青海湖裸鲤肠道显微结构及基于宏条形码的食性分析
2023-11-22蔡伟杰张存芳李柯懋郝佳慧覃敏欣关弘弢强刘聂苗苗贾军梅祁得林
蔡伟杰 张存芳 李柯懋 郝佳慧 覃敏欣 关弘弢 高 强刘 丹 聂苗苗 贾军梅 祁得林
(1.青海大学省部共建三江源生态与高原农牧业国家重点实验室,西宁 810016;2.青海大学生态环境工程学院,西宁 810016;3.青海省渔业技术推广中心(青海省渔业环境监测中心),西宁 810012)
青海湖裸鲤(Gymnocypris przewalskii)作为青海湖特有的土著四倍体硬骨鱼类[1],也是青海湖唯一的商业鱼类,属于国家二级保护动物,于2004年被列入《中国濒危动物红皮书》和《中国生物多样性保护行动计划》。作为青海湖特有的洄游鱼类,成年鱼每年春末和夏季会迁徙到连通河流的淡水环境中进行产卵,并在初秋和冬季又返回到青海湖[2]。青海湖裸鲤作为青海湖鸟岛及其邻近淡水河候鸟的主要食物来源,每年大约有3000吨被鸟类捕食[3],说明它在维持青海湖生物多样性以及生态系统稳定性中处于极其重要的地位。但由于其生活在高盐、强紫外、低温和食物极度匮乏的极端环境中,其生长缓慢、性成熟晚和生命周期长等特点[4],也是其资源保护的制约因素。在20世纪50年代到90年代,一度由于过度捕捞和水质污染[5]等人类活动导致野生青海湖裸鲤数量急剧下降,种质资源严重衰退,青海省人民政府于1994年正式禁止商业捕捞[6]。除此之外,湖泊盐度升高和水位下降等自然原因给青海湖裸鲤的生存也带来了严峻的挑战[7,8]。青海湖裸鲤直至今天依然是当地主要的渔业保护和发展资源,为了保护和挖掘特色资源,许多学者从生理、生化、分子遗传基础等方面进行了研究,发现其生长速度缓慢、繁殖能力低,具有耐低温、耐盐碱、耐低氧和耐贫养等特性[7,9],是研究物种进化和环境适应的理想模式生物[10,11]。
鱼类肠道由内胚层和中胚层发育而来,前端接于食道或胃,后止于肛门处,是鱼类与外界环境接触面积最大的器官,起着免疫、消化吸收和新陈代谢[12]等众多功能的重要器官,肠道功能的正常与否直接关系着鱼类整体的健康。在组织学上,大多数鱼类肠道的组织结构比较相似,许多鱼类的肠道壁组织由黏膜层、黏膜下层、肌层和浆膜层构成[13],且根据已有的研究结果,鱼类的食性[14]和年龄[15]等都是影响肠道组织结构的重要因素。除了对肠道的基本组织结构进行研究外,不同鱼类消化道黏液组织化学的研究也是相关的热点[16,17],鱼类消化道中的黏液细胞包括杯状细胞、胃肠道上皮细胞和腺体中的一些细胞[18],鱼的种类不同消化道黏膜细胞分泌的黏液蛋白分布也不相同[19],从而发挥的功能也不相同。另外,前、中和后肠的肠绒毛形态、柱状上皮细胞和杯状细胞等的分布及数量主要在黏膜层上体现出结构差异性[20]。
鱼类的肠道结构特点主要与食性相关从而直接影响鱼类的生长发育,如肉食性鱼类拥有更大的肠道直径和更发达的肠道结构能容纳和吸收更多食物[21];杂食性鱼类前肠容量不大,但黏膜皱褶高且密,极大地增加了前肠的消化功能[22];草食性鱼类肠绒毛发达,对水生植物中含量低的营养物质有极强的吸收作用[23]。随着第二代测序技术的飞速发展,DNA宏条形码(Metabarcoding)技术以成本低、通量大和能快速鉴定大量物种等优势,被广泛应用于浮游植物群落的监测[24]、浮游动物的鉴定[25]和食性分析[26]等生态学领域的众多方面,在鱼类食性鉴定[27]方面也逐渐成为了有利的工具。
无胃鱼的消化道一直是一些超微结构研究的热点,青海湖裸鲤作为典型的无胃鱼,有关其肠道结构的研究报道较早,史建全等[28]从形态学到组织结构的异同等方面对青海湖裸鲤肠道组织学进行分析,并讨论了其肠道组织学和食性之间的相关关系;陈付菊等[15,29]的研究中表示青海湖裸鲤的前肠是消化和吸收的主要功能部位,并对不同发育时期青海湖裸鲤肠道的杯状细胞和上皮内淋巴细胞做了比较分析。但经过几十年的封湖禁渔,对青海湖裸鲤的保护取得了明显的成效,其数量得到了有效提升,保护级别由濒危降为易危,随着鱼类数量的提升鱼类的生长发育又有了新的变化。因此,尽管对于青海湖裸鲤肠道结构的研究已经相对清楚,但相关的报道都比较早,缺乏较新的研究结果与之进行比较分析,在此基础上,本研究通过扫描电镜、HE染色和DNA宏条形码等技术手段来分析青海湖裸鲤肠道结构及食性的特点,进一步揭示青海湖裸鲤肠道自身生理结构的差异性和特殊性,为青海湖裸鲤的研究和保护工作提供基础的科学依据,同时也为青海湖裸鲤的人工增殖工作提供参考和新思路。
1 材料与方法
1.1 实验材料
实验鱼采自青海省海南藏族自治州共和县西北部的青海湖附属河流—黑马河(E99°46'53″,N36°43'20″),pH为8.0—8.2,海拔为3110.8 m,采样时间为7月上旬,在河道中随机捕捞20尾青海湖裸鲤,装入供氧容器,保持存活带回临时采样点进行取样,将活鱼麻醉后,进行常规的生物学测量,然后使用无菌的剪刀和镊子进行解剖,快速将肠道从其体腔中分离出来并测量长度,再选取体长体重相近的8尾鱼,收集肠道内容物于2 mL冻存管中置于液氮中用于测定食性,最后取其中3尾鱼的肠道样品用于扫描电镜和HE染色切片的制备。
1.2 肠道扫描电镜样品的制备
分别取新鲜的前、中和后肠1 cm×1 cm大小的肠道组织1—2块(前肠在第1个弯曲前取材,中肠在第1个弯曲到最后一个弯曲之间取材,后肠在最后一个弯曲到肛门之间取材),用PBS磷酸缓冲液冲洗3次,迅速固定于4℃预冷的2.5%戊二醛溶液中保存24h,再依次用浓度为30%、50%、70%、80%、95%和100%的乙醇进行梯度脱水,叔丁醇置换酒精后再次脱水,放入-80℃冰箱中冷冻4h,再置于冷冻干燥仪中干燥6—8h。在干燥完毕之后进行镀金,最后在扫描电镜下(FESEM)观察并摄片。
1.3 肠道HE染色样品的制备
制备方法如下: (1)70℃下烤片5min,置于二甲苯(Ⅰ)、二甲苯(Ⅱ)中各脱蜡10min,再用100%乙醇(Ⅰ)、100%乙醇(Ⅱ)各5min洗去二甲苯;(2)经过在不同浓度梯度(95%、85%和75%)的乙醇中水化后,置于蒸馏水中待用;(3)苏木素染色10min,流水冲洗2min,1%盐酸酒精分化2s,用流水冲洗切片15min,蒸馏水过洗1—2s,伊红染色10min;(4)根据颜色深浅用80%的乙醇分化,用乙醇(85%和95%)脱水5min,再用无水乙醇脱水(10min,两次);(5)二甲苯(Ⅰ)透明10min,二甲苯(Ⅱ)再次透明10min;(6)在石蜡切片中央滴加1滴中性树胶,盖上盖玻片封固,显微镜下观察并拍照。
1.4 肠道内容物的DNA提取和PCR扩增
按照DNeasy®PowerSoil®Pro Kit Handbook DNA提取试剂盒的步骤对青海湖裸鲤肠道内容物的总DNA进行提取,选择ITS、18S rDNA、cox-1、18SV9、rbcL、ITS和COI等7个基因片段进行食性分析(表1),引物筛选只有18S rDNA和rbcL引物扩增得到了比较好的产物。PCR扩增体系为: 5×TransStart Fast-Pfu缓冲液4 μL,2.5 mmol/L dNTPs 2 μL,上下游引物(5 μmol/L)各0.8 μL,TransStart FastPfu DNA聚合酶0.4 μL,模板DNA10 ng,ddH2O补足至20 μL。PCR扩增程序为: 95℃预变性3min;95℃变性30s,55℃退火30s,72℃延伸30s,设定27个循环;72℃延伸10min后4℃保存。

表1 标记基因及引物序列Tab.1 Marker gene and primer sequence
1.5 Illumina Miseq测序及生信分析
2%琼脂糖凝胶回收PCR扩增产物,用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City,CA,USA)试剂盒进行回收产物纯化,并用Quantus Fluorometer(Promega,USA)对回收产物进行检测定量。使用TruSeqIM DNA Sample Prep Kit进行建库,并在Illumina的Miseq PE300平台上进行测序。所得测序结果进行序列拼接、质量控制和过滤,在Uparse7.0(http://drive5.com/uparse/)软件上进行OTU聚类分析,采用RDP Classifier(http://sourceforge.net/projects/rdp-classifier/)贝叶斯算法对97%相似水平的OTU代表序列进行基于Silva(http://www.arb-silva.de)、Unite(http://unite.ut.ee/index.php)数据库的分类学分析,并分别在Phylum(门)、Class(纲)、Order(目)、Family(科)、Genus(属)和Species(种)六个分类学水平下统计各样本的群落物种组成。以上分析过程在美吉生物云平台(https://cloud.majorbio.com)上完成。
2 结果
2.1 体重、体长和肠长之间的变化关系
对20尾繁殖期性成熟的青海湖裸鲤的体重、体长及肠长进行了测量,并按照体长大小对测量结果进行统计分析(表2),结果显示各相邻体长阶段之间裸鲤的平均体重、头长和体高差异均不显著(P>0.05),而在体长差异相对较大的两个不相邻阶段之间均体现出了显著差异性(P<0.05),且体长为20—21 cm的裸鲤与18—19 cm的裸鲤相比其肠长差异较显著(P<0.05)。根据青海湖裸鲤体重、体长和肠长的实测数据,用幂函数模型,拟合出体重-肠长的关系(WGR)、体长-肠长(BGR)的关系分别为:
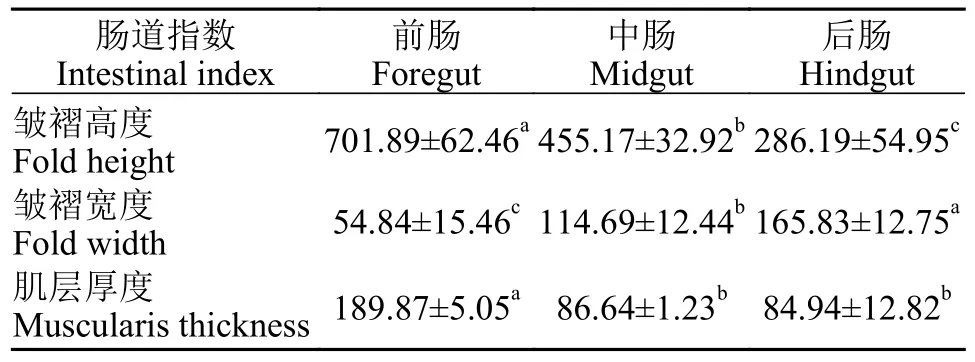
表3 青海湖裸鲤肠道皱褶高度、宽度和肌层厚度Tab.3 Fold height,fold width and Muscularis thickness in G.przewalskii (μm)

表4 基于18S rDNA和rbcL宏条形码分析的青海湖裸鲤主要食物组成Tab.4 Main food composition of G.przewalskii based on 18S rDNA and rbcL macro barcode analysis
上述两个方程存在显著差异,但均体现出了体重与体长、肠长之间的拟合关系,且根据所测20组数据计算均值可以粗略得出,体重在32—68 g、体长在18—22 cm内的青海湖裸鲤其体重和肠长是1.4倍的变化关系,而肠长约为体长的1.7倍。
2.2 肠道扫描电镜结果分析
对不同放大倍数下青海湖裸鲤各肠段扫描电镜的结果进行分析,总体来说,前肠黏膜皱褶(肠绒毛)明显且发达,数量最多,褶层很深宽度较窄呈波浪状排列,大皱褶中又包含了许多小皱褶,增加了皱褶的总长度和表面积,褶与褶之间相互交联形成较无序的网状排布(图1a),前肠黏膜层由数量众多的单层柱状上皮细胞构成,且黏膜层表面可见大量的分泌孔、分泌颗粒及杯状细胞散布其间(图1b和1c)。中肠和后肠黏膜表层仍由单层柱状上皮细胞构成,但相比于前肠,中肠和后肠的肠道黏膜皱褶深度逐渐变浅,宽度逐渐增加,皱褶分支的数量相对减少,但皱褶之间排列相对较整齐,呈较规律的“V”“M”等字型排列(图1d和1g)。中肠和后肠黏膜表面的杯状细胞的数量较前肠相对减少,分泌孔及周围黏附分泌物颗粒的数量也逐渐减少,其中后肠的数量最少(图1b、1e和1h),但后肠黏膜表面的微绒毛的密度和数量较多,高度相近且排列整齐,尤其是肠黏膜上的呈六边形分布的短簇状微绒毛,细胞之间的界限也很清晰(图1h和1i),这些肠道黏膜皱褶和微绒毛都极大地增加了肠道的内表面积,能促进对食物的消化和吸收。

图1 青海湖裸鲤肠道扫描电镜结果Fig.1 Results of intestinal SEM of G.przewalskii
2.3 肠道HE染色结果分析
肠道组织经HE染色处理后在光镜下进行观察,可见前肠肠绒毛顶部呈尖圆状,排列最为紧密(图2a),中肠肠绒毛纹状缘清晰,绒毛之间的排列逐渐稀疏(图2b),后肠肠绒毛顶部呈钝圆状,纹状缘较模糊,数量最少排列较整齐(图2c),从前肠到中肠再到后肠皱褶高度显著减小(P<0.05),皱褶宽度显著增加(P<0.05),后肠肌层厚度最小与中肠差异不显著(P>0.05),前肠肌层厚度最大与中肠和后肠相比差异均显著(P<0.05;表 3),且从前肠到后肠肠腔逐渐变小,肠道直径也逐渐变小;青海湖裸鲤的肠道组织经HE染色处理后,质核区分显著(图2d),肠道组织边缘清晰且结构完整,肠壁组织结构层次分明,从内向外依次由黏膜层、黏膜下层、肌层和浆膜层四层组成,但因其缺乏黏膜层中的黏膜肌致使固有膜与黏膜下层间的区分不明显,故光镜下只有三层膜结构清晰可见(图2e),前肠黏膜上皮密布柱状上皮细胞,并有许多杯状细胞散布其间,而中肠至后肠黏膜层向肠腔内褶形成的皱褶高度较前肠逐渐减小,且柱状细胞的数量逐渐减少(图2f),故前、中和后肠的黏膜层结构差异较为明显(P<0.05),而这三个肠段的黏膜下层、肌层和浆膜层结构基本相似,并无明显的结构差异。

图2 青海湖裸鲤肠道HE染色结果Fig.2 HE intestinal staining results of G.przewalskii
2.4 DNA宏条形码食性结果分析
对青海湖裸鲤8份肠道内容物样本进行18S rDNA和rbcL序列高通量测序,共获得443024条有效序列,有效序列平均长度为261 bp,剔除青海湖裸鲤自身序列、细菌、古菌及真菌的序列后,按照97%相似水平对所有序列进行OTU划分及物种注释,共得到39031个OTU,分为43个门、114个纲、282个目、435个科、599个属、817个种。图 3分别为门、纲、目、科、属、种水平相对丰度前十的物种,其他物种则合并为Others项,在门水平上主要以链型植物门(Streptophyta,53.73%)、硅藻门(Bacillariophyta,23.19%)、丝足虫门(Cercozoa,6.67%)、轮虫动物门(Rotifera,0.67%)和绿藻门(Chlorophyta,0.67%)等为主(图3a);在纲水平上主要以真藓纲(Bryopsida,35.79%)、硅藻纲(Bacillariophyceae,23.05%)、双子叶植物纲(Magnoliopsida,18.03%)、金藻纲(Chrysophyceae,3.36%)和Bdelloidea(0.45%)等为主(图3b);在目水平上主要以桥弯藻目(Cymbellales,22.60%)、灰藓目(Hypnales,17.97%)、蔷薇目(Rosales,17.97%)、丛藓目(Pottiales,17.92%)和Glissomonadida(3.39%)等为主(图3c);在科水平上主要以蔷薇科(Rosaceae,18.03%)、丛藓科(Pottiaceae,17.99%)、柳叶藓科(Amblystegiaceae,17.63%)、金囊藻科(Chrysocapsaceae,2.91%)和Heteromitidae(0.57%)等为主(图3d);在属水平上主要以丛藓属(Pottia,18.03%)、中国毛莓草属(Sibbaldianthe,17.86%)、拟细湿藓属(Campyliadelphus,17.68%)、Chrysochaete(2.93%)和Heteromita(0.57%)等为主;在种水平上主要以Cymbellalessp.TF-2014(22.61%)、Pottia intermedia(18.02%)、Sibbaldianthe sericea(17.85%)、Campyliadelphus chrysophyllus(17.67%)和Chrysochaete britannica(2.93%)等为主。

图3 丰度前十的各水平生物分类单元Fig.3 Taxa of the top ten levels of abundance
为进一步探讨丰度占比较大的各食物类群之间的从属关系,我们对几个优势门下属的各分类水平的食物组成又做了详细分析,DNA宏条形码食性测序结果显示门水平上丰度前四的食物组成,在纲、目、科和属水平上占比较大的物种与前面所述结果一致,四个优势门包含着丰富的食物类群,且主要以链型植物门和硅藻门种类居多、占比较大(图4),其中链型植物门中主要包括真藓纲、双子叶植物纲;真藓纲主要包括灰藓目和丛藓目,双子叶植物纲主要包括蔷薇目和伞形目;灰藓目主要包括柳叶藓科和绢藓科,丛藓目包括丛藓科,蔷薇目包括蔷薇科,伞形目主要包括伞形科和五加科;柳叶藓科主要包括拟细湿藓属和牛角藓属,绢藓科包括绢藓属,丛藓科主要包括丛藓属和花栉藓属,蔷薇科主要包括中国毛莓草属和沼委陵菜属,伞形科包括当归属,五加科包括人参属;拟细湿藓属包括Campyliadelphus chrysophyllus,牛角藓属包括牛角藓,绢藓属主要包括广叶绢藓和Entodon rubicundus,丛藓属包括Pottia intermedia,花栉藓属包括Pseudocrossidium replicatum,中国毛莓草属包括Sibbaldianthe sericea,沼委陵菜属包括西北沼委陵菜,当归属和人参属主要是当归和人参。硅藻门主要包括硅藻纲和圆筛藻纲;硅藻纲主要包括桥弯藻目和硅藻目,圆筛藻纲主要包括角毛藻目和Thalassiosirales;桥弯藻目主要包括桥弯藻科和f_unclassified_o_Cymbellales,硅藻目包括杆状藻科,角毛藻目主要包括角毛藻科,Thalassiosirales主要包括Stephanodiscaceae和Skeletonemataceae;桥弯藻科主要包括内丝藻属和桥弯藻属,f_unclassified_o_Cymbellales包括g_unclassified_o_Cymbellales,杆状藻科主要包括Denticula和Nitzschia,角毛藻科包括角刺藻属,Stephanodiscaceae包括Discostella,Skeletonemataceae包括骨条藻属;内丝藻属主要包括西里西亚内丝藻和默利内丝藻,桥弯藻属包括近缘桥弯藻,g_unclassified_o_Cymbellales包括Cymbellalessp.TF-2014,Denticula包括库津细齿藻,Nitzschia主要包括两栖菱形藻和Nitzschia pusilla,角刺藻属主要包括Chaetoceros peruvianus,Discostella主要包括Discostellasp.HYK0210-A2和Discostella stelligera,骨条藻属主要包括骨条藻(图4和表 4;其中“unclassified”为无分类信息的标记)。

图4 四个优势门所包含纲、目、科和属水平主要组成的桑基图Fig.4 The four dominant phyla consist of class,order,family and genus level mainly composed of Sankey chart
3 讨论
3.1 青海湖裸鲤肠长与体长、体重之间的关系
青海湖裸鲤属于鲤科裂腹鱼亚科的杂食性鱼类,食性范围广,无胃,食道之后便是肠道,从形态学方面,其肠道包裹在腹膜中盘旋复杂,肠道长度介于肉食性鱼类和草食性鱼类之间,吴强等[30]的研究也表明肉食性鱼类肠道最短,草食性鱼类肠道最长,杂食性鱼类则介于中间。青海湖裸鲤肠道的长度除了和食性有关之外,还和其体重、体长等相关,朱奕龙[31]在青海裸鲤生长特征的研究中,通过拟合体重和体长的关系,也发现雌雄性鱼的体长与体重都接近等速生长。而本研究的统计结果显示,体长相近的各生长阶段之间,体重变化也较相近,且通过分别拟合体重和肠长、体长和肠长之间的相互关系,结果表明青海湖裸鲤不仅在体重和体长之间存在相应的变化关系,肠道长度也会随着体重的不同、体长的变化而变化,并表现出一定的正向倍数变化关系,而体长与肠长的变化关系会有较大的差异性,王基琳[32]在青海湖裸鲤食物的研究中也表明其体长与肠长之比变化幅度较大。
3.2 青海湖裸鲤肠道组织特点与功能之间的关系
以肠道上两个弯曲部分的位置加以区分,可以将青海湖裸鲤的肠道划分为前、中和后三部分肠段,前肠长度最短肠腔最大,扫描电镜下可见肠道黏膜皱褶数量最多且凸起高度最大,肠壁内表面积最大,黏膜表面分泌孔及分泌物颗粒数量最多,而中肠和后肠部分占比较长,肠腔逐渐变小,肠绒毛逐渐变短变粗,这与其他鱼类相似,主要是因为前肠承担着大多数的消化吸收功能,其次是中肠和后肠。房慧伶等[33]对杂食性尼罗罗非鱼(Oreochromis niloticus)肠道组织结构的研究表明,前肠和中肠的柱状细胞顶端有密而长的微绒毛,后肠柱状细胞顶端的微绒毛稀而短,并且前、中和后肠的组织结构差异也主要表现在黏膜层;范瑞青等[34]在对蓝非鲫(Oreochromis aureusSteindachner)肠道结构的研究中表明前肠是消化和吸收的主要功能区,中肠和后肠的微绒毛不及前肠发达,主要起对碳水化合物及剩余食物的吸收、消化和运输的作用;何敏等[35]在重口裂腹鱼[Schizothorax(Racoma)davidi]的肠道组织结构研究中发现,从前肠至后肠其肠道肠壁内黏膜褶高度和黏膜上皮高度逐渐递减,肠道消化吸收能力逐渐降低,再一次表明鱼类消化道形态学组织结构与生理功能之间的密切关系。
HE染色处理下青海湖裸鲤的肠道结构组织,与其他大多数鱼类相似,由内向外依次由黏膜层、黏膜下层、肌层和浆膜层构成,前、中和后肠肠道结构基本相似,前肠肠壁和肌层都较中肠和后肠厚,可以提供更多的肠道蠕动力,促进消化和剩余食团向后继续的运送。黏膜层的差异主要体现在肠绒毛的排列、数量、长短、宽窄及前肠黏膜上皮分布的更多数量的柱状细胞和杯状细胞等方面,柱状细胞的数量与肠道的吸收功能直接相关,杯状细胞除了能分泌消化酶促进对食物的消化以外,还能分泌肠道内的黏蛋白,在抵御外源细菌和肠道固有微生物侵袭、维持肠黏膜动态平衡、调控微生物-宿主免疫应答中起重要作用[36]。这两类细胞都极大地增加了肠道对食物的消化和吸收功能,中肠和后肠黏膜上皮分布的柱状细胞和杯状细胞数量则相对减少。除此之外,陈付菊等[15]对不同年龄青海湖裸鲤的肠道组织结构进行了比较分析,研究表明肠道中绒毛长度、黏膜厚度、肌层厚度和肠壁厚度还会随着年龄的增长而逐渐增大。
3.3 青海湖裸鲤的食性特点及相对变化
利用DNA宏条形码技术对青海湖裸鲤的肠道内容物进行检测,依据检测结果,按照物种序列的门、纲、目、科、属、种及所占比例,可以初步判断本次研究采集到的青海湖裸鲤其采食食物中种类最丰富的有链型植物、硅藻、丝足虫类、轮虫类及目前还未被分类但占比较大的一些物种等。相比传统形态学鉴定其食性的研究方式[37],硅藻、绿藻、摇蚊幼虫、钩虾、轮虫类和介形类等均也在本次研究中被检出,而蓝藻、裸藻、枝角类和桡足类等未被检测出。另外本研究还检测到红藻、金藻、黄藻、脊索动物、节肢动物、软体动物、扁形动物、环节动物、缓步动物、变形虫类、纤毛虫类和眼虫类等传统镜检法未检测到的类群。从检出食物的丰度上,链型植物和硅藻的数量最多丰度最大,丝足虫类和轮虫类等也在其各分类学水平上较传统镜检法检出食物的类别和数量更加系统和全面。此外,本研究检测到占比较大的链型植物(53.73%)在之前的研究中未见详细报道,季强[22]在拉萨裸裂尻鱼、巨须裂腹鱼和拉萨裂腹鱼的肠道内含物中均发现了高等植物碎片及种子,我们推测链型植物也可能是青海湖裸鲤的主要食物类别之一。在本研究中植物性食物占比为77.59%,因此我们推测青海湖裸鲤为杂食性偏植食的鱼类。而对于未检测到的食物及食物组成占比大小的问题,对比前面所参考的研究及有关文献[32],我们推测这可能与青海湖不同月份食物组成及所检测鱼的数量相关,也可能是由于气候变化及湖泊水环境质量的变化导致青海湖中浮游植物、浮游动物、底栖动物种类及丰度发生了变化[38,39],从而使得青海湖裸鲤的食性也发生了变化。除此之外,结合青海湖裸鲤肠道内容物DNA宏条形码食性分析的结果,可以在一定程度上对作为杂食性鱼类的青海湖裸鲤肠道组织结构的特异性做出进一步的解释,同时也能说明鱼类食性与其肠道结构之间存在相互适应关系。
青海湖裸鲤野生种群数量较少,生长缓慢,繁殖率低,对青海湖裸鲤肠道显微结构形态学和组织学、生长特性、食性的研究,在青藏高原鱼类适应极端环境的研究及阐明鱼类食性与其肠道结构之间的密切关系中具有重要的价值和地位,填补了利用高通量测序技术探究其食物类别的空缺,提供更新的数据参考,同时给青海湖裸鲤的人工增殖工作提供一定的科学基础。
