早期阶段饥饿对鲢仔鱼存活和生长的影响
2023-11-22朱峰跃杨锦毅雷汪登强段辛斌陈大庆
王 敏 朱峰跃 杨 浩 蒲 艳 杨锦毅,3 高 雷汪登强 段辛斌 陈大庆
(1.西南大学生命科学学院,淡水鱼类资源与生殖发育教育部重点实验室,重庆 400715;2.中国水产科学研究院长江水产研究所,武汉 430223;3.上海海洋大学水产与生命学院,上海 201306)
仔鱼期是所有鱼类早期生活史阶段的一个关键时期,在此期间,许多生物和非生物因素均会影响仔鱼的生长和存活,进而对仔鱼年际强度和种群补充量产生显著影响[1,2]。其中,仔鱼从内源营养转入外源性营养的初次摄食期,对其随后的生长和存活至关重要[3],且饥饿被认为是仔鱼在此“临界期”死亡率高的主要影响因素之一[4]。Cushing[5]在“临界期假说”的基础上发展了“匹配-不匹配假说”,认为仔鱼的存活和补充取决于仔鱼在时间和空间上与适口浮游生物饵料的匹配程度。然而,大多数鱼类在仔鱼早期游泳能力较差,其野外分布主要取决于水流,漂流的仔鱼不一定会被输送到有充足浮游生物饵料的地区[6]。如果在开口后不久未能成功开始初次摄食,大多数仔鱼会出现形态畸形,甚至导致无法游泳和搜索食物[7]。为了测定仔鱼的耐饥饿能力,Blaxter等[8]首次提出了不可逆点(The pointof-no-return,PNR)的概念,即饥饿仔鱼到达该时间点时,50%的仔鱼已虚弱得不能恢复摄食能力,最终进入生态死亡阶段。
鲢(Hypophthalmichthys molitrix)隶属于鲤形目(Cyprinidformes)、鲤科(Cyprinidae)、鲢属(Hypophthalmichthys),是我国特有的重要淡水经济鱼类,长江中游是其重要的繁育场所[9]。鲢为江河洄游性鱼类,在河道中产漂流性卵,卵随水流漂流孵化,孵出的仔鱼被水流冲入河漫滩或泛滥平原湖泊中育幼。仔鱼到达河漫滩后,通常会开始主动摄食和发育[10,11],但是低流速和较差的河道-漫滩连通性意味着鲢仔鱼在向下游漂流过程中将经历较长的饥饿期[12]。近年来,包括形态测量学[13]、组织学[14]、消化酶活性[15,16]和游泳能力[17]等诸多方法已被用来研究饥饿对仔鱼存活和生长的影响,我国曾有学者对鲢仔鱼开口摄食时间、开口大小和耐饥饿能力进行了初步研究[18—20],而针对延迟投喂对鲢仔鱼生长和存活的影响研究未见报道。鉴于此,本研究通过不同程度的饥饿胁迫对鲢仔鱼的摄食、生长和存活的影响实验,旨在探讨鲢在早期发育阶段面临饥饿时的生态适应策略,对解析其早期补充过程、资源衰退机制和管理对策具有重要意义。
1 材料与方法
1.1 实验材料
初孵仔鱼来源2021年5月上旬,在湖北石首老河长江“四大家鱼”原种场对鲢亲鱼进行人工催产,自然产卵受精获得受精卵,在室内水池中流水孵化获得仔鱼。选取同一批仔鱼样本,装入充氧塑料袋中,随即转移到实验室。
仔鱼培育将鱼苗刚孵化出膜当天记为1日龄,根据林浩然[21]和预备实验的观测结果,鲢仔鱼在本实验条件下3日龄开始陆续摄食。因此孵化后第2天,将初孵仔鱼按照分组分别放于25 L玻璃水族缸(0.40 m×0.25 m×0.25 m)中饲养,缸内保持水量为20 L,饲养密度为25尾/L。实验养殖用水为先沉淀曝气,再经双层250目筛绢网过滤的天然河水,确保水体中没有鲢仔鱼能摄入的浮游生物饵料。培育水温保持在(24±1)℃,光周期设置为13L∶11D,微充气,溶解氧>7 mg/L。每日换水1次,换水量为总量的1/3,通过虹吸法早晚2次从玻璃缸底部移出饵料残渣和死亡的仔鱼。饥饿处理组仔鱼不予投饵,摄食组仔鱼每天9:00和18:00定时投饵。采取饱食投喂的方式,在仔鱼3—10日龄的实验前期投喂熟蛋黄水悬浊液,后期11—30日龄投喂人工孵化的丰年虫无节幼体。
1.2 实验方法
延迟投饵实验设计本实验共设5种延迟投饵水平组,每组3个重复,每个重复组喂养500尾仔鱼。前4个处理首次摄食的延迟时间分别为0、2d、4d和6d,即在3日龄、5日龄、7日龄和9日龄对仔鱼提供首次投喂,对应T0(对照摄食组)、T2、T4和T6组。第5个处理组为完全饥饿组FG(Fasted group),实验期间对仔鱼一直不投喂,直至全部死亡。实验设计概要如图1所示。

图1 延迟首次投喂的实验方案概要Fig.1 Summary of the experimental schedule of delayed initial feeding
在实验开始后,从1—14日龄每天从各个实验组中取24尾仔鱼(每个重复组随机取8尾),14日龄之后隔日抽样1次,直到30日龄实验结束。仔鱼样本用4%福尔马林溶液固定,使用与计算机相连的OLYMPLUS SZX16型解剖镜对鲢仔鱼进行拍照,用cell-Sens Entry图像处理软件测量全长、体长、卵黄囊长径和短径(精度为0.01 mm)。拍完照后的仔鱼用滤纸拭去体表水分,在电子天平(精度为0.1 mg)上测定体质量。特定生长率(Specific growth rate,SGR,%/d)和卵黄囊体积(V,mm3)的计算公式如下:
式中,SGRW为体质量特定生长率,W2和W1分别代表t2和t1日龄时的体质量;SGRL为全长特定生长率,L2和L1分别代表t2和t1日龄时的全长;r为卵黄囊短径(mm),R为卵黄囊长径(mm)。
用差异系数(Coefficient of variation,CV,%)研究延迟首次投喂对仔鱼个体生长差异的影响。CV=,式中,SD为标准差,为t日龄时的全长平均值。
形态性状测量为了通过形态测量法揭示饥饿对仔鱼形态性状的影响,在3—14日龄每天记录饥饿组和摄食组仔鱼发育阶段(卵黄囊期、弯曲前期和弯曲期),并在10日龄用cellSens Entry软件测量全长、体长、肛前长、头长、头高、眼径和肌节高7个可量性状参数,所有形态特征均沿平行或垂直于鱼体水平轴线进行测量(图2)。为校正鱼体大小差异对形态特征值的影响,将特征值转化为比值参数,取头长/体长(HL/SL)、头高/体长(HD/SL)、肛前长/体长(AL/SL)、肌节高/体长(MD/SL)、眼径/体长(ED/SL)、头高/头长(HD/HL)、眼径/头长(ED/HL)、肌节高/头高(MD/HD)共8个形态比值参数。对2组共48尾仔鱼的形态比值参数进行主成分分析,以确认饥饿组和摄食组仔鱼组间形态变异程度和主要影响变量。

图2 鲢仔鱼形态特征测量Fig.2 Morphological characteristics measured in silver carp larvae
初次摄食率和PNR确定为了确定鲢仔鱼到达PNR所需要的时间,在进行延迟投饵实验的同时,另外设置1个饥饿组,3个重复,每个重复500尾初孵仔鱼。饥饿组仔鱼全部死亡时实验结束。从3日龄开始,每天8:30从3个缸中各取10尾仔鱼放入1 L烧杯,投喂经100目筛绢网过滤后的熟蛋黄水悬浊液。2h后将仔鱼取出,在解剖镜下逐尾检查仔鱼摄食情况,记录摄食的仔鱼数量,计算初次摄食率。初次摄食率和不可逆点的确定参照Blaxter等[8]和殷名称[22]的方法确定。初次摄食率指肠道内含有饵料的仔鱼尾数占测定总尾数的百分数。PNR以孵化后的日龄表示,当测定的初次摄食率降为最高初次摄食率的一半时,即达到PNR的时间。
仔鱼存活实验本实验与上述延迟投喂实验同步进行,但在实验期间不取样,旨在准确评估不同初始投喂处理对仔鱼存活率的影响。饲养条件和投喂策略与延迟投喂实验方案中所述一致,共设5个处理组,分别为初次摄食开始于3日龄、5日龄、7日龄、9日龄和完全饥饿组,每组3个重复,实验持续30d。鲢仔鱼在25 L玻璃缸中以20尾/L的比例饲养,缸内保持水量为20 L,每缸饲养400尾仔鱼。在实验开始后,每天记录并移除各缸中死亡仔鱼数目,进而计算各缸中仔鱼存活率(存活数/初始仔鱼数量)。
1.3 数据分析
使用IBM SPSS Statistics 26和GraphPad Prism 9软件对实验数据进行统计分析并绘图。用单因素方差分析方法(One-way ANOVA)检验不同延迟投喂水平对仔鱼形态参数、体质量、特定生长率和存活率的差异显著性,对差异显著的参数(P<0.05)用Tukey氏多重比较检验组间差异。实验数据均表示为样本平均值±标准差(mean±SD),以P<0.05作为显著性水平。
2 结果
2.1 初次摄食率和PNR的确定
初孵鲢仔鱼全长为(6.95±0.21) mm,卵黄囊体积为(1.29±0.34) mm3,1—5日龄卵黄囊体积随着日龄增加而被迅速吸收。摄食组在3日龄开始摄食后,进入混合营养期阶段,此时卵黄囊的体积与饥饿组差异显著(P<0.05)。摄食组仔鱼卵黄囊吸收速度快于饥饿组仔鱼,5日龄时摄食组卵黄囊已消耗完毕,开始转入外源性营养期,混合营养期约3d(表1)。
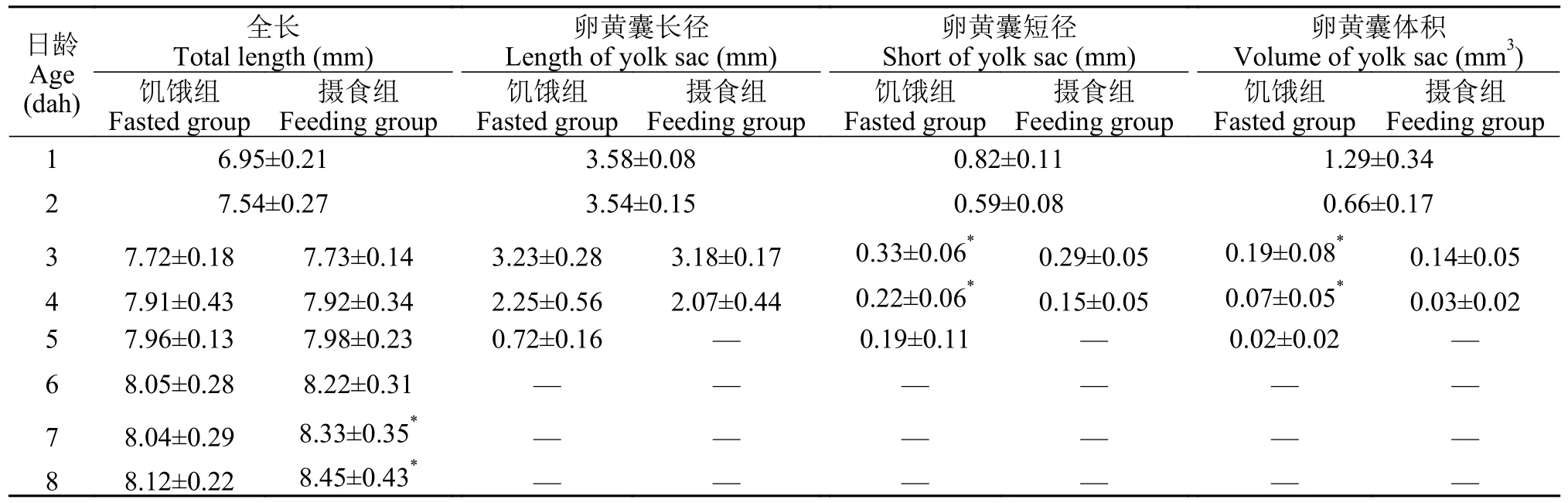
表1 饥饿组和摄食组鲢仔鱼全长和卵黄囊的变化Tab.1 Changes of total length and size of yolk sac of silver carp larvae in the fasted group and feeding group
如图3所示,在水温(24±1)℃条件下,仔鱼出膜后3日龄开口摄食,初次摄食率仅为36.67%,此后逐步增加;5—8日龄初次摄食率均在90%以上,初次摄食率在6日龄达到最高值,为96.67%;10日龄,初次摄食率下降到60%,11日龄为46.67%;12日龄,仔鱼摄食能力骤减,大部分个体已死亡;15日龄初次摄食率为0,仔鱼全部死亡。根据PNR定义,饥饿鲢仔鱼的初次摄食率下降到最高初次摄食率的一半左右的时间发生在11日龄(46.67%),即为饥饿不可逆点的时间。可见,鲢仔鱼具有摄食能力的时间约8d。

图3 饥饿鲢仔鱼初次摄食率和存活率的变化Fig.3 Changes in initial feeding rate and survival rate during the period of starvation of early development of silver carp larvae
2.2 延迟投喂对鲢仔鱼存活率的影响
从图 3可以看出,鲢仔鱼在完全饥饿条件下,50%的仔鱼死亡发生在10—11日龄,在抵达PNR时的存活率仅为41.33%,初次摄食率的显著降低伴随着存活率的急剧下降,直至15日龄仔鱼全部死亡。而在相同饲养条件下,正常投喂T0组在11日龄(初次摄食后8d)的存活率达到72.83%,在15日龄(初次摄食后12d)的存活率达到69%。随着延迟投饵时间的延长,仔鱼的存活率明显下降,T4组在15日龄(初次摄食后8d)的存活率为63.50%,T6组在15日龄(初次摄食后6d)的存活率为52.83%(图4)。
T0组和T2组的存活率在初次摄食后随时间的变化差异不大,初次摄食后2d的存活率均保持在87%左右,初次摄食后20d的存活率分别为62.58%和57.67%。延长初次投喂期,会导致首次摄食的仔鱼死亡率增加,当延迟4d和6d开始初次投喂时,摄食后2d的存活率明显低于T0组和T2组的存活率,分别为67.42%和64.08%;初次摄食后20d,T4组的存活率略低于正常投喂T0组和T2组,为52.33%;T6组在初次摄食后存活率下降明显,摄食20d后存活率仅为25%(图4)。
2.3 饥饿对鲢仔鱼形态特征的影响
在正常投喂下,鲢仔鱼的体长随日龄增长而不断增加,而饥饿组仔鱼体长无明显变化。体长在8日龄之前饥饿组和摄食组之间不存在显著性差异(P>0.05),从8日龄起摄食组显著高于饥饿组(P<0.05),10—14日龄摄食组极显著高于饥饿组(P<0.0001),16日龄饥饿组仔鱼已全部死亡(图5A)。摄食组仔鱼从8日龄开始出现脊索弯曲,14日龄全部转入脊索弯曲期,其对应的仔鱼体长分别为(7.91±0.19)和(10.08±0.56) mm(图5B和图 6A)。饥饿组仔鱼在14日龄前一直处于弯曲前期,14日龄首次出现脊索弯曲,此时弯曲期的相对频率仅占4.17%(图6A)。

图6 摄食组和饥饿组鲢仔鱼弯曲期的相对频率(A)和形态特征主成分分析图(B)Fig.6 Relative frequency of flexion (A) and principal component analysis plot of morphometric parameters (B) in fed and unfed silver carp larvae
对各样本的标准化形态比值参数进行主成分分析,结果表明前3个主成分对群体总变异的方差贡献率较大,分别为52.37%、17.84%和12.06%,累计贡献率为82.27% (表2)。将载荷值大于0.7的变量确定为主要影响变量,主成分1的肌节高/体长、头高/体长、头长/体长、头高/头长是影响两组仔鱼形态差异的主要性状,主要反映头部形态和肌节高度的变化;主成分2的眼径/头长、眼径/体长是影响两组仔鱼形态差异的主要性状,主要反映眼径的形态变化。为进一步分析仔鱼形态变异程度,绘制主成分1和主成分2的得分散点图(图6B),结果表明,饥饿和摄食鲢仔鱼在主成分1轴上的分值点分离明显,而在主成分2轴上,2个群体分布区域有较多重叠,难以区分。
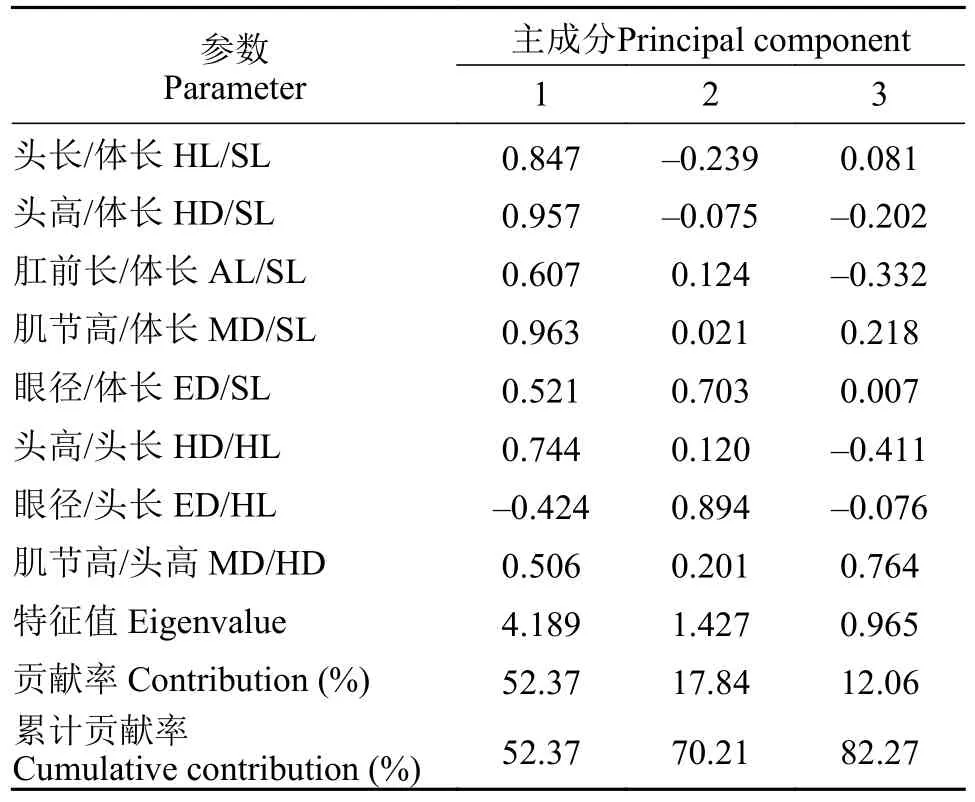
表2 形态比值参数数据的载荷矩阵及主成分对总变异的贡献率Tab.2 Loading matrix of morphological ratio parameter data and contribution rate of principal components to total variation
2.4 延迟投喂对鲢仔鱼生长的影响
在饥饿期间,随着首次投喂时间的延长,不同处理组鲢仔鱼全长SGRL和体质量SGRW呈总体下降趋势;各处理组的SGRL具有显著差异(P<0.05),Tukey氏多重比较结果显示,T2组的SGRL显著高于T4、T6和FG组(P<0.05),而SGRW在各组间差异不显著(P>0.05;表 3)。在摄食后,正常摄食的T0组SGRL最高,其次是T4组,其SGRL显著高于T6组(P<0.05);T2组SGRW显著低于其他3组(P<0.05),随着首次投喂时间的延长,T4和T6组的SGRW有所增加,但均低于T0摄食组SGRW(表3)。
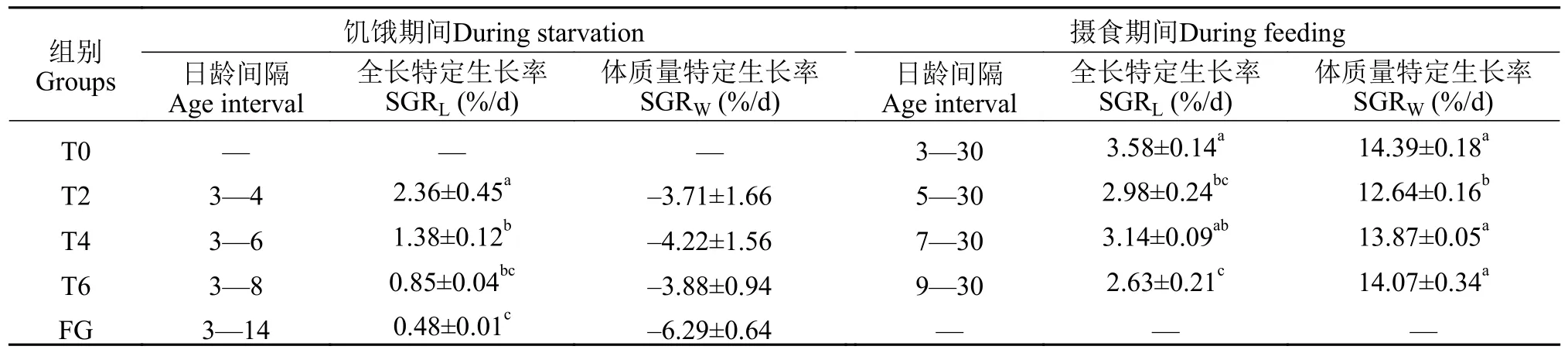
表3 不同处理组的鲢仔鱼在饥饿和摄食期间全长、体质量特定生长率的变化Tab.3 Changes of the specific growth rate of body weight and total length of silver carp larvae for different treaments during starvation and feeding
饥饿组仔鱼在15日龄时已全部死亡,其他各组仔鱼在30日龄时结束实验,因此选择3—14日龄和3—30日龄两个时期研究不同延迟投喂水平对仔鱼生长的影响。延迟首次投喂的时间对仔鱼在14日龄和30日龄的全长和SGRL的影响显著(P<0.05;图 7)。14日龄时,正常摄食的T0组全长和SGRL最高,分别为(12.05±0.86) mm和(4.05±0.14)%/d (图7A和7C);T0、T2和T4组的全长组间差异不显著(P>0.05),但均明显高于T6和FG组(P<0.05;图 7A);T0、T2和T4组的SGRL同样显著高于T6和FG组(P<0.05;图 7C)。30日龄时,T0组全长和SGRL分别为(20.60±3.52) mm和(3.63±0.07)%/d,均显著高于T2、T4和T6组(图7B和7D)。随着延迟投喂时间的延长,14日龄和30日龄仔鱼个体生长差异均呈现先增大后减小的趋势,14日龄时饥饿组全长差异系数最低(CV=2.9),30日龄时T6组全长差异系数最低(CV=16.7;图 7A和7B)。

图7 不同延迟投喂组对鲢仔鱼个体全长和特定生长率的影响Fig.7 Effects of delayed first feeding on total length and specific growth rate of silver carp larvae
3 讨论
3.1 饥饿对鲢仔鱼初次摄食和存活率的影响
初孵仔鱼初次摄食的时间因种类而异,通常受到卵的质量、卵大小和温度等因素的影响[23]。对于大多数仔鱼而言,开口摄食的时间往往发生在卵黄囊完全吸收之前,即会经历一段混合营养期[24,25],而北美牙鲆(Paralichthys californicus)[26]和铅色匀鯻(Leiopotherapon plumbeus)[27]在卵黄囊完全吸收后不久才开始初次摄食。同一种仔鱼在不同温度下,其开口摄食时间和内源性营养消耗速率也有差异。例如,鲢仔鱼在水温(19±1)℃下,孵化后6d初次摄食,在孵化后8—9d卵黄耗尽[20],本研究中鲢仔鱼在水温(24±1)℃下,孵化后3d开口摄食,在孵化后5d卵黄囊耗尽。这说明鲢仔鱼开口摄食时间受到温度制约,可能与水温升高引起仔鱼代谢和食物消化速率加快有关[23],在狭鳕(Gadus chalcogrammus)[6]和兔头瓢鳍虾虎鱼(Sicyopterus lagocephalus)[28]的研究中也发现了相同现象。在本研究中,3日龄时只有36.67%的鲢仔鱼摄取了食物,表明在混合营养初期立即进行外源性摄食的能力有限,这主要是由于仔鱼在刚开口摄食时,其运动和捕食器官尚未发育完善,因此初次摄食率较低[29]。另外,饥饿组仔鱼卵黄囊吸收速率相比摄食组更慢,这可能是由于在饥饿胁迫下,仔鱼延缓了内源营养的吸收速率,是鲢仔鱼对饥饿产生的适应性生态反应[30]。
PNR是评价仔鱼耐受饥饿能力的常用指标,从初次摄食到PNR的时间越长,越有利于仔鱼建立外源摄食,从而提高其存活率。在本实验条件下,鲢仔鱼在11日龄抵达PNR,具有摄食能力的时间约8d,这在淡水鱼类中相对较长[29],反映出鲢在早期补充阶段对自然河流饵料生物缺失的适应性策略。本研究发现,饥饿仔鱼在抵达PNR时的存活率仅为41.33%,于15日龄全部死亡,而摄食组仔鱼在11日龄和15日龄的存活率分别达到72.83%和69%,随着延迟投饵时间的延长,仔鱼的存活率明显下降。这说明初次摄食时间对仔鱼的存活率有较大影响,这与Lima等[31]对银鲶(Rhamdia voulezi)仔鱼延迟首次投喂实验的研究结果一致。从鲢仔鱼初次摄食后随时间变化的存活率曲线来看,摄食组仔鱼和延迟2d投喂组仔鱼的存活率曲线基本一致,而延迟4d和6d投喂组的存活率则一直低于前两者,这可能是因为卵黄囊期仔鱼是鱼类生活史过程中最不能耐受饥饿的阶段[20],因此在卵黄囊完全吸收前建立成功的初次摄食有助于提高仔鱼存活率。
3.2 延迟投喂对鲢仔鱼生长的影响
有研究表明,当仔鱼受到不同程度的饥饿胁迫时,会表现出生长速率下降的趋势[32]。本研究也发现,在饥饿期间,各处理组随着饥饿程度的加深,其全长特定生长率和体质量特定生长率呈总体下降趋势。在恢复摄食后,延迟2—6d投喂组间的全长特定生长率呈先增加后下降的趋势,体质量特定生长率则持续增加,但均低于对应摄食组的特定生长率。这可能是仔鱼在遭受饥饿再恢复投喂后时使用的一种补偿机制,一旦恢复投喂后会出现一个快速生长过程,在其他鱼类中也有类似的补偿生长现象报道[33,34]。但若恢复投喂前经受的饥饿时间过长,则可能导致仔鱼补偿生长能力减弱,造成延迟6d投喂组的全长特定生长率有所下降。延迟6d投喂组在摄食期间的体质量特定生长率并未明显下降,这可能只是摄食量增大的一种假象,一段时间的饥饿会导致仔鱼食欲增强,往往会过度摄入超出自身所需能量的食物[35]。
本研究发现,在14日龄时,摄食组仔鱼的全长和全长特定生长率均高于其他处理组,且延迟0—4d投喂组显著高于延迟6d投喂组和饥饿组,这说明在卵黄囊吸收完成的前后2d内建立初次摄食对仔鱼随后的生长发育至关重要。在30日龄实验结束时,摄食组全长和全长特定生长率显著高于其他处理组,这相较于14日龄差异变大。因此推断,短时间的饥饿胁迫也会对仔鱼的生理状况造成一定损伤,即使在恢复摄食后也无法达到正常摄食组生长水平[36],这种差距可能会随着日龄的增加体现得愈发明显。另外,本研究还发现鲢仔鱼个体生长差异水平也受延迟投喂时间的影响,无论是14日龄还是30日龄,仔鱼个体生长差异均随延迟投喂时间的延长而呈先增大后减小的趋势。但在条石鲷(Oplegnathus fasciatus)仔鱼延迟投喂的实验中,发现个体生长差异随延迟投喂时间(延迟0—2d)的延长而增大的结果[29]。主要原因可能是短时间的饥饿会促使仔鱼摄食能力和行为多样化,导致个体大小差异变大,但当饥饿时间超过4d再摄食后,那些生长发育不佳的个体逐渐被淘汰死亡,只有生长状况良好的仔鱼才可以存活下来,并顺利建立主动摄食能力,从而造成个体大小差异又不断缩小。
3.3 饥饿对鲢仔鱼形态的影响
大量研究表明,仔鱼在饥饿过程中,只能通过消耗自身组织来获取生命活动所需能量,这将导致仔鱼的形态特征受到显著影响[37,38]。例如中华倒刺鲃(Spinibarbus sinensis)仔鱼随饥饿时间的延长,全长、头长和眼径均显著下降[39];饥饿对智利狮鲳(Seriolella violacea)仔鱼的形态性状影响主要体现在体长和肌节高上,饥饿的狮鲳仔鱼体型相比正常摄食的仔鱼更显“细长”[40];太平洋红鲷(Lutjanus peru)仔鱼在受到饥饿胁迫后,肌节高、头长和尾长急剧下降,并对游泳能力造成一定的影响[41]。这与本研究的结果基本一致,本研究中鲢仔鱼遭受5d的饥饿后,饥饿组仔鱼在8日龄体长已显著小于摄食组。此外,由主成分分析得分结果也发现,摄食组和饥饿组的变异主要是由头部形态和肌节高度引起。主成分分析提取的3个主成分累计贡献率达82.27%,表明该方法保留了大部分的变量信息,预测准确度高。
饥饿对仔鱼形态的影响还体现在仔鱼生长发育期间脊索末端无法及时弯曲上。脊索末端向上弯曲是大多数仔鱼发育过程中的一个基本特征,但本研究中饥饿鲢仔鱼直到全部死亡均未见明显弯曲现象,这可能与饥饿仔鱼的体长未达到7.5—8.0 mm有关,即鲢仔鱼脊索开始向上弯曲的大小[42]。有研究也证实,条纹鲮脂鲤(Prochilodus lineatus)仔鱼向下游漂流时脊索未弯曲是一个典型特征[43,44],这是制约硬骨鱼类主动游泳和集群的一个重要因素[45],意味着弯曲前期仔鱼只能依靠水流被动扩散到育幼区。另有研究表明,欧洲鳀(Engraulis encrasicolus)的脊索弯曲与昼夜垂直迁移的开始、耳石生长加快、饵料选择性的变化及逃避被捕食的能力增强有关[46]。因此,可以推断早期饥饿对鲢仔鱼形态的影响还可能会导致鲢仔鱼游泳能力的降低,甚至会增加被捕食的风险。
4 结论
综上所述,在水温(24±1)℃下,鲢仔鱼3日龄开口摄食,11日龄抵达不可逆点,于5日龄卵黄囊耗尽前进行投喂,在实际生产中能有效避免因饥饿而导致鲢仔鱼成活率降低。早期饥饿会影响鲢仔鱼生长速率,随着饥饿程度的加深,鲢仔鱼全长特定生长率和体质量特定生长率呈总体下降趋势。鲢仔鱼在饥饿过程中,头部形态和肌节高度特征受到影响较大,饥饿对鲢仔鱼脊索末端的影响还可能会导致鲢仔鱼游泳能力的降低。因此,在鲢早期发育阶段,应适时投饵或加强野外饵料生物监测,评估饥饿对鲢种群资源补充过程的影响。
