体重管理中的能量补偿:机制和意义
2023-11-30译校
康 杰(美),刘 畅(译校)
全球超过6.5 亿, 约占世界13%的人口受肥胖影响[1]。肥胖是诱发心血管疾病、高血压、癌症和2 型糖尿病等一些严重疾病的危险因素[2],其造成的医疗资源成本提高和带来的低生产力给国家经济带来一定压力[3-4],因此,肥胖被认为是21 世纪世界公共卫生界的“劲敌”[5-6]。 肥胖的产生可归因于多种因素,其中自主选择的生活方式带来的能量正平衡(即摄入的能量大于消耗的能量)是主要原因[7]。 长期研究证明,身体活动会对能量消耗产生影响,且在能量摄入方面起着关键作用[8]。
体重增加的主要原因是能量摄入与消耗的不平衡。 图1 显示了导致这种不平衡的各种能量成分的变化情况。 然而,能量平衡看似概念简单,实则非常复杂,其调节受生理、行为和环境等多方面因素的影响。 生理驱动的饥饿感、饱腹感与环境,均会影响饮食行为,继而作用于能量摄入。 同样,日常能量消耗是生理和行为相互作用的结果, 且二者同样会受到环境因素的影响。 静息代谢率 (Resting Metabolic Rate, RMR)和食物热效应主要由生理因素决定[9-10]。RMR 代表维持基本生命机能所需的能量,占一般人群日常能量消耗的大部分[9]。 然而,身体活动是日常能量消耗中最可变的组成部分,主要由人的行为决定。一般来说, 身体活动占日常能量消耗的25%~35%,但在持续剧烈运动的极端情况下可能达到75%[11]。为此, 一般通过进一步区分运动和习惯性身体活动或非身体活动产热(Non-Exercise Activity Thermogenesis, NEAT) 来解决身体活动性质差异的问题。 运动反映了针对特定目标的有计划、 结构化和重复性的活动[12]。 NEAT 反映了除主动运动以外所有身体活动的能量消耗,包括日常生活活动、小肌群运动、肌肉自主收缩和姿势维持[13]。
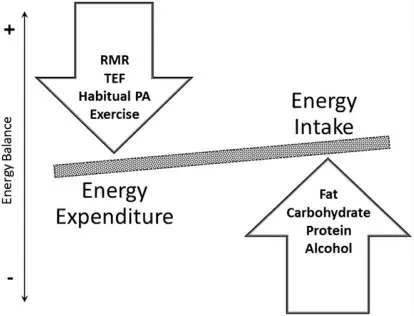
图1 对能量平衡起主要作用的因素Figure1 Major components contributing to energy balance
生活方式干预是治疗肥胖的基本方式[14],可在短时间内达到减重的效果(减少初始体重的5%~10%),包括限制饮食、身体活动和行为疗法[15-16]。 一般来说,有久坐习惯的人在日常生活中增加身体活动后,身体会更健康,同时可降低患慢性疾病的风险,且能更好地将体重控制在正常范围。 身体活动也会影响营养能量的分配, 能量分配仅指宏量营养素被吸收后会变成什么。如果一个人定期进行身体活动,摄入的能量就会被用于恢复和补充, 而不是储存为脂肪组织[17-19]。 此外,大多数人可以进行长时间、强度为2~16 倍RMR 的体育活动[20]。 因此,单次运动的能量消耗可达到250~2 500 kcal,且在一段时间内多次重复后,会因为显著的能量负平衡,达到减重的效果[20-21]。因此,更多的体力活动或锻炼消耗的能量,以及伴随的代谢通量是能量平衡的重要且可改变的决定因素, 多数人可以使用运动训练作为逆转和预防肥胖及其相关并发症的经济有效的解决方案。
遗憾的是,运动的减重效果通常远低于预期[22]。实际上,有研究报告在运动干预4 周后发现,无活动对照组和运动组之间的体重没有显著差异[23]。 也有研究发现,尽管运动组与对照组的耗能不同,但干预后体重减轻的程度相似[24-25]。最初成功减轻体重的人大多数会出现显著的体重反弹情况, 甚至回到初始体重[26-27]。 1999—2006 年美国国家健康和营养检查调查(National Health and Nutrition Examination Survey,NHANES)的数据显示,只有六分之一的超重或肥胖者报告在干预生活方式后1 年内仍能保持至少10%的减重效果[27]。
1 能量补偿的概念
对于减重结果通常低于预期的现象,可以用能量补偿现象来解释。如果每天运动且每次消耗300 kcal,即每日能量消耗增加300 kcal。 事实上,这是有关活动如何影响能量消耗的传统认知,进行的活动越多,到一天结束时燃烧的能量就越多。几个世纪以来,人们对“生理限制”或“物质补偿”的认识可以追溯到亚里士多德时代,达尔文也对此提出过相关理论,他认为并非所有器官都可以在同一时间处于高活动状态。现有大量数据表明,长期进行运动或其他体育活动的这种生活方式的改变, 不会相应提高日常能量消耗。相反,人类和动物在机体水平上会表现出一定程度的能量补偿, 即通过降低其他生物过程中的能量消耗来平衡日常能量消耗量[28-29]。从生命史理论所依据的分配原则[30]的角度上来看,能量补偿是可预期的,因为资源有限,生物体会将能量优先分配给生长发育、繁殖、喂养等关键生物过程[31]。
在不断变化的环境中维持内环境稳态, 这是机体最重要的生物学功能之一, 例如, 维持酸/ 碱平衡、血糖、身体水/ 电解质平衡和激素调节。 能量稳态是一个经常被忽视的调节过程, 人体一直在努力维持能量平衡。像其他维持内平衡的行为一样,维持能量平衡的能力可以被视为一种进化上的保守机制,尤其是保留身体能量储存以保持生殖功能,这在饥荒时期是有效的生存策略[32]。遗憾的是,这种维持能量稳态的能力, 对当今生活在发达国家的大多数人较为不利, 因为这个过程会对许多人期望/ 需要的减肥有抵制效应。 由运动或长期能量限制导致的负能量平衡所引起的代偿反应可能是代谢性的,如RMR 和NEAT 降低,即能量摄入增加,身体活动减少[33-35]。 图2 概述了当人体处于能量不足状态时,补偿机制如何恢复能量稳态(平衡)的过程,这与试图减肥时的情况一样。
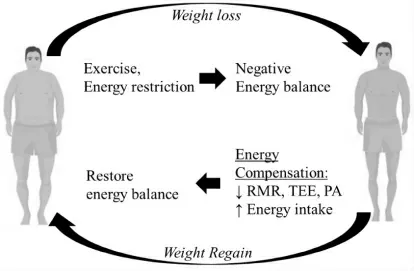
图2 包括RMR 的降低和能量摄入增加在内的伴随能量不足并减轻负能量平衡的功能, 从而抵抗进一步体重减轻的代偿机制Figure2 Schematic of how compensatory mechanisms,including decreases in RMR, TEE and PA, and increases in energy intake, accompany an energy deficit and function to mitigate negative energy balance and thus resist further weight loss
由运动引起的能量不足所带来的能量补偿,可以根据预期体重减轻与干预引起的脂肪质量(Fat Mass, FM)和去脂质量(Fat Free Mass, FFM)的实际变化之间的差异来量化。 FM 和FFM 的实际变化是由累积能量平衡(Accumulated Energy Balance, AEB)确定的,因为从长远来看,身体成分的变化反映了能量平衡的变化[25]。 据假定,增加1 kg FM 和1kg FFM分别等于12 000 kcal 和1 780 kcal[36],而减少1 kg FM和1kg FFM 分别为9 417 kcal 和884 kcal[37]。 期望的体重减轻是根据运动能量支出 (Exercise Energy Expenditure, ExEE)来确定的,后者是从训练引起的能量支出(Training-induced Energy Expenditure, TrEE)计算得出的,再加上15%的运动后多余能量支出[38]。其中,运动期间发生的静息能量消耗(REE×1.2)应该从TrEE 中减去。 因此,ExEE=(TrEE×0.15)+[TrEE-运动持续时间×(REE×1.2)][25]。 一旦获得了既代表实际体重减轻的AEB 又代表预期体重减轻的Ex-EE,就可以通过补偿指数(Compensation Index, CI)来预估补偿量,即由(ExEE-AEB)/ExEE×100%计算得出。 CI 为零,代表身体成分变化的AEB 等于因运动而消耗的能量(ExEE),即没有发生能量补偿。 当CI>0 时,表示发生了正补偿,表明运动引起的实际体重减轻低于预期。 当CI<0 时, 表示发生了负补偿,表明运动引起的实际体重减轻大于预期。
2 代谢性能量补偿机制
由前文可知, 通过运动或能量限制实现的能量负平衡可能会导致代谢过程出现不自主波动, 部分代谢过程足以抵消运动或饮食引起的能量不足。 这些不自主的代谢变化可能包括RMR 和NEAT 的降低,以及骨骼肌工作效率的提高,进而导致每日总能量消耗减少[32]。
2.1 静息代谢率(RMR)
RMR 是人体在完全休息时消耗能量的速度,通常可按24 kcal/h 估算[39]。RMR 是每日总能量消耗的最大组成部分(50%~70%),而FFM 约占60%~70%[40]。降低RMR 以补偿运动诱导的能量消耗增加的深层机制可能为:维持体内平衡所需的某些生理过程,如免疫能力、蛋白质表达和组织修复,以某种方式被降低优先级, 因此当能量补偿是一个更紧迫的稳态驱动因素时,会相应下调[41-42]。 在极端情况下,这种下调可能会减缓生长,或会导致疾病的发生发展,以及出现排卵和生殖受损[43]。 大多数人(非全部)广泛接受,在长时间的能量限制和随后的体重减轻期间,身体通过降低RMR 来节省能量, 并恢复能量平衡[32]。血液中儿茶酚胺水平的降低可进一步解释RMR 因能量不足而降低的机制。 降低儿茶酚胺水平可以减少葡萄糖被氧化的能量, 并促进更多的葡萄糖以糖原或脂肪的形式储存在体内[44]。
将能量代谢补偿归因于RMR 降低的论点也存在疑问。 增加活动水平的干预措施通常会导致身体成分的变化,例如体重减轻和FFM 水平增加[45]。 这可能很难证明由RMR 降低所带来的能量补偿,因为RMR 与FFM 呈正相关关系。事实上,在早期的研究中,参与者没有表现出体重因日常活动水平的增加而变化,或仅略有下降,且没有发现RMR 的变化[46-51]。在一些研究中,参与者的体重没有变化,但RMR 略有增加[52-55]。 未来研究在预估通过降低RMR 实现的能量补偿时,应控制体重和体成分。 只有报告RMR降低以及活动水平增加, 且体重没有实质性下降的研究才能合理地推断, 由于活动增加带来的能量补偿是由RMR 下调引起的。
2.2 非运动活动产热(NEAT)
NEAT 反映了除主动运动以外的所有身体活动带来的能量消耗,包括日常生活活动、小肌群运动、肌肉自主收缩和姿势维持[13]。 旨在了解运动干预对NEAT 影响的研究, 均对能量补偿涉及NEAT 的节能变化进行了假设。 如一项针对儿童的跨学校研究显示, 在学校进行的剧烈体育活动量与全天的总活动水平无关,这表明为补偿剧烈体育活动,NEAT 水平有一定程度的降低[55]。同样,一项针对老年参与者的研究发现,在参加体育锻炼干预期间,其日常活动水平并没有增加[56]。 最近一项针对超重女性的实验结果显示, 运动干预并没有引起每日能量消耗或RMR 的增加,这表明NEAT 的减少是补偿能量消耗的一个方面[49]。
通过提高骨骼肌工作效率或通过减少给定工作量每单位FFM 的能量,进一步解释了因能量不足而NEAT 有所减少的情况[57-58]。下丘脑-垂体-甲状腺轴活动降低后会引起骨骼肌工作效率的提高[32]。 减肥后瘦素的减少,导致下丘脑-垂体-甲状腺轴活动的衰减,进而降低甲状腺激素T3 的活跃性[45],这些激素的变化反过来通过降低心率、 血压和肌肉三磷酸腺苷(ATP)的产生来减少能量消耗[32]。 提高骨骼肌工作效率从而减轻体重的另一种机制, 是通过增加骨骼肌氧化脂肪来降低葡萄糖的转化能力[58-59]。有研究观察到磷酸果糖激酶和果糖-二磷酸醛缩酶的下调与能量限制的体重减轻有关, 而参与脂肪酸氧化的基因如3-羟基酰辅酶A 脱氢酶和脂肪酸结合蛋白有所上调[59]。有研究认为,基因表达的变化可以提高内源性脂肪作为燃料的利用率, 减少对葡萄糖的依赖,从而减少活动诱导的能量消耗,导致维持减轻的体重更加困难[59]。
值得注意的是, 也有研究结果显示NEAT 并没有随运动水平的提高而变化[60-61]或提高[62-63],但大多数这方面的研究没有直接对NEAT 进行测量。 最近的一些综述旨在确定人体是否会通过降低NEAT 以补偿运动诱导能量消耗的增加提出了相互矛盾的证据[64-66]。 有关NEAT 结果的差异,可能是由运动强度、方式、测量工具,以及研究对象的年龄与性别的不同而造成的。 如有研究发现老年人更可能表现出NEAT 的补偿性变化[53,67]。最近一项针对超重成年人的研究表明,NEAT 与运动量密切相关,仅高强度运动组(600 kcal/d)有减少,而中等强度组(300 kcal/d)未见变化[25]。早期研究多基于平均数据得出结论,但最近有证据表明,代偿反应的个体差异很大,这种差异解释了为什么NEAT 存在相互矛盾的结果, 同时对与减重相关的运动处方提出了个性化的要求。
3 行为能量补偿机制
行为能量补偿机制是对受某些神经行为机制而影响能量负平衡的自主反应[68]。 这些补偿机制包括增加能量摄入和减少主动体力活动, 前者是负责减少运动减肥时能量负平衡的主要机制[35]。
3.1 增加能量摄入
能量不足会通过激活/ 停用大脑的某些区域来影响进食欲望。随着体重减轻,响应能量负平衡而增加摄入的能量可能是由饱腹感的降低和对食物的反应神经信号变化引起的[69]。 减肥后大脑的奖励回路或通路的边缘系统区域更活跃[70-71]。这导致食物的奖励特性优先于抑制控制,并驱动饮食行为[72]。同时大脑活动的这种变化会触发食欲调节激素的产生[73]。能量负平衡或体重减轻已被证明会增加饥饿激素生长素释放肽的分泌和主观饥饿感[74-77],即使在部分体重恢复后也是如此[74]。此外,体重减轻已被证明会减少餐后饱腹感肽的分泌, 如活性胰高血糖素样肽1(GLP-1)、总肽YY(PYY)和胆囊收缩素(CCK)[74,78]。
尽管有证据支持能量摄入存在补偿性增加,但一些研究仍未能证明急性运动[79-81]或长期运动[82-84]后是否存在能量补偿这种适应性反应。此外,因为缺乏证据证明食欲的变化或血浆中相关激素的血浆浓度与体重恢复之间存在因果关系, 食欲的代偿性变化是否会导致体重恢复很大程度上仍是推测性的。 因此应谨慎解读与运动训练后能量和宏量营养素摄入的变化相关的文献。 能量摄入通常通过自我报告3~7 d 饮食记录进行评估,这种评估方式已被证明会低估能量的摄入。 如Donnelly 等[83]比较了双标水法和饮食记录法测量的能量摄入, 发现饮食记录少估算了约20%~30%的能量。
3.2 减少体力活动
限制非结构化体育活动时间可能会抵消运动期间消耗的能量或因能量限制而产生能量负平衡[85]。然而,这一假设尚未得到充分证实。一次性运动不会对有习惯性身体活动的人产生影响, 引起习惯性身体活动的任何变化[86-88]。Alahmadi 等[86]对久坐不动参与者进行一次运动干预2 d 后, 习惯性身体活动反而有所增加。然而,习惯性身体活动在长达4 个月的锻炼计划中被发现有所减少[46,56,89-90]。一般认为,由于会出现疲劳感和不适感, 特别剧烈的运动会导致习惯性体力活动的减少[91]。
Schutz 等[89]的研究也显示,随着运动量的增加,身体活动会有更大程度的减少。 在6 个月以上的干预中,习惯性身体活动没有变化[48,60,63,92]。缺乏习惯性体力活动的补偿性减少可能是由于在长期参与锻炼方案后健康状况的提高, 这会提高锻炼耐受力并减轻锻炼对习惯体力活动的负面影响[93]。
在评估身体活动以应对能量不足时, 训练计划类型是一个需要重点考虑的因素。 Hunter 等[54]观察到老年人抵抗运动引起的每日总能量消耗和习惯性体力活动有所增加, 身体活动的增加归因于功能能力的增加[61]。 相反,在剧烈运动或长期能量限制后,会出现肌肉酸痛或疲劳, 反而会导致久坐行为的增加,例如以乘坐电梯替代爬楼梯,减少体力活动和总能量消耗。 这可能会对运动处方的依从性有消极作用,降低减肥效果的可持续性。大多数运动干预依赖于有氧运动,因为与抗阻运动相比,有氧运动的能量消耗更大。因此,未来需要对各种运动类型对能量平衡相关性的影响的潜在差异进行研究。
整合所有与身体活动反应相关的研究发现,短期有氧运动干预可以潜在地引起习惯性身体活动的代偿,特别是对老年人和亚健康人群。 然而,随着体适能和功能能力的提升, 抗阻训练和长期运动训练可能会维持或增加习惯性身体活动。
4 运动与能量平衡
运动是减肥的常用疗法。 美国运动医学学会建议想要减肥的成年人每周进行225 min 的适度身体活动[94]。 然而,如上所述,由于补偿机制在努力维持能量平衡, 运动引起的体重减轻通常远低于基于运动消耗能量的预期。 运动可以以不同的强度和量进行。因此,其所引起的代谢和行为适应可能会有很大差异。 下文更具体地概述了运动是如何影响能量补偿, 以及如何通过使用不同的运动方式以最大限度地减少这种适应性反应的文献。
4.1 运动方式
健康研究领域对运动模式如何影响机体对运动的反应非常感兴趣, 因为这对制定有效的运动处方有很大帮助,以将能量补偿的影响降到最低,从而最大限度地减轻体重。 高强度间歇训练(High Intensity Interval Training, HIIT) 是在大于80%VO2max的强度下进行短暂的、激烈的、尽最大努力的运动,且运动时间与间歇时间的比值大于1[95]。 与传统的中等强度有氧运动相比, 这种训练模式因为引起的能量补偿较小而更具优势。 HIIT 被认为与习惯性身体活动的增加有关,进而增加每日总能量消耗[96-97]。 HIIT 还可以通过减少运动后的食物摄入量和食欲感而减少补偿反应[98-99]。 有研究表明,与中等强度间歇训练相比,HIIT 引起的饥饿感和进食欲望更低, 且会导致对高脂肪食物欲望的降低[100]。 这些对宏量营养物质喜好的改变可能是HIIT 引发脂肪质量更大减少的一个原因,即使是能量消耗小于或等于中等强度间歇训练[100]。 HIIT 也可以通过引起更多的运动后氧气消耗来减弱代谢补偿, 从而增加每日总能量消耗[101-102]。与中等强度运动相比,HIIT 仅需要近一半的运动时间来燃烧等量的能量[100]。 随着现代社会闲暇时间的减少, 能够在保持能量消耗增加的同时缩短训练时间具有重要价值。
抗阻训练是一种通过外部负荷为骨骼肌提供渐进式负荷, 以提升力量、 增加骨骼肌维度的运动形式。抗阻训练可带来的益处有很多,包括降低血脂和血压、促进骨骼肌维持/ 生长、改善血糖水平和胰岛素敏感性,以及促进脂肪量的减少[103]。 由于抗阻训练在减肥期间起到保存FFM 的作用,因此它可以消除或减弱代谢代偿反应, 例如饮食或运动引起的体重减轻中常见的RMR 下降[104]。 事实上,与单纯有氧运动相比, 抗阻训练或抗阻结合有氧训练的方案会增加RMR[105]。 一项16 周抗阻训练与有氧运动的比较研究表明,抗阻训练会促进非运动性身体活动,特别是在非运动日, 而有氧运动组的身体活动会出现补偿性减少[106]。 在比较抗阻训练和有氧运动之间能量摄入的补偿性增加差异时,需要考虑性别因素,而即使在控制运动能量消耗的情况下, 也只有男性在抗阻后更容易进行补偿性进食[93]。与有氧运动相比,抗阻训练对身体成分的影响结果更具差异性[107],当与足够的蛋白质摄入相结合时, 男性能量摄入的补偿性增加可能与抗阻训练的合成代谢性质和随后的肌肉质量增加有关[108-109]。因此,抗阻训练增加能量摄入可能是由肌肉生长和修复引起的, 而与补充能量储存以维持能量平衡无关。 在抗阻训练及其合成代谢特征的背景下, 男性更高的能量摄入不应被看作为传统的补偿机制。
有氧运动是以次最大强度进行的连续运动,涉及大量骨骼肌参与。 有氧运动因为可以快速引起能量负平衡,一直被用来对抗肥胖。 尽管如此,有氧运动可以准确地影响能量补偿的大小, 因此它对体重减轻的效果是有争议且具有个体差异的, 如有些人会出现快速的体重减轻, 而另一些人实际上会增加体重[110-111]。一些研究表明,急性有氧运动对即刻的能量摄入几乎没有影响[112-113],而其他研究表明,有氧运动可以影响饥饿激素并改变肌肉和肝脏中的底物氧化,这可能与运动后减少饥饿感和食物摄入相关[114-115]。
4.2 运动剂量
运动剂量是影响有氧运动干预能量补偿和体重减轻的另一个重要变量。最近一项研究表明,每周有氧运动消耗1 500 kcal 或3 000 kcal 的个体在能量补偿方面没有差异(即每周1 000 kcal),但较大的运动剂量组(即每周3 000 kcal)在干预12 周后会出现显著的体重减轻[116]。 这一发现表明,更多的有氧运动可能不会产生更多的能量补偿。相反,需要大剂量的运动来克服代偿反应,以促进脂肪的显著减少。这些发现在另一项研究中得到重复验证, 通过有氧运动的能量消耗为每周2 753 kcal 和1 490 kcal, 出现了类似的能量补偿, 但较大量的运动在体重减轻方面更有效[51]。 这些发现似乎与Rosenkilde 等[25]的研究结果不一致,后者证明在运动期间每周消耗1 800 kcal或3 600 kcal, 在12 周后产生几乎同等程度的能量负平衡和脂肪减少, 因为3 600 kcal 组产生了更大的能量补偿。 一项大型的运动诱导体重补偿机制(Examination of Mechanisms of Exercise-Induced Weight Compensation, E-MECHANIC)的研究提供了额外的证据,即大运动量组(运动能量消耗为20 kcal/kg体重)比低运动量组(8 kcal/kg 体重)补偿更多[117]。然而与8 kcal/kg 组相比,20 kcal/kg 组的体重减轻也更多(即分别为-1.6 和-0.4,P<0.05)。 这些结果表明,减轻体重需要更大的运动量, 但同时也会引起更大的能量补偿。 综上所述,能量补偿与运动剂量呈正相关。然而, 这种补偿是否会在大运动量下达到平台仍有待确定。 未来的研究可能会受益于评估对更大运动剂量的锻炼干预的补偿反应,即每周4 000~5 000 kcal。
5 结论与展望
参与的活动越多,当日结束时消耗的能量就越多,这个概念被形式化为能量消耗的加法模型, 如图3(左)所示,该模型已应用于人类和其他动物中[118-119]。在“加性总能量消耗”模型中,总能量消耗是身体活动的简单线性函数, 身体活动能量消耗的变化决定了总能量消耗的变化。在“约束总能量消耗”模型中,身体通过减少其他能量过程来适应增加的身体活动,从而将总能量消耗保持在狭窄的范围内。 然而,在对人体层面进行干预后成功减轻体重的结果有限, 这表明某些潜在发生的补偿机制抵消了运动或热量限制引起的能量负平衡。 目前已积累了关于人类和其他动物的能量补偿的明确证据, 表明能量补偿可以是相当可观的,约600 kcal/d[51,116,120]。 简单地说, 一旦通过运动或节食创造了负能量平衡,RMR或NEAT 也会随之降低, 从而使每日总能量消耗保持在合理范围内。 各种能量过程中的相互变化的能量约束模型如图3(右)所示。 能量平衡调节有利于防御能量缺失,而非盈余。 可以说,长时间的能量不足受到感觉反馈系统的保护, 该系统的功能是防止饥饿、促进生存和物种延续。在人类存在的大部分时间里,这是一个确保生存的工具系统。 然而,在食物充裕、方便、能量密集和生活方式多为久坐不动的现代, 曾经必要而刚性的补偿机制正在肥胖趋势上升中发挥作用。
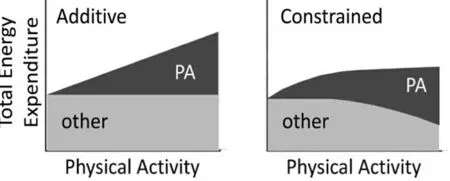
图3 “加性总能量支出”和“约束总能量支出”模型示意图Figure3 Schematic of “Additive Total Energy Expenditure” and “Constrained Total Energy Expenditure” models
尽管已认识到这一重要现象, 就如何实现能量补偿以及它如何破坏减肥努力而言, 结果仍然没有定论。仍有许多重要而有趣的问题需要研究,包括每日能量消耗的上限是否会随体重、 年龄和性别等因素而变化,以及如何变化,活动类型和强度如何影响能量补偿的程度和性质[121-122],以及当能量负平衡是由增加体力活动或减少饮食摄入引起时, 能量补偿是否更明显[123]。 未来的研究需要是长期的,并且要能够准确和直接测量参与者在减重干预中体重和体成分变化较小的情况下的全部能量变化过程, 包括RMR、NEAT 和重度体力活动。行为改变是否受主观影响还有待确定(即,“我可以吃得更多,因为我锻炼了”,“我已经锻炼了,所以我不需要在一天的剩余时间里进行或不进行身体活动”)[65]。最后,解决能量补偿的奥秘可能需要识别对运动干预的生理和行为反应的实质性表型变化[65,124-125]。 例如,有些人可能会变得不那么活跃, 有些人可能会表现出较低的RMR波动,有些人可能会表现出更高的线粒体产能效率,而另一些人可能根本没有反应。
了解能量补偿的概念可能对临床运动生理学研究者、医生或其他专门从事减肥和将运动用作药物的医疗保健领域大有裨益。 本综述中提供的证据可用于优化现有减肥指南中的运动剂量和强度。 增加能量消耗是实现负能量平衡的直观方法,通常通过锻炼增加身体活动来实现。 对于有氧运动来说,如果消耗多达每周3 000 kcal,总能量补偿可以达到约1 000 kcal[51,116]。 这一发现表明为克服对新运动计划的约每周1 000 kcal 的补偿反应提供了证据,如果将减肥作为目标,能量消耗应约为每周3 000 kcal。更大的能量消耗可能会导致更大的补偿反应, 而较低的消耗将更容易得到补偿,从而阻碍体重的减轻。可能存在一种适合减重的最佳锻炼量, 这种最佳量在增加锻炼量与因伴随的能量补偿增加而带来的收益递减之间权衡。 更高强度的训练(HIIT)也是有利的,因为对这种训练类型的补偿反应没有那么大。因此,在提出减肥建议时应考虑运动强度。可以将抗阻训练作为减重的运动干预方式, 因为这种运动方式已被证明可以增加RMR 和NEAT,从而有利于维持能量负平衡。
一些心理方面干预也可以减轻代偿反应, 使运动干预在减肥中更有效。例如,一个人如何看待运动(即,害怕的、困难的、愉快的)可能会影响减肥结果。 如果在运动方式、强度、持续时间和音乐方面为人们提供结构化的选择, 人们将更倾向于认为运动是愉快的, 而不是在运动后从食物中寻求奖励[126]。当对运动方案的最初想法更积极而非消极时, 情况也是如此。如果有人认为运动会很困难,那么更有可能为完成运动任务来寻求奖励[127]。 未来针对减肥的建议不仅需要考虑运动时的能量代谢, 还需要考虑运动计划可能导致的行为反应, 以及这些反应如何影响各种补偿机制,从而影响减肥效果。
