施肥和刈割协同对藏北高原禾草混播群落动态和超产的影响
2023-10-21周娟娟魏巍
周娟娟,魏巍*
(1. 省部共建青稞和牦牛种质资源与遗传改良国家重点实验室,西藏 拉萨 850000;2. 西藏自治区农牧科学院草业科学研究所,西藏 拉萨 850000)
青藏高原高寒草地是响应气候变化和人类扰动敏感脆弱的陆地生态系统,承载着碳固存、畜牧业生产、生物多样性保护和藏族文化传承等多项生态系统服务功能。长期以来,由于忽视高寒草地的生态功能,过度利用其生产功能,造成草-畜关系失衡,草地大面积退化。据报道,青藏高原高寒草地面积为1.27×108km2,占青藏高原总面积的50.9%,退化草地面积约4.5×107km2,其中近1/3 高寒草地退化形势严峻[1]。甚至形成了无法自然修复的大面积次生裸地。现阶段,建植人工草地是重度退化草地和次生裸地恢复的有效方式[2-3],是缓解天然草地放牧压力、提升草地生产力、恢复草地生态功能的重要手段。
众所周知,与单一种群或单播系统相比,多物种群落可有效增加环境资源利用效率,进而长期维持较高的生产力和稳定性[4-6]。Hector 等[7]通过多点联合控制试验指出,生态位分化与互惠理论(互补效应)以及物种/种群属性(选择效应)机制是多物种群落导致草地初级生产力增加的理论基础,而植物多样性维持草地稳定性则是通过超产效应和均衡效应得以实现[8-9]。与经典的人工群落构建研究类似,生态位分化并不明显的禾草混播草地,也会通过选择效应和互补效应,显著提高其地上生物量[10]。
生态学理论认为,环境变化或扰动对群落生产力有重要作用[11-12]。诸多研究发现,施肥明显改变了植物的繁殖分配策略,使个体高大、能对养分做出快速响应的物种获得竞争优势,低矮物种在竞争中处于劣势,易被竞争排除出群落,这是选择效应作用的结果[13-14]。与此相反的观点认为,施肥对一年生草本植被的选择效应并不明显,但却提高了群落的补偿效应[15]。一些学者通过刈割模拟放牧,发现刈割改变了局地光、水和土壤养分利用率,间接影响群落物种结构与分布[16-17],进而促进群落植被有效利用环境资源的正效应[18]。相反,刈割通过抑制群落高度,也会造成植被的垂直空间分布发生分化,这种生态位分化增强了物种间的竞争[19]。尽管这些研究解释了施肥、刈割对物种共存的影响,但还没有形成统一的认识。环境波动、干扰和物种组成紧密联系,甚至协同共变、交互影响物种共存。
受藏北高原寒、旱气候条件的约束,草种适应性是当前构建人工草地修复藏北高原高寒退化草地面临的主要问题。选择与退化草地恢复区域地带性植被接近的乡土物种进行草地恢复,明确不同乡土草种对环境的适应以及群落组成中的共存与竞争关系,厘定物种多样性中的选择效应和互补效应对草地超产的贡献大小,才能确保重建草地群落的稳定性和可持续性。鉴于此,本研究基于在藏北高原选择驯化的乡土草种建植的人工草地群落,对比分析3 种禾草单播与混播及嵌套施肥、刈割处理后,草地群落组分生物量的消长动态,明晰超产效应成因及与多样性的相关性,揭示高寒人工草地生产力维持策略,以期为藏北高原退化草地的生态恢复提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于青藏高原腹地西藏自治区那曲市色尼区那曲镇14 村,地理坐标为北纬31°26′、东经92°07′,平均海拔4450 m,属于高原亚寒带半干旱季风气候,干燥、多风,雨热同季。年均温-2.9 ℃,最冷月(1 月)平均气温为-20 ℃,最热月(7 月)平均气温8 ℃,年均降水量约400 mm,年相对湿度约50%,降水主要集中在6-9 月,无绝对无霜期。试验区地势平坦,为重度退化的高寒草原,主要杂类草有银洽草(Koeleria argentea)、二裂委陵菜(Potentilla bifurca)、紫花针茅(Stipa purpurea)、藏豆(Stracheya tibetifca)、狗娃花(Heteropappus hispidus)、独行菜(Lepidium apetalum)等。土壤类型为高寒草原土,土壤有机质为17.58 g·kg-1,土壤全氮、全磷、全钾分别为1.22 g·kg-1、0.41 g·kg-1和2.42 g·kg-1,土壤速效氮、速效磷和速效钾分别为118.18 mg·kg-1、9.97 mg·kg-1和87.47 mg·kg-1,土壤pH 为8.30,呈微碱性。
1.2 试验材料
本研究供试材料分别为垂穗披碱草(Elymus nutans),麦宾草(Elymus tangutorum)和中亚早熟禾(Poa litwinowiana),均采自藏北高原,在那曲市那曲镇“藏北高寒草地生态科技园”(31°26′N,92°01′E,海拔4512 m)经3 年栽培驯化后,用于多年生混播草地建植。
1.3 试验设计
试验于2019 年6 月初播种,设单播和混播。单播为垂穗披碱草(S1)、麦宾草(S2)和中亚早熟禾(S3)单独播种,播量分别为2.25、2.25 和1.50 g·m-2;混播分为两两混播和3 种混播,有4 个播种组合,为垂穗披碱草+麦宾草(M1)、垂穗披碱草+中亚早熟禾(M2)、麦宾草+中亚早熟禾(M3)和垂穗披碱草+麦宾草+中亚早熟禾(M4),二者混播播量为单播播量的50%,3 种混播为单播播量的33.3%;共计7 个播种组合。播种组合间随机放置,重复4 次,每个组合播种在3 m×4 m 的小区内。播种小区之间设置1 m 的缓冲带。每个播种小区内再设横、竖各0.5 m 的隔离带,将播种小区分割成4 个1.5 m×2.0 m 的子区,分别为对照(control,Con)、刈割(cutting,C)、施肥(fertilization,F)和刈割+施肥(fertilization+cutting,F+C)。施肥处理所施肥料为(NH4)2HPO4,于播种第2 年6月初施肥1 次(施肥后喷灌),施肥量为60 g·m-2(纯氮施用量10.8 g·m-2,纯磷施用量27.6 g·m-2);于牧草灌浆期,在C 和C+F 子区内进行留茬5 cm 的刈割处理。
1.4 种植方法
采用人工开沟条播,混播组合为同行播种。播深3~5 cm,行距25 cm,播后覆土镇压,出苗后人工除杂2 次,入冬前灌冬水,未采取覆土措施,均为自然越冬。
1.5 样品采集与测量
播种翌年在7-9 月(植物生长季)每月20 日进行采样。为了消除相邻子区之间处理效应的相互干扰,每个子区内两边各留1 行不做测量和取样。为确保刈割处理与生长季末的测定处于同一样方内,第一次施肥时在每个子区随机固定2 个0.25 m×0.25 m 的样方。灌浆期在C 和C+F 子区内进行留茬5 cm 刈割,为防止遮阴影响样方内植物的再生长,清除样方周边0.5 m 范围内的植物。地上生物量测定采用刈割收获法,各亚区随机选取2 个0.25 m×0.25 m 样方分种刈割,65 ℃烘干至恒重,分种称重,精确至0.0001 g,计算地上生物量。地下生物量测定采用挖土块法,各子区内用铁锹随机挖取长×宽×高为0.20 m×0.10 m×0.25 m 的根-土混合土块,每个子区随机挖取2 个,装入尼龙网袋,用清水冲洗泥土,除去石块和砂子,带回实验室,按种分开(对已离体的少量毛根,按地上部分的组分分配给各草种),去除地上部分,在65 ℃烘干至恒重,称重,计算地下生物量。
1.6 统计分析
1.6.1 种间竞争力测定 RYT(relative yield total)即相对总生物量,可表征混播植物间竞争力。其计算公式如下:
式中:Yij为混播中i的生物量;Yji为混播中j的生物量;Yii为单播中i的生物量;Yjj为单播中j的生物量。当RYT>1 时,混播组分种间竞争小于种内竞争,表现出共生关系;当RYT=1 时,混播组分种间竞争与种内竞争相等;当RYT<1 时,混播组分种间竞争大于种内竞争,表现出拮抗关系[20]。
1.6.2 超产、超产效应和多样性效应测定 超产(overyield,OY)为混播群落生物量与该群落各组分物种单播生物量均值的差值。OY=Bmc-Bs,式中:Bmc为混播群落地上生物量;Bs为该混播群落各组分物种单播平均地上生物量,OY>0,表明存在超产。
超产效应1(transgressive overyielding effect 1,OY1)为混播群落地上生物量超过该群落中地上生物量最高物种的单播地上生物量,强调了混播物种成分的差异与超产效应之间的联系[21]。
OY1=(Bmc-maxBimno)/maxBimno,式中:maxBimno为混播群落组分中最高产物种的单播地上生物量,OY1>0,表明存在超产效应。
燃料电池的工作参数影响着冷启动的成败,此方法可以不必增加外部设备,直接改变工作参数实现冷启动,这样可以减少成本,使系统结构简化。
超产效应2(transgressive overyielding effect 2,OY2)为混播群落地上生物量超过该群落内各组分物种的平均单播地上生物量,解释了混播群落地上生物量与混播组分单播地上生物量之间的关系[21]。
OY2=(Bmc-Bimno)/Bimno。式中:Bimno为混播群落中各组分物种的平均单播地上生物量,OY2>0,表明存在超产效应。
参照Loreau 等[9]的方法,将混播群落地上生物量的增加量(多样性净效应,ΔY)分解为选择效应(selection effect,SE)和互补效应(complementary effect,CE)。
式中:YO 为混播群落的实际地上生物量,YE 为以单播地上生物量为基础计算的混播群落期望地上生物量,N为互补效应(CE),Ncov(ΔRY,M)为选择效应(SE),M 为混播群落中各组分的单播地上生物量,Δ RY 为混播群落中各组分实际相对地上生物量与其期望相对地上生物量的差值,N为混播群落的物种数。当Δ Y=0 时,表示植物多样性对地上生物量无影响(零假设);当ΔY>0 时,表示群落高产受物种生态位互补作用影响;ΔY<0 时,表示群落中多物种竞争导致群落减产。
1.6.3 单个物种的表现 引用自然对数响应比值(the natural log response ratio,LNRR)来表征单个物种在混播中的表现。
式中:Oi为物种i在混播中的生物量,Mi为物种i的单播生物量,Pi为物种i在混播群落中所占比例。LNRRi>0时,表示混播群落中物种i超产,物种i的种内竞争大于种间竞争;LNRRi<0 时,表示混播群落中物种i低产,物种i的种间竞争大于种内竞争[22]。
1.7 数据处理
采用Excel 2010 进行数据整理计算,采用Origin 18.0 制图,SPSS 20.0 统计软件,进行双因素方差分析(Two-way ANOVA),Duncan 法进行多重比较。
2 结果与分析
2.1 生长季内多年生禾草草地群落组分的消长动态
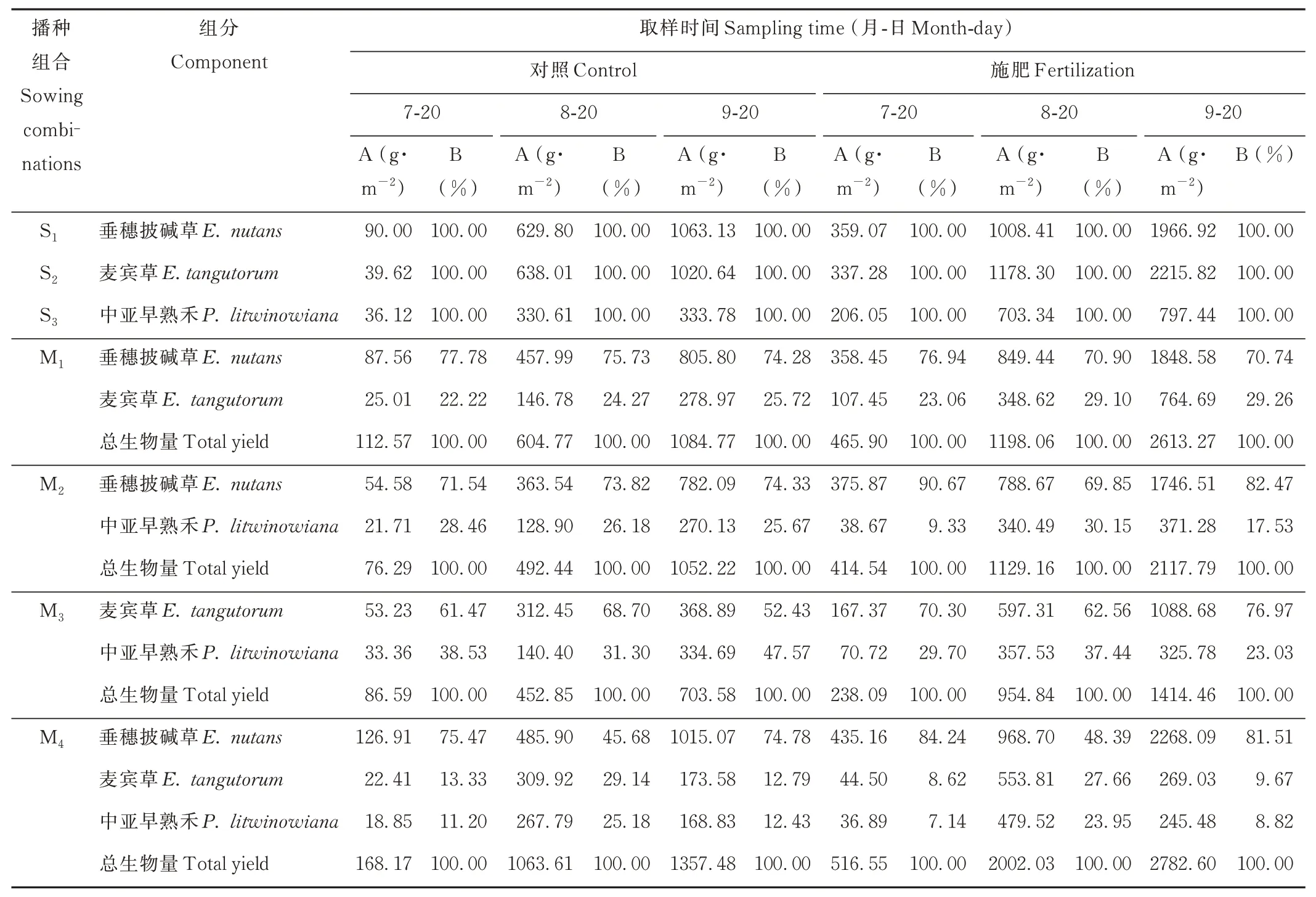
表1 不同播种组合地上生物量各组分的动态变化Table 1 Dynamic change of aboveground biomass under different sowing combinations
2.1.2 生长季内多年生禾草草地地下生物量的群落组分动态变化 单播S1~S3中,3 个取样时期S3的地下生物量均为最高(表2)。施肥增加了地下生物量的积累,单播S1、S2、S3均在9 月20 日最高,分别为264.89、350.01和356.43 g·m-2。混播M1~M4中,地下生物量呈先上升后下降的趋势,地下生物量积累均在8 月20 日达到峰值;其中M4混播组合施肥处理下地下生物量最高,为669.61 g·m-2。M1、M2和M4混播中垂穗披碱草根系生物量占当期地下总生物量比例较高。M3和M4混播组合,中亚早熟禾根系的比例随着物候期推进逐渐增加,9 月20 日,施肥和不施肥条件下分别为53.97%、57.62%和31.06%、32.38%,中亚早熟禾对群落根系的贡献超过麦宾草。
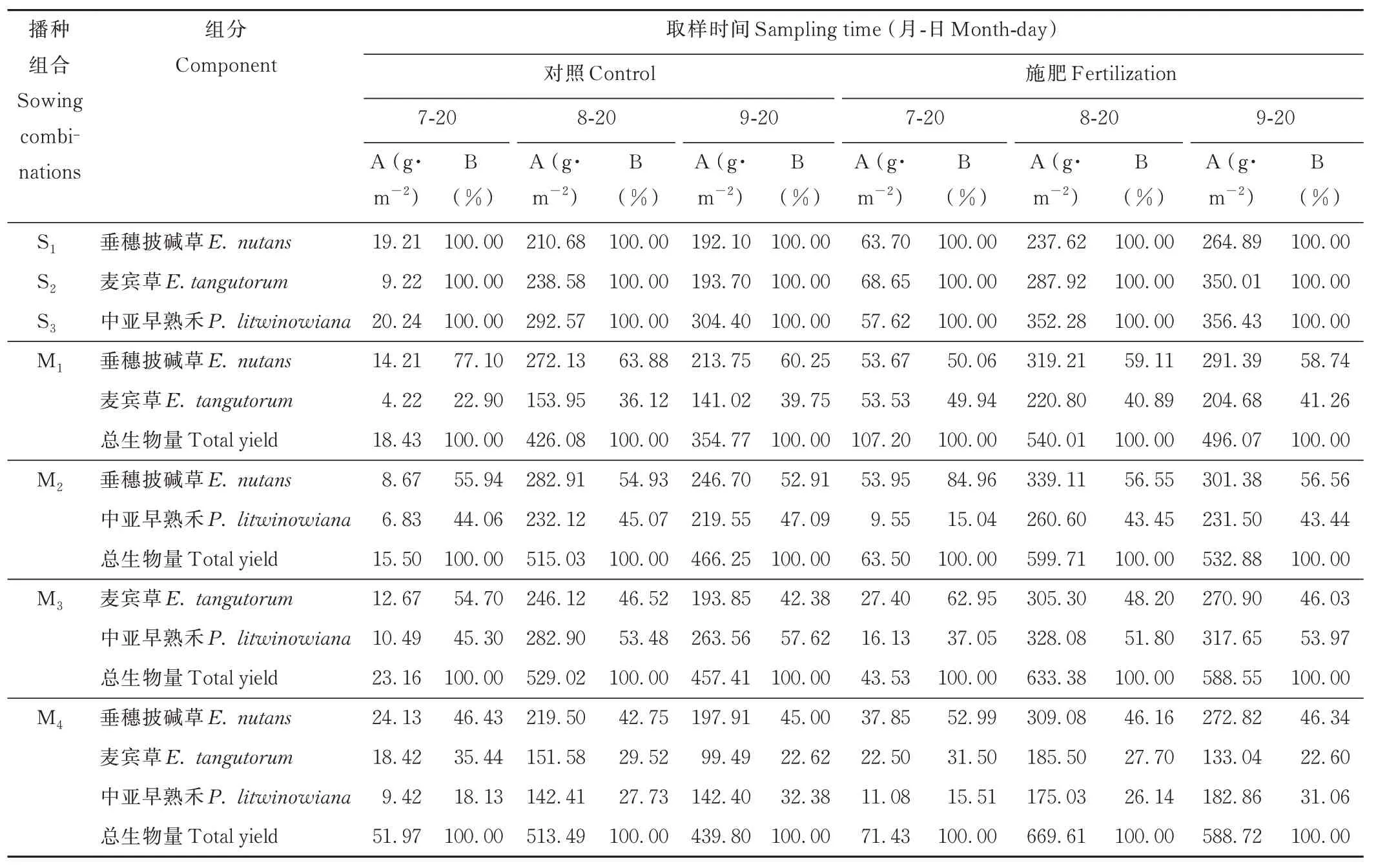
表2 不同播种组合地下生物量各组分的动态变化Table 2 Dynamic change of underground biomass under different sowing combinations
2.2 施肥和刈割对多年生禾草草地地上生物量的影响
方差分析显示(表3),播种组合显著影响群落的地上生物量,施肥和刈割对群落的地上生物量具有显著的交互作用,施肥或刈割分别与播种组合交互影响群落的地上生物量(P<0.05);无论单播还是混播,施肥或刈割对草地地上生物量均有不同程度的影响。
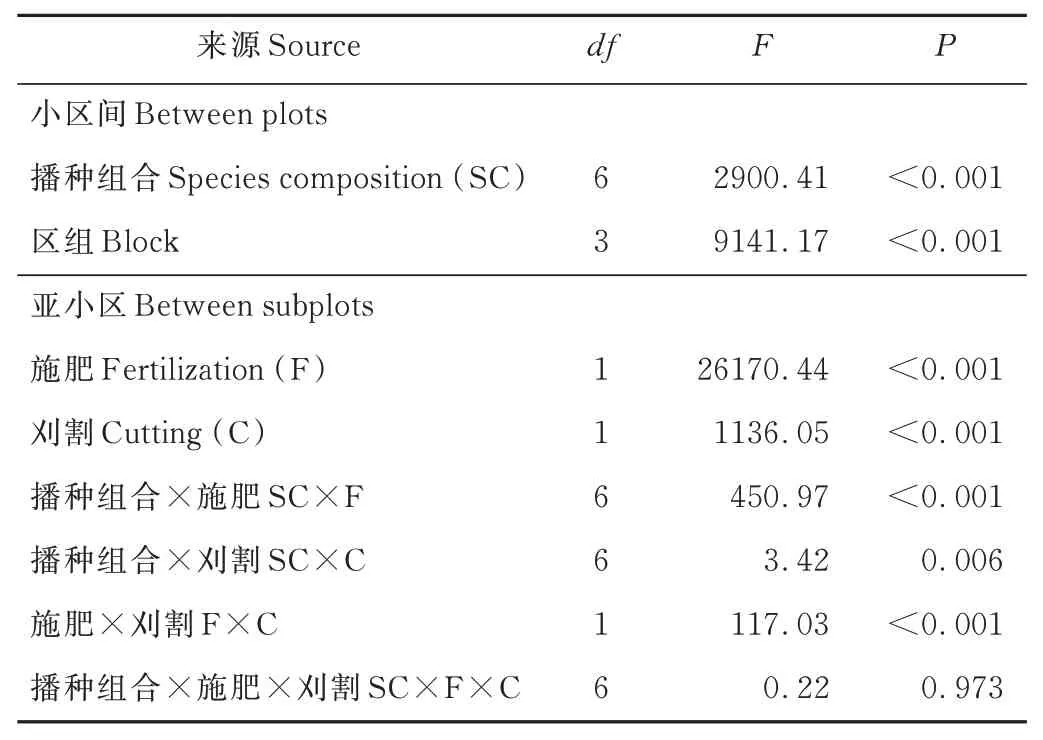
表3 播种组合、施肥和刈割对群落地上生物量影响的方差分析Table 3 ANOVA analysis for the effects of sowing combination (SC),fertilization (F),cutting (C) and their interaction on the aboveground biomass
播种组合间比较,S1和S2的地上生物量高于S3。M1、M2混播组合的地上生物量低于其各组分的单播产量;M4混播的地上生物量大于S1~S3单播和M1、M2、M3混播。与Con 相比,F、C 及C+F 处理均增加了各播种组合的地上生物量,其中F 及C+F 处理均达到显著水平(P<0.05)(表4)。F 处理显著增加了M1、M2和M4播种组合中垂穗披碱草组分的地上生物量,与Con 相比涨幅分别达129.41%、123.31% 和123.44%。
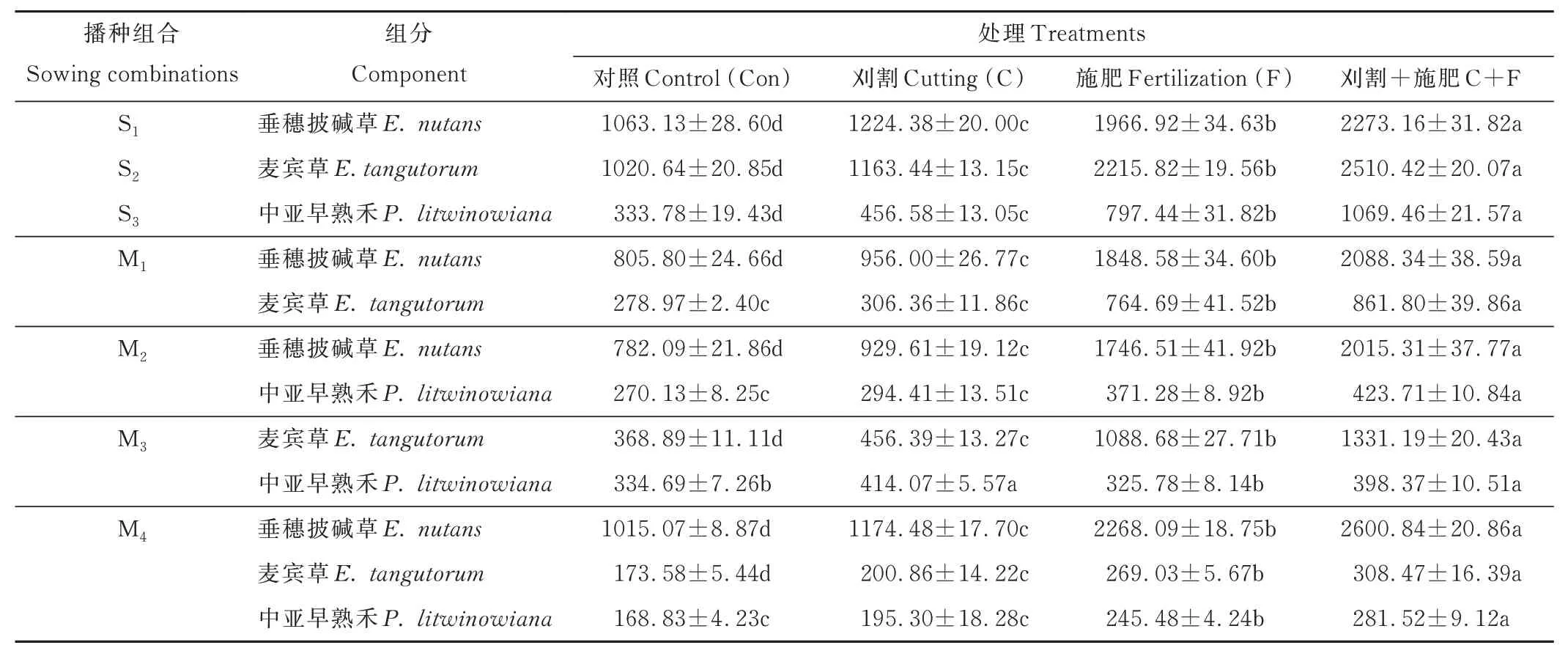
表4 不同播种组合在不同处理下的地上生物量Table 4 Aboveground biomass of different treatments under different sowing combinations (g·m-2)
2.3 施肥和刈割对多年生禾草草地相对产量总和(RYT)的影响
由图1 可知,Con、C、F 和C+F 处理下混播M1、M2和M4的RYT 值均大于1.0;其中,混播M4的RYT 值最高,分别为1.63、1.53、1.58 和1.53,表明垂穗披碱草+麦宾草混播、垂穗披碱草+中亚早熟禾混播和垂穗披碱草+麦宾草+中亚早熟禾混播各组分的种内竞争均大于种间竞争。Con 和C 处理下混播M3的RYT 值大于1.0,而在F 和F+C 处理下小于1.0,表明麦宾草+中亚早熟禾混播组分的种内竞争大于种间竞争,施肥后种间竞争增强,趋近于种内竞争。
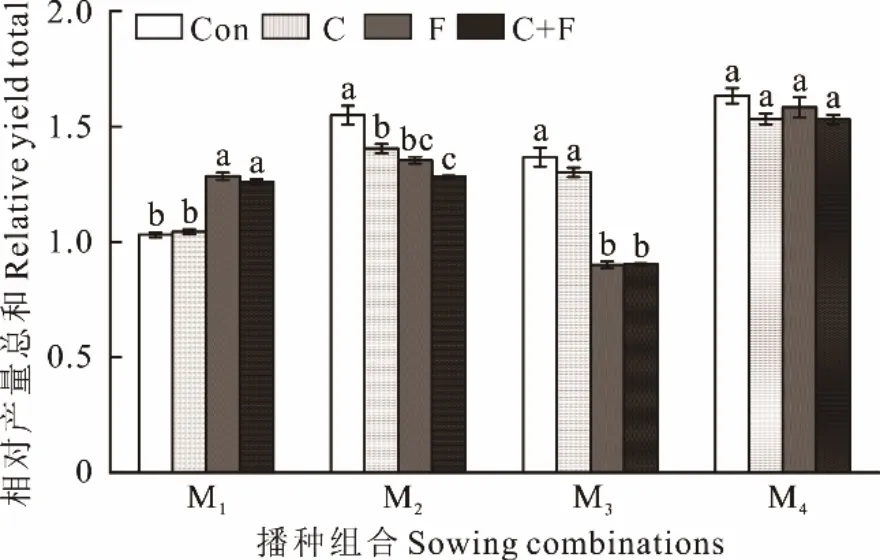
图1 不同播种组合多年生禾草草地的RYTFig. 1 Relative yield total of perennial grass pasture under different sowing combinations
2.4 施肥和刈割对多年生禾草草地超产及超产效应的影响
除了M3混播组合的F 和C+F 处理,其他混播组合的不同处理与单播相比均存在超产,M4混播组合在Con、C、F 和F+C 处理下分别超产551.70、622.51、1122.57 和1239.87 g·m-2(图2)。F 和C+F 处理可显著提高M1、M2和M4混播组合的超产效应(P<0.05)。Con、C、F 和C+F 处理下M1和M4混播组合OY1大于0,F 和F+C 处理下M2混播组合的OY1大于0,表明存在超产效应1;Con 和C 处理下M2和M3混播组合的OY1均为负值,表明不存在超产效应1。M1混播组合的OY1值在F 和F+C 处理时显著高于Con 和C 处理(P<0.05)。Con、C、F 和C+F 处理下M1、M2和M4混播组合的OY2大于0,Con 和C 处理下M3混播组合的OY2均大于0,表明存在超产效应2。M1混播组合的OY2在F 和F+C 处理时显著高于Con 和C 处理(P<0.05);M2混播组合的OY2在F 处理时显著高于C 和F+C 处理(P<0.05);M4的OY2在F 处理时显著高于F+C 处理(P<0.05)。
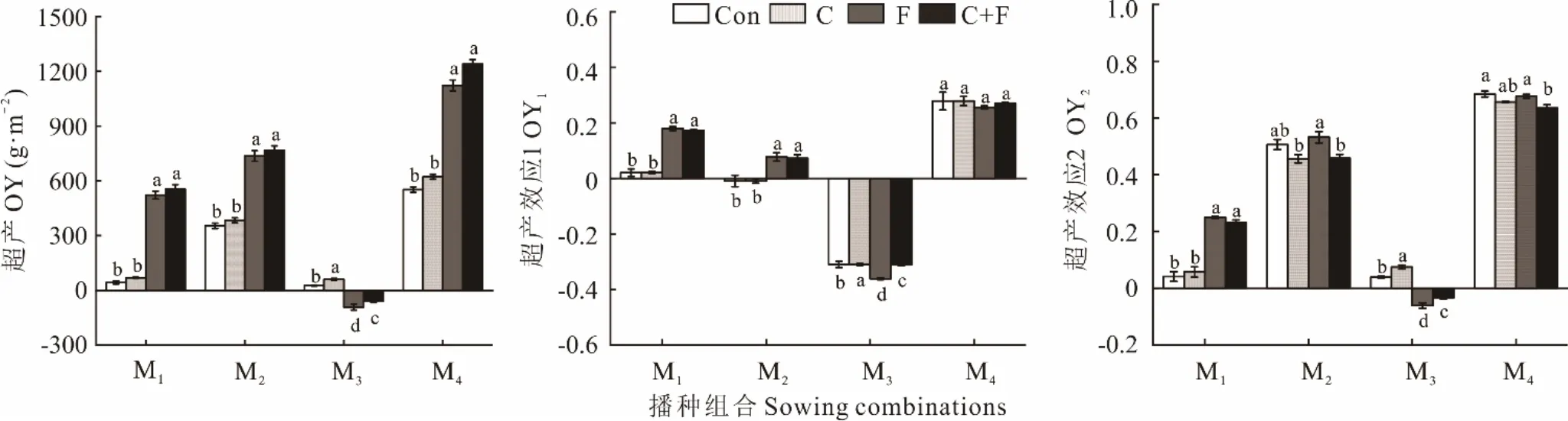
图2 不同播种组合多年生禾草草地超产和超产效应Fig. 2 Change in over yielding and transgressive overyielding effect of perennial grass pasture under different sowing combinations
2.5 施肥和刈割对多年生禾草草地多样性净效应的影响
除混播M3的F 和F+C 处理,其他不同混播组合的多样性净效应均大于0,说明物种生态位的互补使得群落地上生物量高产(图3)。M1、M2和M4混播组合的多样性净效应在F 和F+C 处理时显著高于Con 和C 处理(P<0.05)。混播M3的多样性净效应在F 和F+C 处理时为负值,说明群落物种因竞争导致减产。M2混播组合在F 和F+C 处理时选择效应较高,M3混播组合在Con 和C 处理时较低且为负值;M1混播组合在Con 和C 处理时选择效应大于0;M4混播组合中各处理的选择效应均大于0。M3混播组合中F 和F+C 处理的互补效应小于0,其他播种组合在各处理下互补效应均大于0,且相同处理下M4混播组合的值最大。M1、M2和M4混播组合的互补效应在F和F+C 处理时显著高于Con 和C 处理(P<0.05)。

图3 不同播种组合多年生禾草草地的多样性净效应、选择效应和互补效应Fig. 3 Change in net effect of biodiversity,selection effect and complementarity effect of perennial grass pasture under different sowing combinations
由图4 可知,将OY1与多样性净效应、选择效应和互补效应进行拟合,发现均呈显著正相关关系(P<0.01)。

图4 超产效应1 与多样性净效应、选择效应和互补效应的关系Fig. 4 The relationship between transgressive overyielding effect 1 and net effect of biodiversity,selection effect and complementarity effect
2.6 施肥和刈割对多年生禾草草地单个物种表现的影响
区组水平上,垂穗披碱草和中亚早熟禾的LNRR 值都大于0,说明这两个物种在混播群落中都表现出超产,其种内竞争大于种间竞争(图5a)。麦宾草在所有混播中LNRR 值均小于0,混播群落减产,说明其种间竞争大于种内竞争。亚区组水平,与对照相比,刈割一定程度上削弱了中亚早熟禾的生长表现(图5c);施肥促进了垂穗披碱草的超产表现,抑制了中亚早熟禾的生长,造成其减产(图5d);施肥和刈割交互影响麦宾草+中亚早熟禾混播组合两个物种的表现,刈割存在时,施肥增强了对中亚早熟禾的抑制作用(图5e)。
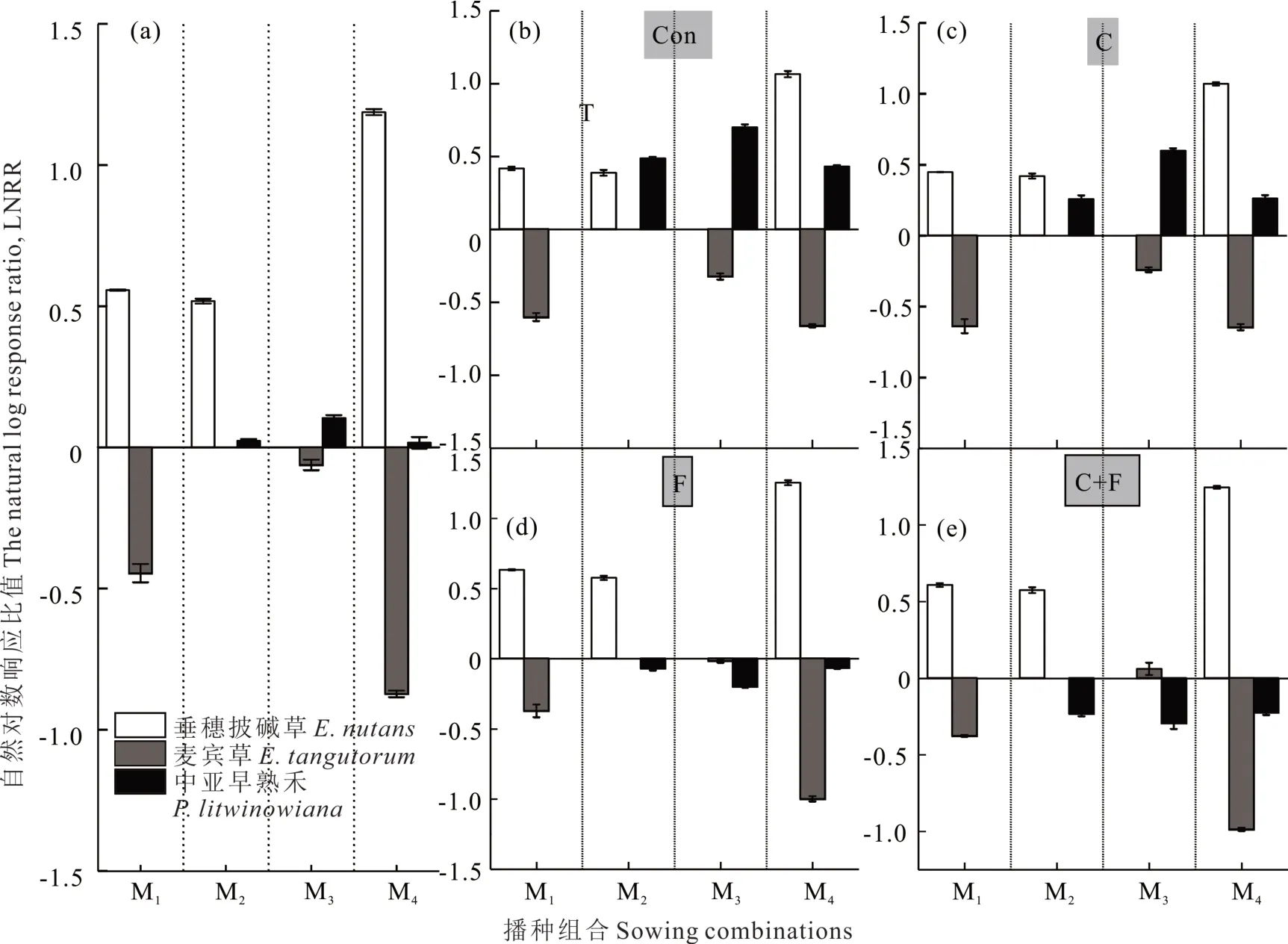
图5 不同播种组合多年生禾草草地的LNRR 值Fig.5 The value of LNRR of perennial grass pasture under different sowing combinations
3 讨论
3.1 混播对禾草草地物种共存的影响
生态位理论认为,生态位分离是物种实现共存的一条途径,混播系统降低种群在环境资源利用方面的生态位重叠,使群落占据更广的功能空间,提高资源利用率。本试验中,M1、M2和M4混播组合中(除8 月20 日),垂穗披碱草地上生物量均能达到当期群落总生物量的70%以上(表1),这主要是因为垂穗披碱草个体较大、竞争能力强,为优势物种,能获得更多光资源,能够迅速占领较高生态位,在混播系统中居于主导地位,而麦宾草、中亚早熟禾竞争能力次之,长势较弱。8 月20 日,M4混播组合中垂穗披碱草对群落地上生物量的贡献降低至50%以下,相反,麦宾草和中亚早熟禾对群落产量和地下生物量的贡献率均达到峰值。是因为垂穗披碱草前期生长较快,群落中地上、地下生物量均占优势;而中亚早熟禾返青后生长缓慢,到8 月20 日其生长进入旺盛期,地下生物量积累迅速,造成垂穗披碱草地下生物量占比下降。说明物种间根系生长的物候差异有利于混播系统的互补性[23]。与单播相比,垂穗披碱草、麦宾草和中亚早熟禾3 物种混播群落地上生物量最高,为1357.48 g·m-2,植物多样性的增加提升群落整体资源利用率,进而获得高产。混播系统的生态位分化造成空间、光照和养分资源在物种间的分配差异。本研究中,除了混播M3的F 和F+C 处理,各混播组合的RYT 均大于1.0(图1),表明这些混播组合的种间竞争小于各组分物种的种内竞争,不同物种利用生态位分离,达到稳定共存的目的。揭示物种多样性与生产力之间的作用机理是多样性-生产力关系研究中的核心问题,超产效应可作为解释植物多样性与群落或生态系统生产力正相关的重要证据[24-25]。本试验中,除M3混播组合中的F 和F+C 处理,其他混播组合的不同处理均存在超产效应,且M4的OY1和OY2均高于其他混播组合(图2)。混播群落中,当选择效应占主导地位时,群落中竞争能力较强的物种对系统的影响较大,而当互补效应占主导地位时,群落中不同物种相互促进,提高资源利用率,从而对系统产生较大的影响[26]。本试验发现,在多年生禾草混播草地建植初期,M1、M4混播组合中的Con、C 处理和M2混播组合中的C、F、F+C 处理是由选择效应、互补效应共同主导超产效应,M1混播组合中F 和F+C 处理、M2混播组合中的Con 处理、M3混播组合中的Con 和C 处理是由互补作用主导超产效应(图3)。超产效应与选择效应、互补效应和多样性净效应均呈显著正相关性(图4),且互补效应值大于选择效应(图3),说明互补效应是影响超产的主要因素。Cardinale 等[27]对44 个生物多样性试验整合分析发现,互补效应可解释大部分生物多样性效应。黎松松等[28]比较一年生和多年生禾豆混播草地超产效应表明,互补效应对超产效应的作用更为关键。互补效应通过物种间的生态位分化,使群落中物种功能特性的多样性增加,最终促进系统功能水平的提高[29]。这也印证了青藏高原草地植物群落物种间具有较强的正相互作用[30]。
3.2 施肥和刈割对禾草草地物种共存的影响
施肥和刈割等管理措施,是调节草地生态系统生产的关键因子[31]。本研究中,与未施肥相比,施肥后不同生育阶段各播种组合组分物种的生物量(地上和地下)均有不同程度增加(表1 和表2),植株高大的垂穗披碱草、中等高度的麦宾草和低矮的中亚早熟禾对施肥均为正向响应,地上生物量均获得提升,这种正响应降低了物种的生态位重叠,减少了物种间的环境竞争,促进了物种共存。尽管垂穗披碱草和麦宾草为同属植物,对资源需求相似,但优势物种垂穗披碱草种内竞争大于种间竞争,即同种个体在资源需求上比异种个体更为相似[32],故垂穗披碱草+麦宾草仍能共存,获得超产。吴晓娟等[33]认为,禾草地下生物量与施肥存在正效应,通过根系分泌物与脱落物返还有机物提高土壤养分,显著增加群落生产力。施肥条件下中亚早熟禾地上生物量的涨幅小于两种披碱草属植物,其地下生物量增幅却优于两种披碱草属,表明与地上部分相比,垂穗披碱草根系的竞争优势弱化,中亚早熟禾的竞争力增强,生长潜势较强。不同物种对环境扰动的响应具有异步性,环境波动后物种的反应存在差异[34]。本研究中,不同混播组合中刈割处理垂穗披碱草组分地上生物量显著增加,麦宾草和中亚早熟禾组分地上生物量增加较小(表4)。刈割对混播群落的超产作用不显著,而施肥对混播群落的超产作用显著,且施肥与刈割有显著的交互作用(图2),即刈割减弱了施肥引起的光竞争,促进物种共存,M1、M2和M4混播组合均出现超产效应。单个物种的LNRR 值表明,刈割对中亚早熟禾生长有一定限制;且刈割增强了施肥对中亚早熟禾的抑制作用(图5d),可能的原因是刈割削弱了施肥对植被地上空间分异的正面效应[35]。尽管刈割条件下各混播群落在施肥后地上生物量显著增加,但并不表示建植的禾草草地在解除养分限制后是刈割引起的超产,因为混播群落超产对刈割响应不显著。因此,本研究中混播草地的表现效应主要是由施肥所致。对于建植的禾草人工草地,适量追加肥料,既可弥补随着时间推移土壤流失的养分,促进人工草地生态系统呼吸[35],又能提高草地生产力。选择高产低碳的乡土草种配套种植管理方式是青藏高原可持续发展的关键。另外,草地物种间的竞争具有不对称性,由此衍生的竞争力会因草地龄级发生变化,因此物种竞争需要开展长期的研究。
4 结论
生长季(拔节期-孕穗期-成熟期)内,地上生物量均在生长季末达到峰值。垂穗披碱草+麦宾草+中亚早熟禾混播地上生物量最高,施肥和施肥+刈割显著增加群落生产力。乡土草种混播群落组分种间竞争小于种内竞争,且都存在超产;其中,垂穗披碱草+麦宾草、垂穗披碱草+麦宾草+中亚早熟禾混播组合中对照和刈割处理和垂穗披碱草+中亚早熟禾混播组合中刈割、施肥、施肥+刈割处理均是选择效应和互补效应共同主导超产效应,垂穗披碱草+麦宾草混播组合中施肥和施肥+刈割处理、垂穗披碱草+中亚早熟禾混播组合中对照处理、麦宾草+中亚早熟禾混播组合中对照和刈割处理是互补作用主导超产效应。
