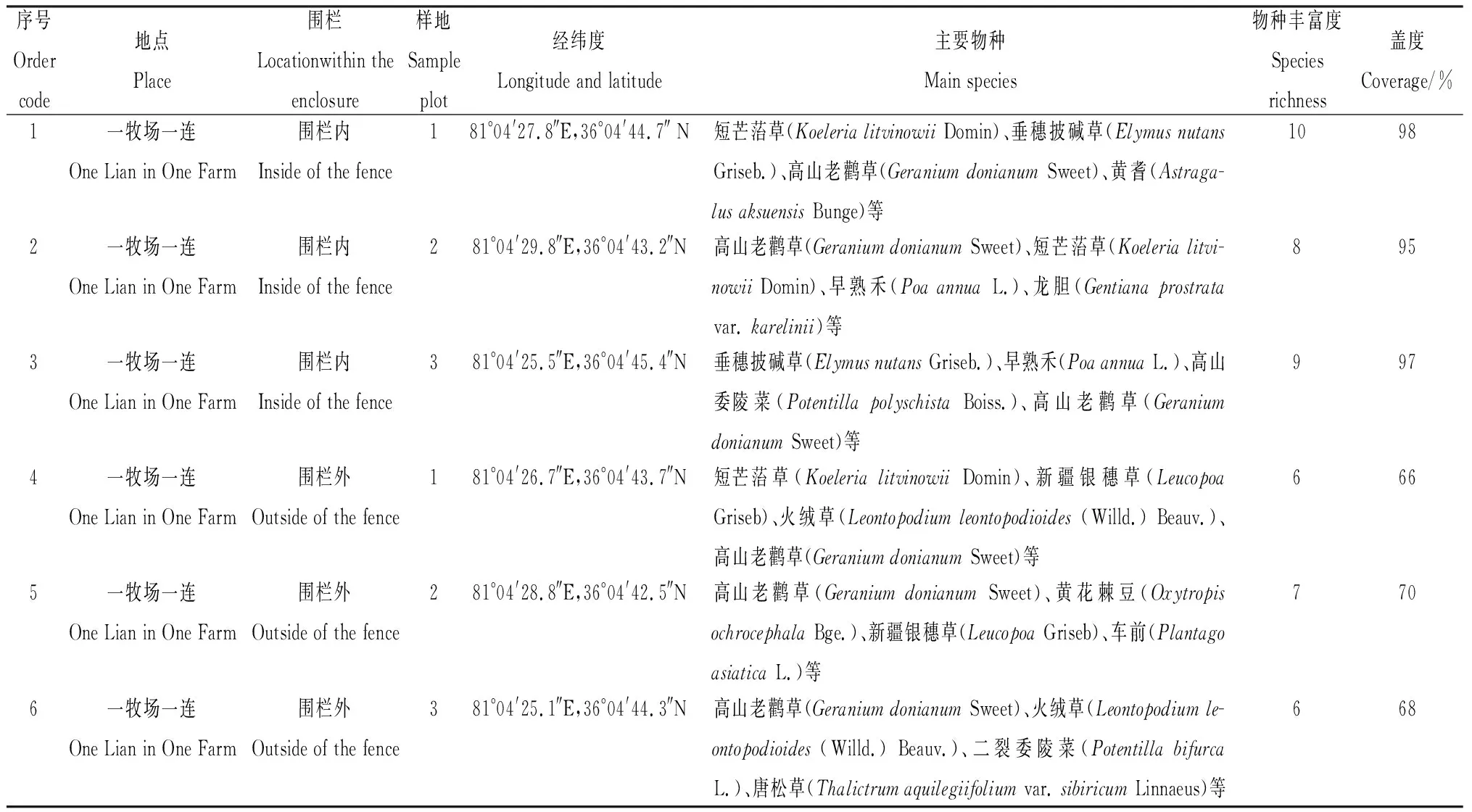
表1 研究区概况Table 1 Overview of the study area
1.2 试验设计
1.2.1植被调查 在围栏内外随机以半径为25 cm设置样圆,各调查30个样圆,记录每次样圆内出现的植物种名、株高与频度。
1.2.2生物量测定 围栏内外各样地随机调查3个1 m×1 m样方,记录样方内植物的种类、多度、盖度,并剪取样方内所有植物,分物种测量地上生物量。
1.2.3土壤取样 围栏内外为了避免不同物种的根际效应,随机选取5个采样点,用直径38 mm的土钻进行分层取样,每个采样点打三钻,取0~5 cm、5~10 cm、10~20 cm共3层土壤。将各层土壤中草根等杂质去除后,分成2份,一份带回实验室风干,备测土壤养分;另一份自封袋封装,冷藏箱内带回实验室,置于超低温冰箱保存,用于测定微生物。
1.3 测定指标
1.3.1植物多样性 重要值(IV)是群落中评价物种重要性的常用指标[19],计算公式如下:
式中:相对频度=调查样方内某种植物的频度/全部植物的频度之和;相对密度=调查样方内某一物种出现的总株数/该调查样方所有物种总株数;相对盖度=调查样方内某一种植物盖度/全部植物盖度总和;相对高度=调查样方内某一物种高度之和/该调查样方所有植株总高度;相对生物量=调查样方内某一物种地上生物量之和/该调查样方地上总生物量。
物种多样性指数计算Patrick指数、Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数[20],计算公式如下:
①Patrick丰富度指数(R):R=S
③Shannon-Wiener多样性指数(H):H=-∑PilnPi
⑤Margalef丰富度指数(MA):MA=(S-1)/lnN
式中,S为物种数目,N为群落中所有物种个体数目之和,Pi为代表i物种的重要值。
1.3.2土壤微生物DNA提取、扩增和高通量测序 称量0.5 g土壤,用TIANamp soil DNA kit试剂盒对土壤微生物总DNA提取。提取之后使用2%琼脂糖凝胶电泳及NanoDrop One检测提取DNA纯度和浓度,用无菌水稀释至1 ng·μL-1,稀释后的基因组DNA为模板扩增16S rRNA、ITS1区域,取1 μL DNA样本进行PCR扩增。扩增后的产物进行产品纯化,构建DNA文库,用llumina NovaSeq 6000测序平台进行测序。使用Trimmomatic v0.33软件,对测序得到的Raw Reads进行过滤;然后使用Cutadapt 1.9.1软件进行引物序列的识别与去除,得到不包含引物序列的Clean Reads;使用QIIME2 2020.6中的dada2方法进行去噪,双端序列拼接并去除嵌合体序列,得到最终有效数据(Non-chimeric Reads)。使用USEARCH(version 10.0)在相似性97%的水平上对可操作分类单元(OTUs)进行聚类。通过OTU序列比对Silva 132(细菌)、UNITE 8.0(真菌)数据库进行注释,可以得到每个OTU对应的群落组成分类信息,挑选出丰度最高的一条序列,作为该OTU的代表序列,使用QIIME 2.0软件计算分析物种组成和Alpha (α)多样性。以上测定委托北京擎科生物科技有限公司完成。
1.3.3构建微生物稀疏曲线 根据细菌16S rRNA基因、真菌ITS1基因高通量测序数据,按照序列相似度97%对有效序列进行OTU划分,按照每个OTU在每个样本中所包含的序列数,构建OTU丰度矩阵,并对获取样本的序列总数逐一进行随机抽样,以抽取到的序列数为横坐标与其对应的OTU数为纵坐标构建稀疏曲线。
1.3.4土壤理化性质测定 风干土壤样品研磨粉碎后过筛(2 mm),利用电位法以5∶1的水土比测定土壤pH值和电导率;重铬酸钾容量法-外加热法测定有机质;半微量凯氏定氮法测定全氮;0.5 mol·L-1NaHCO3浸提-钼锑抗比色法测定速效磷;NH4OAc-火焰光度法测定速效钾[21]。
1.4 数据的统计分析
根据物种注释结果,选取每个样本或分组在门(Phylum)水平上最大丰度排名前10的物种生成柱状图,以便直观查看样本在门水平上的细菌、真菌相对丰度。用Excel软件对试验数据进行初步整理,用SPSS 26.0单因素方差分析(One-Way ANOVA)检验围栏内外土壤理化性质和植物、土壤微生物多样性的差异显著性(Duncan法,α=0.05)。采用Pearson相关分析植物多样性、土壤微生物多样性与土壤理化性质之间关系,显著性区间定义为95%水平,图表中数据为平均值±标准误。利用Origin 2021进行绘图。
2 结果与分析
2.1 围栏封育对草地群落植物物种组成的影响
本次调查温性草原群落共出现17种植物,隶属10个科,15个属(表2)。

表2 围栏封育对温性草原物种群落的影响Table 2 Effect of fencing on the communities in temperate grasslands
表3可见,围栏内优势种为高山老鹳草,亚优势种短芒菭草、银穗草。围栏外以短芒菭草为优势种,银穗草、高山老鹳草和火绒草为亚优势种。围栏内药用价值植物(高山老鹳草、黄耆、车前、龙胆)和优良牧草(垂穗披碱草、早熟禾)的重要值均大于围栏外,而围栏外杂类草(二裂委陵菜、高山委陵菜)、有毒有害草类(黄花棘豆)的重要值均大于围栏内。
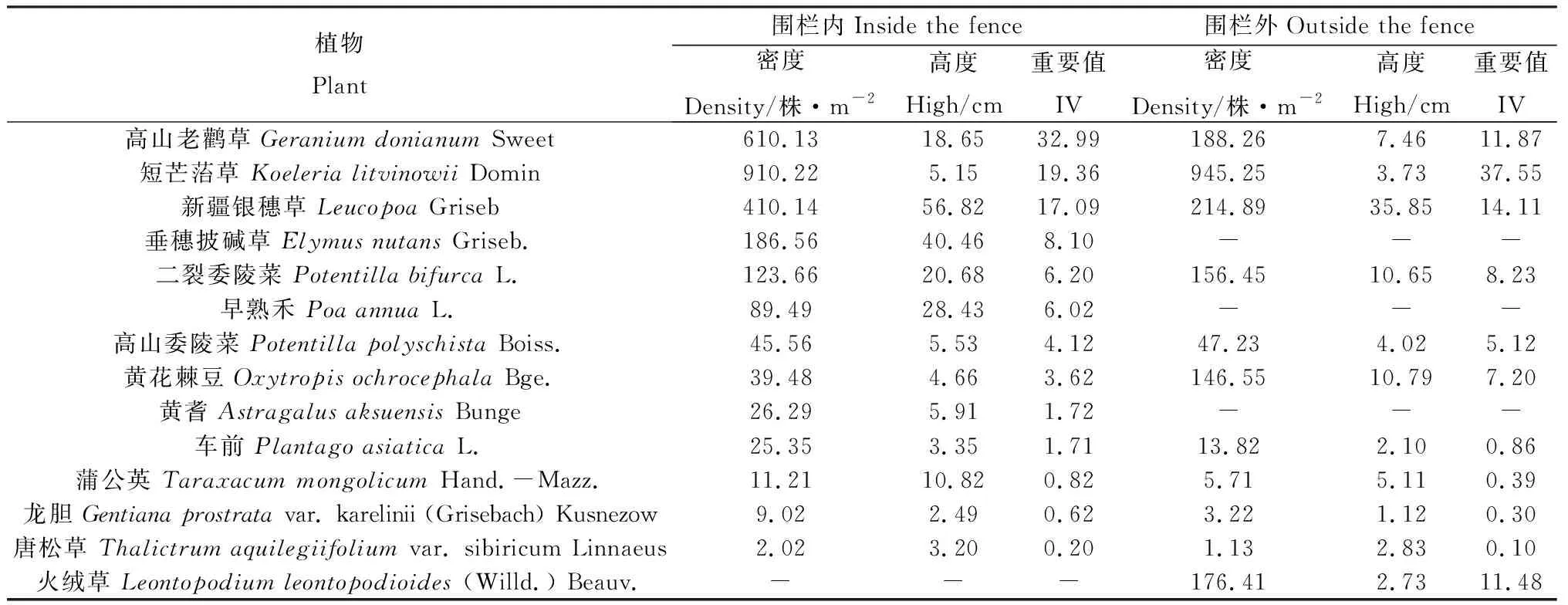
表3 围栏封育对温性草原物种组成的影响Table 3 Impact of fence on grassland plant diversity
2.2 围栏封育对温性草原物种多样性的影响
围栏内植物群落地上生物量(836 g·m-2)、盖度(97%)和Margalef指数极显著高于围栏外(P<0.01)(表4),Shannon-Wiener指数显著高于围栏外(P<0.05);围栏内外的Simpson指数、Pielou指数差异不显著。
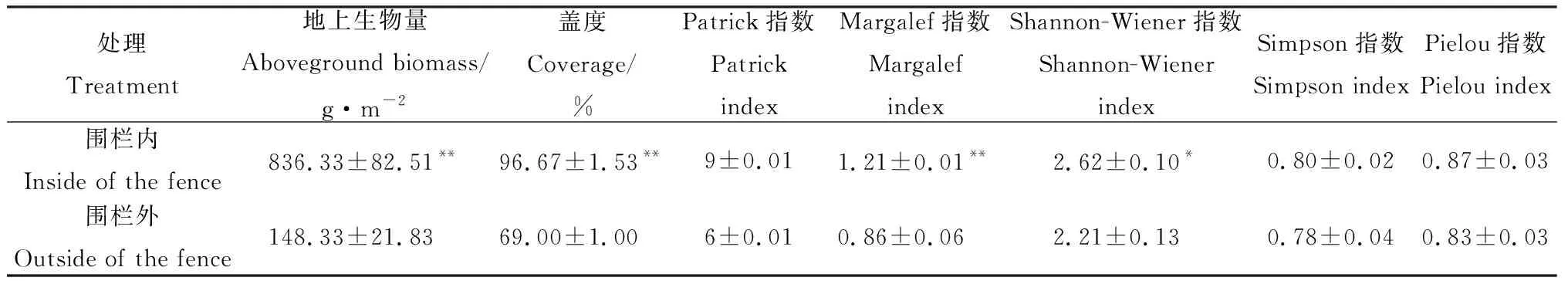
表4 围栏封育对草地群落物种多样性的影响Table 4 Quantitative characteristics of grassland vegetation inside and outside of the fence
2.3 围栏封育对温性草原土壤理化性质的影响
与围栏外相比,围栏封育显著提高0~20 cm土层速效磷、速效钾与总盐含量(P<0.05),显著增加了0~10 cm土壤有机质和0~5 cm土壤全氮含量(P<0.05);相反,围栏外0~20 cm各土层的pH值均高于围栏内,其中围栏内外10~20 cm土壤pH值差异显著(P<0.05) (图1)。垂直方向,土壤总盐、有机质、全氮、速效磷和速效钾均随土层加深呈降低趋势,围栏封育显著影响土壤表层(0~5 cm)有机质、全氮、速效磷与速效钾含量。
2.4 围栏封育对温性草原土壤微生物的影响
2.4.1围栏封育对温性草原土壤细菌、真菌多样性的影响 根据细菌16S rRNA基因高通量测序数据,围栏内(Mpi)6个样品,共含有356 878条有效序列,围栏外(Mpo)6个样品,共含有344 657条有效序列;根据真菌ITS1基因高通量测序数据,围栏内(Mpi)6个样品,共含有442 906条有效序列,围栏外(Mpo)6个样品,共含有455 445条有效序列。构建稀疏曲线(图2)后可知,围栏内外各土壤样品稀疏曲线均趋于平缓,可以认为测序深度已经覆盖到样品中所有的物种,测序结果足够反映各样本所包含的多样性。
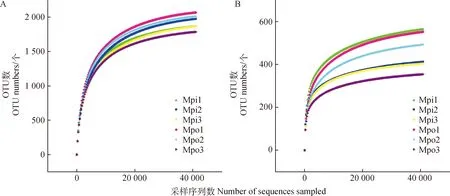
图2 围栏内外草地土壤各层细菌(a)、真菌(b)稀疏曲线分析Fig.2 Analysis of sparse curves of bacteria (panel a) and fungi (panel b) in different layers of grassland soil sampling from inside and outside of the fence注:Mpi为围栏内,Mpo为围栏外;1,2和3表示不同土层,分别为0~5 cm、5~10 cm和10~20 cm。下同Note:Mpi denotes inside the fence,Mpo outside the fence. 1,2 and 3 represent soil layers:0~5 cm,5~10 cm and 10~20 cm,respectively. The same as below
根据细菌OTU,ACE指数和Chao1指数、Simpson指数、Shannon-Wiener指数和PD whole tree指数,综合评估围栏内外土壤细菌a多样性(表5)。与围栏外相比,围栏封育2年显著提高了土壤细菌的OTU、ACE指数、Chao1指数(P<0.05);Shannon-Wiener指数提高了1.65%,PD whole tree指数提高了10.28%。
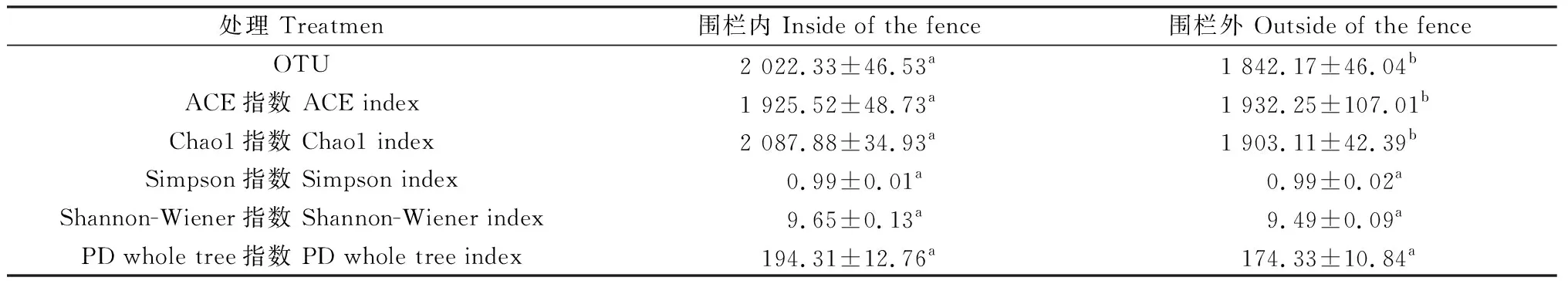
表5 围栏封育对温性草原土壤细菌a多样性的影响Table 5 Diversity of soil bacteria a in each soil layer inside and outside the fence
根据真菌OTUs、ACE指数和Chao1指数、Simpson指数、Shannon-Wiener指数和PD whole tree指数,综合评估围栏内外各土层土壤真菌a多样性(表6)。与围栏外相比,围栏2年显著提高土壤真菌的Simpson指数、Shannon-Wiener指数(P<0.05);OTU、ACE指数、Chao1指数、PD whole tree指数分别提高了4.33%,1.01%,3.30%,3.13%。

表6 围栏内外土壤真菌a多样性Table 6 Diversity of soil fungi a in each soil layer of the inside and outside the fence
2.4.2围栏封育对温性草原土壤细菌、真菌群落组成的影响 围栏内外细菌、真菌优势类群的相对丰度存在差异(图3a,b)。围栏内外细菌中变形菌门(Proteobacteria)与真菌中子囊菌门(Ascomycota) 相对丰度占比均最高,围栏内其相对丰度高于围栏外,二者相差8%,6%;且围栏内外其相对丰度均随着土层深度增加呈降低趋势。围栏内外细菌中酸杆菌门(Acidobacteriota)与真菌中担子菌门(Basidiomycota)相对丰度占比位居第2,围栏外0~20 cm土壤中二者相对丰度大于围栏内,高出1%,9%。细菌中拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)及真菌中未定名真菌(Unidentified),三者在围栏内外相对丰度占比位居第3。细菌中酸杆菌门(Acidobacteriota)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)及真菌中子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、未定名真菌(Unidentified)在围栏内外的土壤中相对丰度较高,为围栏内外的优势菌群。另外细菌中放线菌门(Actinobacteriota)、疣微菌门(Verrucomicrobiota)、未分类的物种(Unclassified_Bacteria)、其他菌门(Others)及真菌中被孢霉门(Mortierellomycota)、壶菌门(Chytridiomycota)、其他菌门(Others)在围栏内外的土壤中相对丰度超过3%,它们在围栏内外差异较小,这些菌门是温性草原土壤微生物群落的主要成分。
2.5 植物、土壤微生物多样性与土壤理化性质的关系
2.5.1植物多样性与土壤理化性质的关系 围栏内植物Shannon-Wiener、Pielou指数与土壤速效磷含量呈极显著正相关(P<0.01),Shannon-Wiener、Pielou指数与土壤总盐、有机质含量均呈显著正相关(P<0.05),群落盖度与全氮含量呈显著正相关(P<0.05);而植物多样性指数与pH值、速效钾含量的相关性均不显著(表7)。围栏外植物Pielou指数与总盐、速效钾含量呈显著正相关 (P<0.05),而Shannon-Wiener、Margalef指数及盖度与土壤pH值均呈极显著负相关(P<0.01),Simpson指数与pH值则呈显著负相关(P<0.05)。植物多样性指数与土壤有机质、全氮、速效磷含量均无显著相关性。
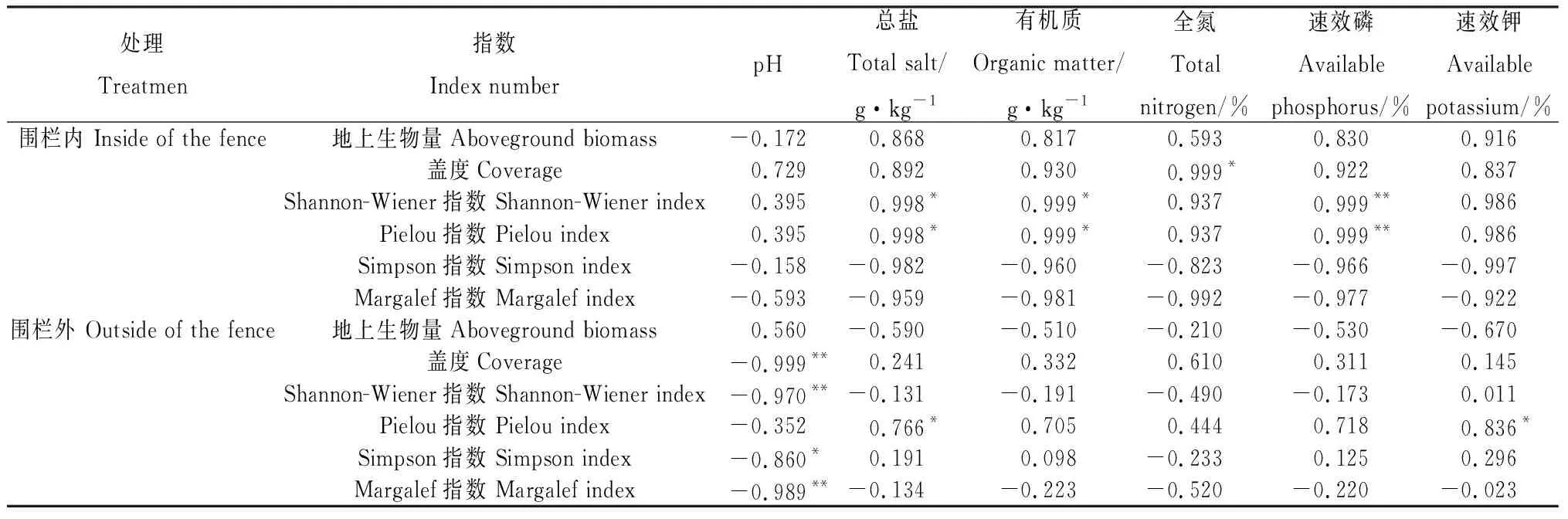
表7 围栏内外植物多样性与土壤理化性质的相关性Table 7 Correlation between the characters of plant community and soil physical and chemical properties
2.5.2土壤微生物多样性与土壤理化性质的关系 图4(a)可见,细菌Shannon-Wiener指数与土壤总盐呈极显著正相关(P<0.01),与全氮、有机质、速效磷、速效钾呈显著正相关(P<0.05),而与pH值呈负相关(P<0.05);细菌Simpson指数与土壤总盐、速效磷呈显著正相关(P<0.05)。图4(b)可见,真菌OUT数、Chao1指数与土壤有机质呈极显著正相关(P<0.01),与速效磷、pH分别呈显著正、负相关(P<0.05)。真菌ACE指数与土壤有机质呈显著正相关(P<0.05),真菌PD whole tree指数与土壤总盐、速效磷呈显著正相关(P<0.05)。
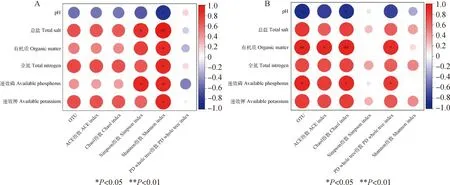
图4 土壤细菌、真菌的多样性指数与土壤理化性质的相关性Fig.4 Correlation between diversity indices of soil bacteria and fungi and soil physicochemical properties
2.5.3植物多样性与土壤微生物多样性的关系 图5(a)、(b)可见,植物地上生物量、盖度与细菌OUT数、Chao1指数、ACE指数及真菌Shannon-Wiener指数呈极显著正相关(P<0.01),与真菌Simpson指数呈显著正相关(P<0.05)。植物Margalef指数与真菌Shannon-Wiener指数呈极显著负相关(P<0.01),与细菌OUT数、Chao1指数、ACE指数及真菌Simpson指数呈显著负相关(P<0.05)。植物Simpson指数与细菌PD whole tree指数呈显著负相关(P<0.05),植物Pielou指数与真菌OUT数、Chao1指数呈显著负相关(P<0.05)。
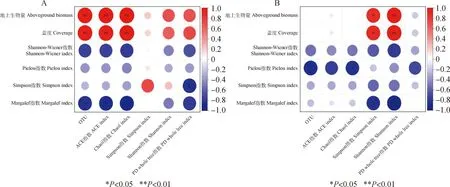
图5 围栏内外植物多样性与土壤细菌、真菌多样性的相关性Fig.5 Correlation between diversity indices of soil bacteria and fungi and plant diversity indices
3 讨论
3.1 围栏封育对干旱区温性草原植物群落与土壤理化性质的影响
围栏封育减少了动物的啃食与踩踏,是恢复草地生态系统的重要措施之一[22]。孙崇之等[23]研究发现,围封能极显著提高温性草原植被盖度、生产力与物种多样性。围栏后草地得到休生养息,促进了植物分蘖、生长与丛幅增大,从而显著提高草地植被盖度[24-26];同时,围封阻止了草食家畜的频繁采食与践踏,物种快速生长与扩繁,增加光合面积和碳同化能力,使物种高度、盖度、多度以及生物量都快速增加,从而促进草地生产力、盖度及Pielou均度指数和Shannon-Wiener多样性指数显著增加。因此,围栏封禁是恢复退化草地和提高生产力的有效措施。本研究结果发现,围栏封育促进了牻牛儿苗科、禾本科等生活能力强的优势种生长,且多年生草本的重要值提高,而一、二年生草本的重要值降低;围栏外高山老鹳草、早熟禾、垂穗披碱草等优质牧草重要值降低,二裂委陵菜、高山委陵菜等杂草及黄花棘豆等有毒有害草占比增加,有占据优势地位的趋势,表明围栏封育措施有利于优质牧草的生长,提高优势牧草比例,促进草地恢复。围栏2年后草地群落多样性指数高于围栏外,符合“中度干扰假说(Intermediate disturbance hypothesis)”,即中等干扰有利于物种共存并能维持物种的高多样性,然而,超过一定阈值的干扰则会打破物种平衡,降低物种多样性。由于围栏外作为当地牧民夏季牧场,受家畜的频繁踩食使牧草的生物量降低,影响其物质能量积累,进而影响了植物的生长繁殖,造成群落中禾本科植物多度、高度及盖度降低;同时频繁采食、抵制优势草生长,促进火绒草、黄花棘豆等适口性差、具毒杂草类生长与增多,降低草场质量。围栏封育能极显著提高温性草原地上生物量与盖度,改变群落物种组成与增加禾草类适口性牧草。从动物轮牧食物资源量的方向来看,建立围栏有益于当地畜牧业的发展[27]。
土壤是陆地生态系统的组成部分[28],是植被茁壮成长的立地条件,是生物与非生物环境之间进行物质与能量交换的重要枢纽[29]。围栏封育提高0~20 cm土壤有机质、全氮、速效磷和速效钾含量且降低土壤pH值,与朱牛等[30]研究结果相同。围栏减少了放牧啃食的影响,提高了草地地上生物量和植被盖度,增加植物凋落物,并在土壤微生物的作用下使土壤养分增加;围栏外家畜啃食、踩踏使土壤疏松沙化,植物生长受抑,导致土壤理化性质与草地生产力降低,加速草地生态系统退化,毒草比例增大[31]。此外,由于家畜踩食,围栏外植被覆盖率低,地表裸露蒸发量大,土壤水分减少,导致地表聚集少量碳酸钙使土壤pH值升高[32]。围栏封育2年显著提高土壤养分含量,土壤质量提升进一步促进植物生产力的增加,从而有利于退化生态系统恢复。
3.2 围栏封育对土壤微生物多样性的影响
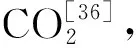
真菌具有分解有机质、转化养分等作用,是土壤微生物的重要组成部分。动植物残体主要依靠腐生真菌来分解,土壤真菌的多少与土壤有机质含量多少密切相关[39]。本研究发现,围栏内外温性草原土壤真菌均以子囊菌门为主,占比达到30%~50%,但围栏封育后子囊菌门的比例较围栏外提高5.4%,与前人研究结果类似[40-41]。史功赋等[36]研究表明,子囊菌与担子菌有较强分解腐殖质的能力,其强分解能力使得二菌受土壤因子变化影响小,验证了在恶劣环境中子囊菌门与担子菌门有较强的适应能力。其次为未定名真菌、担子菌门、被孢霉门和壶菌门在围栏内外土壤中的相对丰度超过3%,是围栏内外土壤中主要真菌群落。与围栏外相比,围栏内被孢霉门、壶菌门、球囊菌门和油壶菌门的相对丰度较高。其中被孢霉门、壶菌门能分解木质素和纤维素、半纤维素,有增加土壤有机质和养分含量的作用;球囊菌门的真菌会侵入植物的根部,与植物形成丛枝菌根,帮助植物吸收土壤中的水分与无机盐。总体来看,与围栏外相比,围栏封育促进了真菌相对丰度增加。
3.3 植物、土壤微生物多样性与土壤理化性质关系
草地植物群落、土壤微生物多样性与土壤养分状况是相互影响、相互制约的,它们共同决定植物群落的变化,而围栏改变了地上植物与土壤之间的相互作用过程。细菌、真菌与土壤有机质、速效磷、全氮、速效钾及植物地上生物量、盖度、多样性呈正相关,表明有益细菌、真菌优势群增加能促进土壤碳、氮的固定、分解有机质、解磷等作用,提高了土壤养分含量[42],进而有利于草地植物生长与物种多样性、盖度、地上生物量增加,提高优势种、适口性牧草比例,促进草地植物群落稳定与生态功能提升。同时,植物-土壤反馈作用得以发挥,围栏封育后植物凋落物与根系分泌物增多,能够给微生物提供更多的养分,促进微生物的生长繁殖,在门水平上优势菌门丰度明显升高[43],进而促进其新陈代谢的效率的增加,促进土壤养分释放与质量提升。围栏封育促进有益细菌的分解作用,促进土壤养分的增加,进而促进植物的生长和生产力提高。因此,围栏封育促进植物-土壤(微生物)之间的反馈协同作用,提高植物在环境极其恶劣条件下的生存概率,丰富物种种类,实现草地生态系统物质良性循环,从而促进退化草地恢复与生态功能提升,实现保护生态环境的目的。
4 结论
围栏封育可促进优势种生长与增加优质牧草比例及提高植被盖度、生产力,并可抑制有毒有害类杂草生长,从而优化温性草原植物群落结构组成,增加物种多样性。围栏封育后草地放牧、践踏土壤等干扰受到控制,草地经过休生养息,增加土壤微生物多样性、有益菌群与促进凋落物分解、沉积态矿质元素释放,提升土壤质量,进而促进植物生长发育与提高草地生产力;植物-土壤(微生物)间的反馈调节全面促进温性草地生态系统恢复。因此,围栏禁牧是恢复退化草地行之有效的措施,本研究为极端干旱区退化草地实施有计划的围栏封育、轮牧及科学恢复生态脆弱区草地生态系统提供理论依据。
