土壤有机磷矿化特征对高寒草甸退化及温度的响应
2023-10-21钱虹宇蒲玉琳郎山鑫李怡燃周南丁
钱虹宇,蒲玉琳,郎山鑫,李怡燃,周南丁
(四川农业大学资源学院,四川 成都 611130)
磷的有效性不仅调控着草地系统初级生产力的高低,还对维持草地诸多生态功能如生物多样性、涵养水源和固存碳氮等具有重要作用[1]。在自然条件下,草地生态系统中的磷主要来源于土壤母质,进入土壤中与土壤组分相互作用形成无机磷和有机磷两种形态。对于有机磷,因草地土壤有机质含量普遍较高,其含量占土壤全磷的比例高达80%[2],使得植物所需的可溶性无机磷酸盐大部分来自有机磷矿化[3]。因而探明草地土壤有机磷的矿化特征可保障草地系统生产力与生态功能的可持续发展。
草地土壤有机磷矿化受气候、地形、植被、人为活动(如放牧、施肥)和土壤自身特性的影响[3],其中气候、地形与人为活动往往通过影响植被与土壤特性间接影响磷矿化[4]。所以,自然成土环境变化与放牧等人为活动都或多或少干扰着草地土壤有机磷矿化,进而影响草地土壤磷的有效供应。在过去人口剧增、城市化和工业化快速发展的几十年,气候暖干化、过度放牧与鼠害等因素改变了草地系统的水热状况与植被建群种等,草地土壤磷有效性因此呈现降低趋势,甚至可能取代氮或者与氮并存成为草地生产力的限制养分[5]。为探明土壤供磷能力对气候与植被等环境及其变化的响应,一些研究探讨了温带稀树草原和低海拔地区草地土壤有机磷因植被[6]、土壤性质(如土壤质地与水分含量等)[7-8]、地形[9]的差异,以及放牧影响下的矿化特征[10]。高寒地区气候寒冷,土壤有机磷矿化作用极其微弱,磷素有效性较低,往往是影响草地生产力的关键因子之一[11]。因此,为维系高寒草地的生态平衡与功能或实现受损高寒草地的生态振兴,不同气候区域高寒草地土壤有机磷的组分及其分配比例、矿化特征等磷循环理论及其实践应用逐渐成为土壤生态等学科关注的热点。
若尔盖高原是我国乃至世界范围内高寒草甸的典型分布区域,平均海拔约3300~3700 m,气温低,生态极为脆弱。近50 年来,气候暖干化和草地资源过度利用及不合理管理方式致使该地区高寒草甸出现不同程度退化,退化面积占比约21.6%[12],土壤磷的输入和有效性因此降低,严重影响了草甸草地的生产力、生态功能和牧业经济发展[13],说明退化草甸草地的生态恢复亟需厘清土壤供磷现状与潜力。可是,纵观国内外文献针对高寒草甸土壤磷循环过程的研究主要集中在土壤磷的含量状况[13-15]、分布特征[16]及生态化学计量比[17],关于不同退化程度高寒草甸土壤有机磷矿化特征与影响机制亟待明晰。因此,探究高寒草甸退化条件下土壤有机磷矿化的变化特征及温度效应,有助于为全球变暖背景下退化高寒草甸土壤磷养分的科学调控提供基础数据与理论支撑,促进区域牧业经济的高质量发展。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东缘的若尔盖高原花湖湿地自然保护区及毗邻地区,地理位置E 102°49′-102°50′、N 33°55′-33°56′,行政上隶属四川省阿坝藏族羌族自治州的若尔盖县(图1)。域内地质地貌是由喜马拉雅山构造运动和新构造运动的剧烈抬升形成的河谷平原,海拔约3500 m,年平均气温0.6~1.5 ℃,生长季平均气温15 ℃,年平均降水量600~750 mm,该区域气候属高原亚寒带大陆性季风气候;植被建群种类为嵩草属(Kobresia)、薹草属(Carex)、扁穗草属(Brylkinia)等莎草科类湿、中生植物,土壤是由冲积物夹少量坡积、洪积物母质发育形成的草甸土,流经的河流属于黄河水系的黑河及其支流。20 世纪80 年代以来,受牧业经济高速发展下的过度放牧,加之暖干化气候、鼠害的综合影响,域内除了花湖湿地保护区内有保护完好的高寒草甸,其他区域草甸都呈现出不同程度的退化,甚至沙化,这正好是探讨高寒草甸退化对土壤有机磷影响的理想场所[18]。
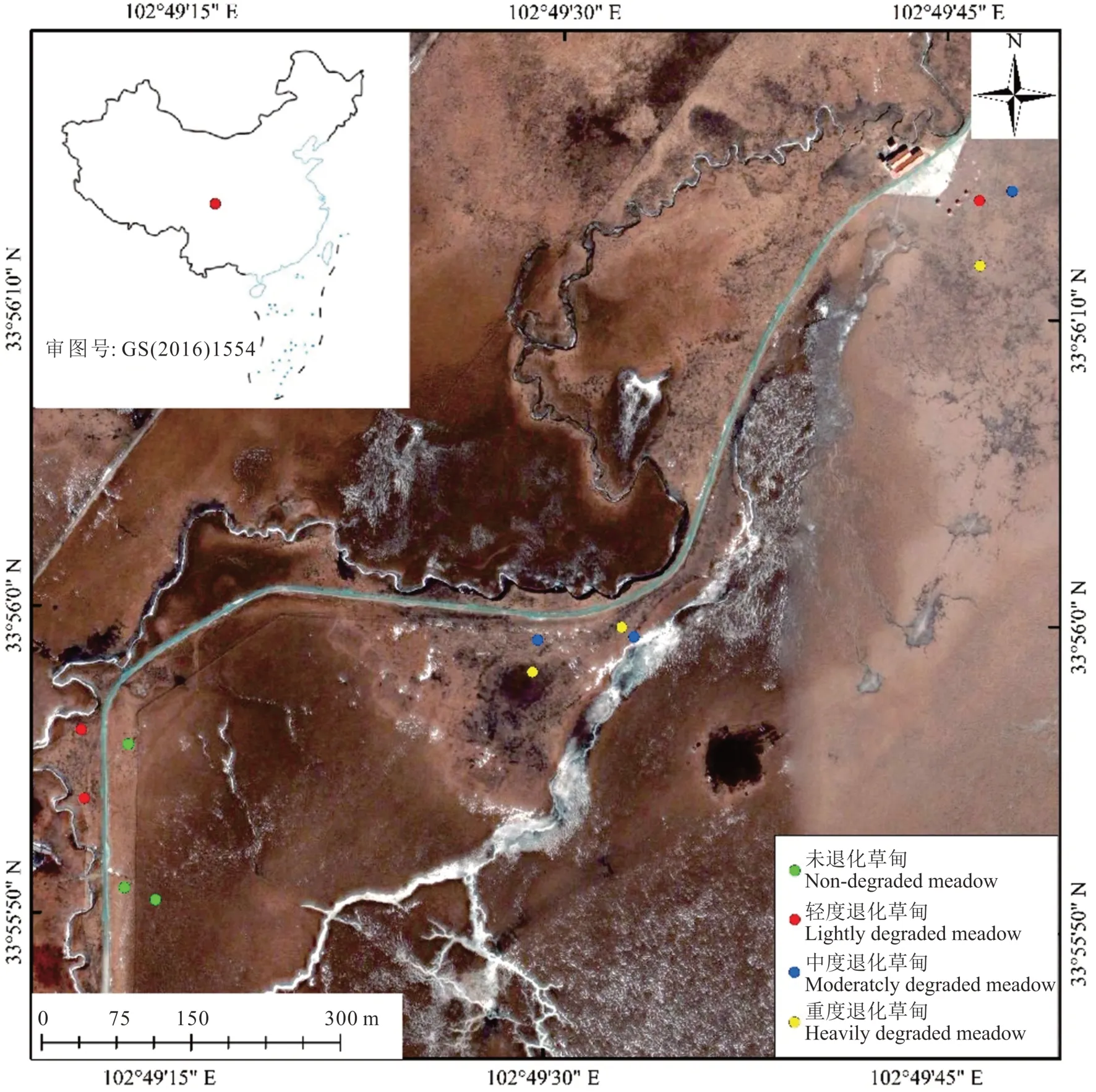
图1 研究区域和样地分布Fig.1 Study area and sampling plots
1.2 样地选择与样品采集
2019 年8 月,根据天然草地退化、沙化与盐渍化的分级指标(GB 19377-2003)[19]的规定和课题组前期的野外调查结果,主要依据植被盖度、高度等植被特征和鼠害程度将高寒草甸分为未退化草甸(non-degraded meadow,ND)、轻度退化草甸(lightly degraded meadow,LD)、中度退化草甸(moderately degraded meadow,MD)和重度退化草甸(heavily degraded meadow,HD)(表1)。

表1 不同退化程度草甸的主要生境特征Table 1 Main habitat characteristics of meadows with different degradation degrees
按照不同退化程度草甸的主要生境特征(表1),选择未退化草甸和3 类退化草甸样地各3 个,大小50 m×50 m。用手持GPS(合众思壮集思宝G120BD,北京)记录地理坐标后,在每个样地随机布设3 个1 m×1 m 的小样方,调查植被盖度,用全收获法采集40 cm×40 cm 内的地上植物,将其装入塑料自封袋用于测定地上生物量;挖取长×宽×高为20 cm×20 cm×20 cm 完整土块1 个,用于测定地下根系生物量;以100 cm3环刀采集原状土,用于测定土壤容重及田间持水量。在每个采样点取0~20 cm 土层土壤样品,将同一样地的3 个小样方土样放置在一起混匀,缩分为4 kg,装入塑封袋。土壤带回实验室后,一部分于室内风干,用于测定土壤理化性质,另一部分过2 mm 筛冷藏于4 ℃冰箱内用于有机磷矿化的培养试验。
1.3 土壤有机磷矿化实验
土壤有机磷矿化采用室内恒温控湿好气培养法。根据植物生长季(4-9 月)的多年月平均气温将土壤培养温度设为5、10、15 ℃,湿度设为有利于微生物活动的最佳含水量即80%田间持水量。称取已剔除根系等非土物质,过2 mm 筛的土壤样品40 g(干重),装入干净干燥的塑料瓶,调节土样水分含量至80%田间持水量,置于预先设置温度为5、10、15 ℃的恒温恒湿培养箱中预培养1 周后分别在第0、1、3、7、14、30、60、90、120 d 时测定土壤无机磷含量。4 种不同退化程度的供试高寒草甸土壤,都在每个取样时间内设置3 个重复,培养期间每隔2~3 d 补水一次。
1.4 指标测定方法
地上生物量测定采用全收获法[20];地下生物量是将带根系的土块晾干过2 mm 筛后,再放置到0.5 mm 筛经水冲洗,得到干净的根系于65 ℃烘至恒重[21]。土壤容重和田间持水量采用环刀法[21],无机磷含量采用1 mol 1/2 H2SO4浸提(液土比50∶1)-钼锑抗比色法[22]。参考鲁如坤[22]《土壤农业化学分析方法》测定土壤有机质、全氮、全磷、碱解氮、有效磷和有效钾含量。
1.5 数据计算与统计分析
1.5.1 土壤有机磷累积矿化量(cumulative mineralization of organic phosphorus,CPM) 土壤的有机磷累积矿化量(mg·kg-1)指一个培养时段内土壤无机磷(Pi)含量的净变化量,即土壤培养第j天时的Pi含量与第0 天的Pi含量的差值,按公式(1)计算:
1.5.2 土壤有机磷净矿化速率(net mineralization rate of organic phosphorus,RNPM) 土壤的有机磷净矿化速率(mg·kg-1·d-1)指一个培养时段内的无机磷变化量(ΔPi)与培养时间(t)的比值,按公式(2)计算:
1.5.3 土壤有机磷矿化的动力学特征 土壤有机磷矿化的动力学特征由抛物线模型参数表征,按公式(3)拟合:
式中:A0是常数,反映了土壤有机磷矿化初始量;B0也是常数,表征土壤有机磷矿化的快慢,值越大矿化速率越快。
1.5.4 土壤有机磷矿化的温度效应 土壤有机磷矿化对温度的响应效应采用二阶函数模拟,按公式(4)、(5)拟合:
式中:R为有机磷累积矿化速率(mg·kg-1·d-1),T为培养温度(℃),A、B和C是常数,其中C为基质质量指数,指温度为0 ℃时的土壤有机磷矿化速率常数(mg·kg-1·d-1);y″值为二次函数的凹率(即二阶偏导数,本研究称为温度敏感系数),值越大说明矿化速率增长越快,对环境升温的敏感性越高。其中R是某一时间段内,土壤有机磷累积矿化量(CPM)与培养时间(t)的比值,按公式(6)计算:
1.6 数据处理
运用Microsoft Excel 2019 和SPSS 20.0 进行数据计算和统计分析,采用单因素方差分析(One-way ANOVA)和邓肯(Duncan)法分析不同退化程度草甸土壤RNPM、A0、B0、R、A、B、C、y″等指标的差异显著性,统计检验的显著水平为P=0.05。采用Origin 2020 和R 语言绘图。
2 结果与分析
2.1 植被和土壤理化性质
植被盖度、地上生物量和地下生物量均表现为随高寒草甸退化程度的加剧而逐渐降低(表2)。与ND 相比,LD 的植被盖度、地上生物量、地下生物量分别显著降低12.2%、22.6%、45.4%,MD 的分别显著降低20.9%、66.1%、52.6%,HD 的分别显著降低42.5%、78.5%、95.1%。3 种退化草甸之间,HD 的植被盖度、地上生物量、地下生物量都显著低于LD 和MD,降低幅度分别为27.3%~34.5%、36.6%~72.3%、89.8%~91.1%;但是LD与MD 间的植被盖度、地上生物量和地下生物量无显著差异(P>0.05)。表明高寒草甸退化显著降低了植被盖度和生物量,且未退化到轻度退化、中度退化甚至重度退化阶段是草甸植被盖度和生物量下降的主要阶段。
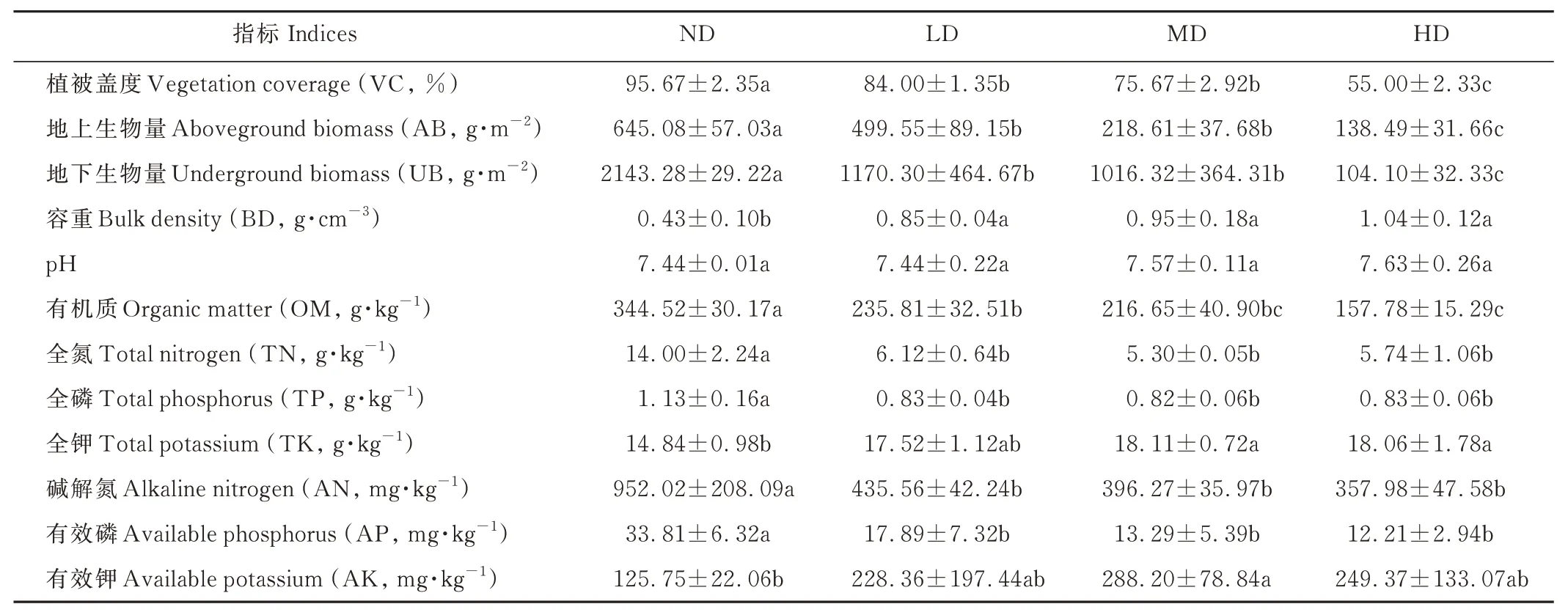
表2 不同退化程度高寒草甸的主要植被特征和土壤理化性质Table 2 Main vegetation characteristics and soil physical and chemical properties of alpine meadows with differently degradation degrees
高寒草甸退化降低了土壤有机质、全氮、全磷、碱解氮、有效磷含量,而增加了土壤容重、pH 值、全钾含量(表2)。其中,退化草甸土壤的有机质、全氮、全磷、碱解氮、有效磷含量分别比ND 显著降低31.6%~54.2%、56.3%~62.1%、26.5%~27.4%、54.2%~62.4%、47.1%~63.9%(P<0.05),HD 有机质含量比LD 显著降低33.1%(P<0.05);退化草甸土壤容重比ND 显著高97.7%~141.9%(P<0.05),MD 和HD 土壤全钾含量分别比ND 高22.0%、21.7%(P<0.05),MD 土壤有效钾含量较ND 显著高129.2%(P<0.05)。不同退化程度土壤pH 之间无显著差异(P>0.05)。
2.2 土壤有机磷矿化动态
如图2 所示,随培养时间增加,4 类草甸土壤CPM 在前期(0~30 d)呈波浪形变化,中期(30~60 d)增加较快,后期(60~120 d)增速逐渐减小。每类草甸土壤CPM 随温度升高而显著增加(P<0.05)。土壤CPM 因草甸退化而减小,在15 ℃的90、120 d 时差异显著(P<0.05)。第120 天时,4 类草甸土壤CPM 的大小为ND(78.8 mg·kg-1)>LD(69.6 mg·kg-1)>MD(40.8 mg·kg-1)>HD(39.8 mg·kg-1),其中ND、LD 显著高于MD 和HD(P<0.05)。

图2 土壤有机磷累积矿化量动态Fig. 2 Dynamic of cumulative mineralization of soil organic phosphorus
如图3 所示,随着培养时间的推移,4 类草甸土壤RNPM在5~15 ℃下基本都是先增加后降低,后期(60~120 d)逐渐趋于稳定。表3 显示,120 d 培养期内,4 类草甸土壤RNPM都对温度变化的响应显著,但各培养阶段又有所差异。ND 土壤RNPM变化范围是-31.0~8.5 mg·kg-1·d-1,在5、10、15 ℃时平均为-4.6、-4.0、-0.8 mg·kg-1·d-1,除14~60 d 外,其余时段内存在温度间的显著差异(P<0.05)。LD 的在-20.4~3.5 mg·kg-1·d-1之间变化,在5、10、15 ℃时平均为-4.0、-3.3、-0.3 mg·kg-1·d-1,且除0~3 d、30~60 d、90~120 d 外的其余时段内RNPM在各温度间差异显著(P<0.05)。MD 的RNPM在-15.1~3.8 mg·kg-1·d-1之间变化,在5、10、15 ℃时平均为-3.2、-2.1、-1.4 mg·kg-1·d-1,各温度间RNPM的差异仅在前期(3~7 d)显著(P<0.05)。HD 的RNPM在-19.2~4.3 mg·kg-1·d-1之间变化,在5、10、15 ℃时平均为-3.7、-2.8、-1.8 mg·kg-1·d-1,在前期(0~7 d)、中期(14~60 d)存在温度间的显著差异(P<0.05)。这初步说明高寒草甸发生退化后土壤有机磷矿化速率波动范围减小,矿化速率对温度的敏感性降低。
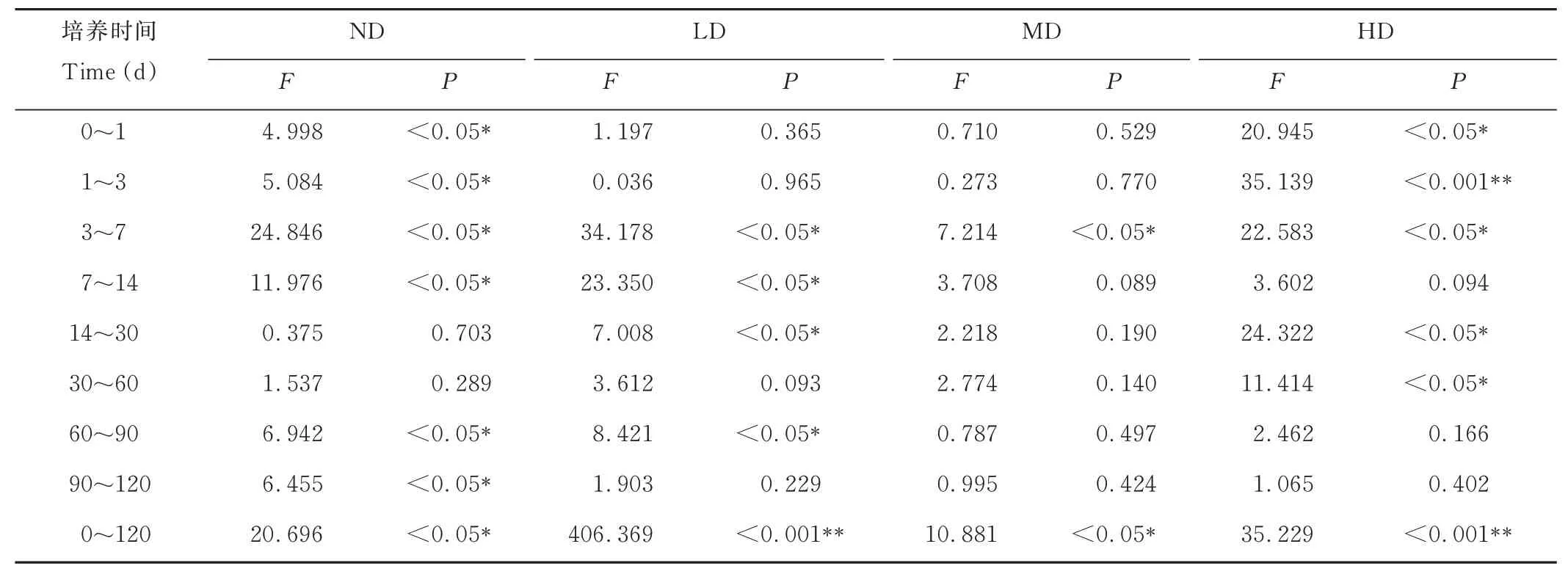
表3 同一类型草甸土壤有机磷净矿化速率在温度间的方差分析结果Table 3 Results of variance analysis of net mineralization rate of organic phosphorus in the same meadow soil among temperatures

图3 土壤有机磷净矿化速率动态Fig. 3 Dynamic of net mineralization rate of soil organic phosphorus
表4 显示,5、10 ℃培养下的每个时间段,土壤RNPM在4 类草甸间的差异不显著(P>0.05),其中120 d 培养期内土壤RNPM大小为ND≈LD>MD≈HD。15 ℃时,0~90 d 内4 类草甸土壤的RNPM均无显著差异(P>0.05);90~120 d 时,与ND 相比,LD、MD、HD 土壤的RNPM分别减少16.9%、52.8%、56.8%,其中ND 与MD、HD 的RNPM差异显著(P<0.05);120 d 培养期,4 类草甸土壤的RNPM均为正,且ND 的RNPM为0.66 mg·kg-1·d-1,分别比LD、MD和HD 显著高11.7%、48.2%、49.5%,LD 的还比MD 和HD 显著高41.3%、42.7%。这说明高寒草甸退化降低了土壤有机磷的净矿化速率,且降低幅度随着温度的升高而增加(图3)。

表4 同一温度下不同类型草甸土壤有机磷净矿化速率的方差分析结果Table 4 Results of variance analysis of net mineralization rate of organic phosphorus in different meadow soils at the same temperature
2.3 土壤有机磷矿化动力学特征
如图2 所示,土壤CPM 在5 和10 ℃培养下的0~7 d 降低、7~120 d 升高,即存在减少和增加2 个明显阶段;在15 ℃培养下120 d 内整体呈波浪形递增,基本符合扩散方程模型。因此,在选择常用的抛物线模型[23]模拟土壤有机磷矿化的动态过程时,5、10 ℃划分为2 个时段0~7 d 和7~120 d。
由表5 可知,4 类草甸土壤有机磷矿化动态的抛物线扩散拟合方程的相关系数都达显著水平(P<0.05),表明抛物线扩散模型能很好地模拟草甸土壤有机磷矿化的动态过程。从5~10 ℃的第二阶段和15 ℃看,每类草甸土壤有机磷矿化动力方程的A0值,都随温度升高而显著增加(P<0.05);同一培养温度下,A0值仅在15 ℃时因草甸类型不同而差异显著,LD、MD、HD 的土壤A0值分别较ND 增加8.0%、3.1%、15.1%,其中ND 与LD、HD 间的差异显著(P<0.05)。每类草甸土壤的B0值在5~15 ℃范围内,均随温度的升高逐渐增加,其中ND 的B0值在15 ℃时相较于5 ℃显著增加34.9%(P<0.05)。5~10 ℃下,B0值虽然在4 类土壤间无显著差异(P>0.05),但随草甸退化程度加剧逐渐减小,15 ℃下ND 的B0值分别比LD、MD、HD 高17.0%、41.9%、67.8%,其中ND 与MD、HD 差异显著(P<0.05)。说明高寒草甸退化减小了土壤有机磷的矿化作用,且在温度较高(≥15 ℃)时,中度与重度退化草甸土壤有机磷矿化作用显著减弱。

表5 土壤有机磷矿化动力学方程的模拟参数Table 5 Simulation parameter of dynamic equation (CPM=A0+ B0t1/2) of soil organic phosphorus mineralization
2.4 土壤有机磷矿化的温度效应
表6 显示,二次函数模型可以较好地描述高寒草甸土壤有机磷矿化速率与温度的关系,4 类土壤拟合方差的R2>0.78,P<0.05。LD、MD、HD 的C值分别比ND 降低58.8%、138.6%、159.7%,y″值分别较ND 降低24.6%、68.9%、75.4%,其中C和y″值在ND 与MD、HD 间差异显著(P<0.05)。说明高寒草甸退化降低了土壤有机磷矿化速率对温度的敏感性,且在草甸中度和重度退化条件下降低幅度达显著水平。

表6 土壤有机磷矿化速率与温度的关系Table 6 Regression relationship between soil organic phosphorus mineralization rate and temperature
3 讨论
3.1 高寒草甸退化对土壤有机磷矿化的影响
土壤有机磷矿化是磷循环的重要过程,能反映土壤供磷潜力[24]。本研究中随土壤培养时间的延长,4 种不同程度高寒草甸土壤有机磷净矿化速率都在初期呈较低的负值,之后增加并逐渐趋于稳定,与林地和草原草地土壤有机磷矿化过程相似[25-26]。究其原因是在培养前期土壤碳氮等养分充足,微生物活性强,发生磷的生物固定作用[27];中期解磷微生物开始分解土壤中易分解的有机磷和死亡的微生物残体[28],但后期养分不断消耗,微生物数量与活性逐渐下降至一个动态平衡状态[27],导致土壤有机磷矿化趋近稳定。
本研究显示草甸退化降低了土壤有机磷的累积矿化量、净矿化速率与矿化动力学拟合参数B0值,即减小了有机磷矿化作用。然而,不同草地土壤有机磷矿化作用的差异主要是由植被盖度、凋落物输入量(近似等于地上生物量)与土壤有机质等的差异引起[26,29],本研究中生长季平均气温下(15 ℃)土壤有机磷矿化作用和大多数植被、土壤因子呈显著相关的结果与其相似(图4)。这是因为草地退化一方面会降低植被盖度和凋落物输入量,另一方面可能限制植物对营养元素的重吸收效率[30],降低进入土壤的凋落物质量和分解效率。这两方面的共同作用导致土壤有机物和养分的输入量减少,加之随着土壤容重的增加(表2),土壤透气性降低,土壤微生物可获得的碳氮源减少,微生物活性和磷酸酶活性因此降低[31-32],致使退化草甸土壤有机磷矿化能力减弱,且这一减弱作用在15 ℃培养下最为明显。同时,土壤有机磷矿化的减弱作用随草甸退化程度加强而增大。当草甸中度和重度退化时,植被盖度、凋落物输入量和土壤有机质、碳氮都显著低于未退化和轻度退化草甸(表2),很可能减少解磷微生物如蜡状芽孢杆菌、巨大芽孢杆菌的数量与活性[28],最终致使土壤有机磷矿化作用显著低于未退化与轻度退化草甸。

图4 土壤CPM 和B0值与土壤和植被因子的相关性Fig. 4 Correlation of soil CPM and B0 value with soil and vegetation factors
3.2 高寒草甸退化下温度对土壤有机磷矿化的影响
温度是影响土壤有机磷矿化的主要因素之一,一般温度升高土壤有机磷矿化进程也会加快[33]。若尔盖高原的4 类草甸土壤在5~15 ℃培养条件下,有机磷的累积矿化量、净矿化速率以及矿化动力学拟合参数B0都显示有机磷矿化作用随温度增加而增大,与阿拉斯加寒带苔原土壤有机磷矿化的增温试验结果一致[34]。这是因为一方面温度升高可以提高微生物活性,促进解磷微生物分泌胞外酶(磷酸酶)和有机酸,然后有机磷被水解或者由微生物产生的酶作用下进行脱磷酸反应,进而促进有机磷的矿化[28,35];另一方面温度升高能增强土壤磷酸酶活性,从而促使有机磷矿化[28,36]。但是,Menge 等[37]发现温度升高对美国加州草原草地土壤有机磷矿化没有显著影响,这可能是该研究区属地中海气候,土壤含水量较低(<35%),微生物活性弱,使得水分而不是温度成为有机磷矿化的关键驱动因子。此外,Menge 等[37]的试验设计仅包括2 个温度,且温差只有1 ℃,设计温度较少和温差过小可能也是导致增温对土壤有机磷矿化率没有显著影响的原因之一。
李靖雯[38]报道温度对不同杉龄土壤有机磷矿化的影响有明显差异,本研究也发现不同退化程度草甸土壤有机磷矿化作用虽在5 和10 ℃下差异较小,但在15 ℃下却差异显著,主要表现为中度与重度退化草甸土壤有机磷矿化作用显著小于未退化与轻度退化草甸。这很可能是因为微生物和磷酸酶活性受温度影响[39],如尹亚丽等[40]发现当环境温度增高时,高寒草甸土壤微生物活性和群落多样性显著高于其他低温时期;高亚君等[41]发现春季和夏季温度高,高寒草甸土壤磷酸酶活性均高于低温时的冬季。因此,低温下(5 和10 ℃)土壤有机磷累积矿化量、B0值和土壤、植被因子均无显著相关性,此时微生物和磷酸酶活性较低,不利于土壤有机磷的矿化[39],进而减小了土壤有机磷矿化作用在不同退化程度草甸间的差异。随着温度的升高(≥15 ℃),有机磷矿化与多个土壤、植被因子出现了显著相关性(图4),这是因为15 ℃时大量凋落物输入的土壤有机磷源多,含解磷菌在内的微生物与磷酸酶也因适宜的温度、良好的土壤性质呈现数量多、活性高等特征[31-32],这样凋落物输入、有机质与氮含量大幅降低的中度与重度退化草甸因土壤微生物和磷酸酶活性的增强效果劣于未退化与轻度退化草甸,最终导致土壤有机磷矿化作用相比未退化与轻度退化草甸显著减小。正是由于上述凋落物输入、有机质与氮含量差异引起的土壤有机磷和微生物特性的不同,导致高寒草甸退化也降低了土壤有机磷矿化的温度敏感性,温度敏感性又能定量反映土壤有机磷矿化对气候变化的响应效应[42]。因此,全球气候变暖很可能会加剧土壤有机磷矿化能力因高寒草甸退化减弱的趋势,尤其是减小中度与重度退化草甸土壤有机磷矿化的温度敏感性,可能进一步引发退化草甸生态系统的磷限制问题。
4 结论
高寒草甸退化降低了土壤有机磷矿化作用,在15 ℃培养条件下降低作用尤为明显,其中120 d 培养期内,中度与重度退化草甸土壤有机磷的净矿化速率与动力学参数B0值均较未退化草甸显著降低。土壤有机磷矿化的温度敏感性因草甸退化程度加剧而逐渐降低,温度敏感系数在中度、重度退化草甸与未退化草甸间的差异达显著水平。因此,高寒草甸在中度与重度退化时显著降低了土壤有机磷矿化作用及其对温度的敏感性,可能会在全球气候变暖背景下加剧磷对退化高寒草甸初级生产力的限制作用。建议重点对中度与重度退化草甸实施轮牧甚至禁牧,控制鼠害,选育富集磷的土著草本植物,适当配施无机速效磷肥,提高土壤供磷潜力。
