酶菌协同发酵改善菜籽饼粕营养价值的研究
2023-10-16邓生青郭诚诺韩明霞王钰明解竞静萨仁娜
邓生青 郭诚诺 韩明霞 王钰明 赵 峰 解竞静 萨仁娜
(中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193)
我国家禽年出栏量居世界第一,长期以来以玉米-豆粕型饲粮为主,这些原料大量依赖进口,使得饲料成本逐年升高[1]。为了缓解养殖业饲料原料的供需矛盾,农业农村部实施“豆粕减量替代”行动,引导养殖企业以杂粕类原料替代豆粕。因此,利用生物技术手段提高杂粕类原料的消化利用率,开发利用不同地区的原料,缓解进口豆粕的压力,具有重要意义。菜籽饼粕是油菜籽榨油加工后的副产物,在2020—2021年产量约1 000万t,是产量最高的杂粕类蛋白质饲料原料[2],其粗蛋白质(CP)含量略低于豆粕,含有丰富的含硫氨基酸,赖氨酸和精氨酸含量略低[3]。由于普通菜籽饼粕含有硫甙(Gls)、芥子碱、植酸、单宁和纤维等抗营养因子,适口性差,消化率低于豆粕[4]。研究发现,饲粮中添加10%的菜籽粕显著降低了肉鸡的生长性能[5]。菜籽饼粕的纤维主要存在于菜籽壳上,是难以被家禽利用的结构性多糖(纤维素、果胶多糖、木质素、半纤维素等),彼此间通过氢键与木葡聚糖相互连接形成刚性网状结构,并将蛋白质等养分缠绕包埋。菜籽饼粕中的大部分纤维不能被畜禽体内消化酶降解,也不能被上消化道吸收[6]。Gls是菜籽饼粕毒理作用最大的抗营养因子,可引起甲状腺肿,导致生长迟缓[7]。
目前通过发酵技术可以消除Gls、单宁等抗营养因子[8],但对纤维的降解作用不显著[9]。Olukomaiya等[10]报道了发酵后菜籽粕的中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)等含量出现升高的现象。研究发现,利用酶解技术可以降解菜籽饼粕中的纤维并增加产物中小肽含量,但对Gls几乎无作用[11-12]。而酶菌协同发酵杂粕可将酶解技术和发酵技术有效结合,从而提高饲料利用率[8,13]。Goodarzi等[14]研究发现,在菜籽饼液态自发发酵过程中加入果胶酶、葡聚糖酶和植酸酶后,可降低结构性多糖和植酸含量,并显著提高了肉鸡生长性能。不同产地不同加工工艺生产的菜籽饼粕,所含有的养分和抗营养因子含量差异较大[15-16],利用微生物发酵技术改善菜籽饼粕的营养价值受菜籽饼粕的性质、来源、榨油的加工工艺、储存条件、发酵菌种的选择、固体发酵的条件等因素影响[17-20],如发酵后菜籽饼粕CP含量升高[18,20]或无变化[10],NDF含量降低[18]或升高[10]。由此可见,菜籽饼粕生产条件、发酵菌种和工艺不同,对菜籽饼粕的改善效果差异很大。目前尚缺乏发酵工艺对不同来源的菜籽饼粕是否具备普遍适用性的研究。因此,本研究以不同来源的菜籽饼粕为底物,开展酶菌协同发酵,分析探讨发酵工艺对不同来源菜籽饼粕的改善作用,为菜籽饼粕的有效利用提供数据与理论参考。
1 材料与方法
1.1 试验材料与试剂
菌种:枯草芽孢杆菌(B.subtilis)购自中国工业微生物菌种保藏管理中心,植物乳杆菌(L.plantarum)为动物营养学国家重点实验室保存菌株。
酶源:纤维素酶,活性20 000 U/g;果胶酶,活性30 000 U/g;α-半乳糖苷酶,活性5 000 U/g。以上酶源均由天津博菲德科技有限公司惠赠。
主要原料:工艺优化用菜籽饼产自天津;另根据不同产地从我国不同区域采集了8个菜籽粕和1个菜籽饼,粉碎过0.42 mm筛备用。
培养基:MRS和LB培养基购自北京奥博星生物技术有限责任公司。
1.2 试验设计
试验于2021年12月至2022年5月在中国农业科学院北京畜牧兽医研究所动物营养学国家重点实验室进行。试验共分为2个部分。试验1:以工艺优化用菜籽饼为发酵底物,以发酵后菜籽饼的NDF降解率(%)、Gls降解率(%)、酸溶蛋白(TCA-SP)增加率(%)为评价指标,以总变化率(总变化率=0.7×NDF降解率+0.15×Gls降解率+0.15×TCA-SP增加率)为综合评价指标,确定酶菌协同最优发酵工艺,并进行发酵效果的验证。试验2:在最佳酶菌协同发酵工艺下对菜籽粕1、2、3、4、5、6、7、8及菜籽饼9进行酶菌协同发酵处理,探讨酶菌协同发酵工艺对不同来源菜籽饼粕的纤维降解效果及养分的改善效果。
1.3 试验方法
1.3.1 菌种发酵液制备
将枯草芽孢杆菌和植物乳杆菌分别在LB和MRS培养基划线培养后,挑取单菌落,分别接种于LB、MRS液体培养基中,在37 ℃、200 r/min条件下培养24 h,按1%的接种量分别接种到含100 mL液体培养基的三角瓶中,培养18 h后备用。
1.3.2 发酵过程
准确称取50.0 g菜籽饼置于500 mL三角瓶中,将菌种发酵液以枯草芽孢杆菌∶植物乳杆菌=5∶1的接种比例,按照12%的接种量接种至菜籽饼培养基中,加入蒸馏水调整料水比为1.0∶1.2,搅拌均匀后用无菌透气过滤封口膜封口,在35 ℃条件下恒温静置发酵96 h。发酵结束后于55 ℃低温烘干,粉碎待测。
1.4 酶菌协同发酵工艺优化
以总变化率为综合评价指标,以纤维素酶、果胶酶和α-半乳糖苷酶不同添加梯度为试验因素,设计L9(34)三因素三水平正交试验,试验设计见表1。
1.5 测定指标及方法
用于测定纤维含量的样品粉碎过0.84 mm筛,用于测定CP和TCA-SP含量的样品粉碎过0.42 mm筛,用于测定氨基酸和Gls含量的样品粉碎过0.25 mm筛。对工艺优化用菜籽饼发酵前后的pH及CP、TCA-SP、NDF、ADF、Gls和氨基酸含量进行测定,以及菜籽饼发酵前后扫描电镜分析和蛋白质分子质量分析。对9个不同来源菜籽饼粕发酵前后的pH及CP、TCA-SP、NDF和Gls含量进行测定。
1.5.1 养分测定
Gls含量采用氯化钯比色法[21]测定,DM含量通过105 ℃烘箱干燥4 h测定,CP含量采用凯氏定氮法测定,TCA-SP含量参照NY/T 3801—2020[22]测定,NDF含量参照GB/T 20806—2006[23]采用Van Soest分析法测定,ADF含量参照NY/T 1459—2007[24]采用Van Soest分析法测定,氨基酸含量利用氨基酸自动分析仪测定。
1.5.2 pH测定
称取发酵前后的样品5.0 g,加蒸馏水50 mL,搅拌30 min后,用预先校正过的pH计测定其pH。
1.5.3 电镜扫描分析
取适量样品粉碎过0.25 mm筛后,置于导电胶固定在扫描电镜专用样品台上,用磁控溅射铂金进行喷金处理,再将处理好的样品用扫描电镜(SU3500,日本)进行观察并拍照留存。
1.5.4 蛋白质分子质量分析
将样品碎过0.25 mm,取适量样品采用变性结合加热的方法来提取菜籽饼粕蛋白质并进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)分析,具体操作参照文献[25]进行。
1.6 数据统计与分析
采用Excel 2010软件对数据进行初步处理,采用SAS 9.4软件对数据进行单因素方差分析(one-way ANOVA),采用Duncan氏法进行多重比较,以P<0.05作为差异显著性判断标准。
2 结 果
2.1 不同来源菜籽饼粕化学成分
由表2可知,菜籽饼9的CP、NDF和TCA-SP含量分别为41.31%、40.39%和2.61%。菜籽粕1~8的CP、NDF和TCA-SP含量分别在37.07%~44.93%、27.03%~40.22%和2.11%~3.19%;CP的变异系数为6.17%,NDF和TCA-SP的变异系数均超过了10%。菜籽饼9的Gls含量为18.81 μmol/g。菜籽粕1~8的Gls含量在18.10~57.64 μmol/g,Gls的变异系数为38.67%,Gls含量最高的是产地为安徽的菜籽粕6,Gls含量最低的是产地为福建的菜籽粕5。
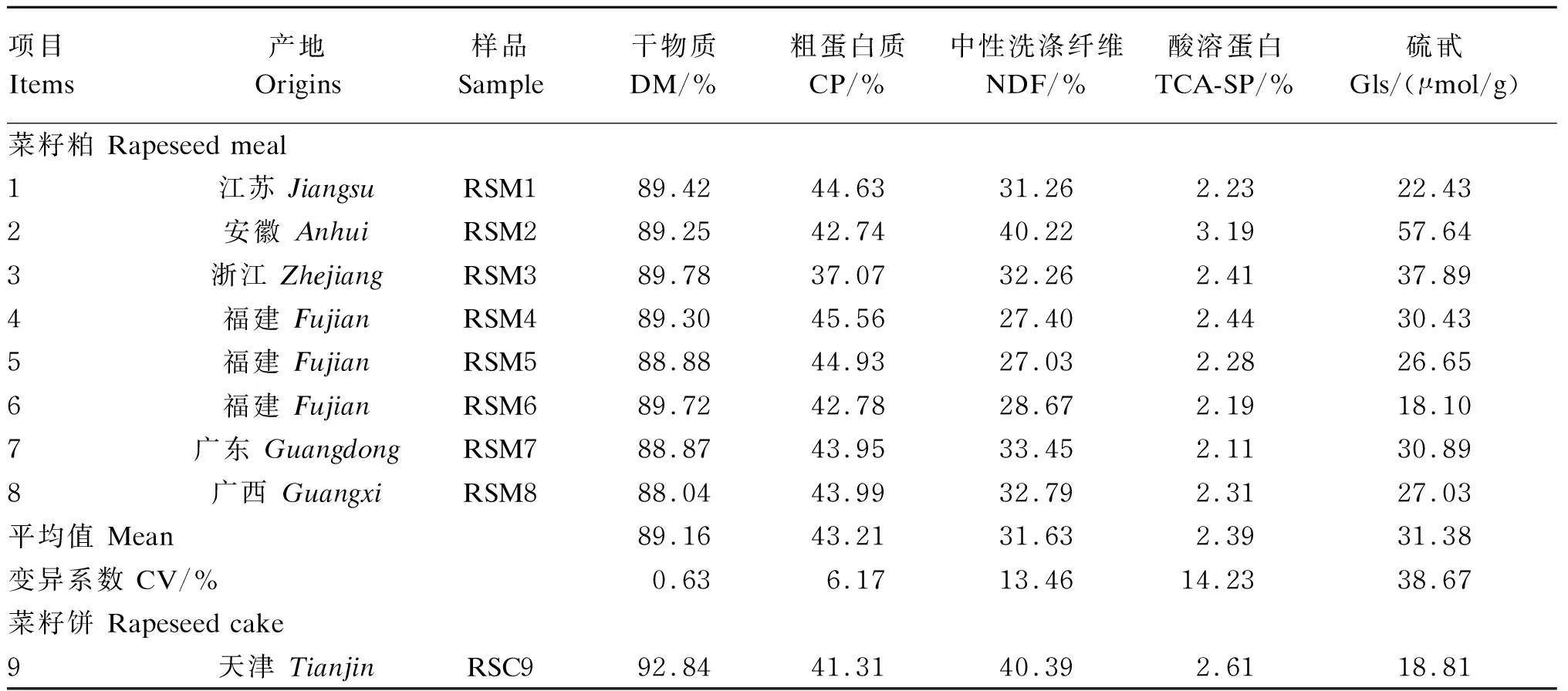
表2 不同来源菜籽饼粕化学成分(干物质基础)
2.2 酶菌协同发酵菜籽饼的正交试验结果
由表3可知,从极差结果来看,果胶酶对总变化率的影响最大,纤维素酶次之,α-半乳糖苷酶的影响最小,即果胶酶>纤维素酶>α-半乳糖苷酶,最优配比组合为纤维素酶∶果胶酶∶α-半乳糖苷酶=3∶3∶3,此时总变化率为46.44%。
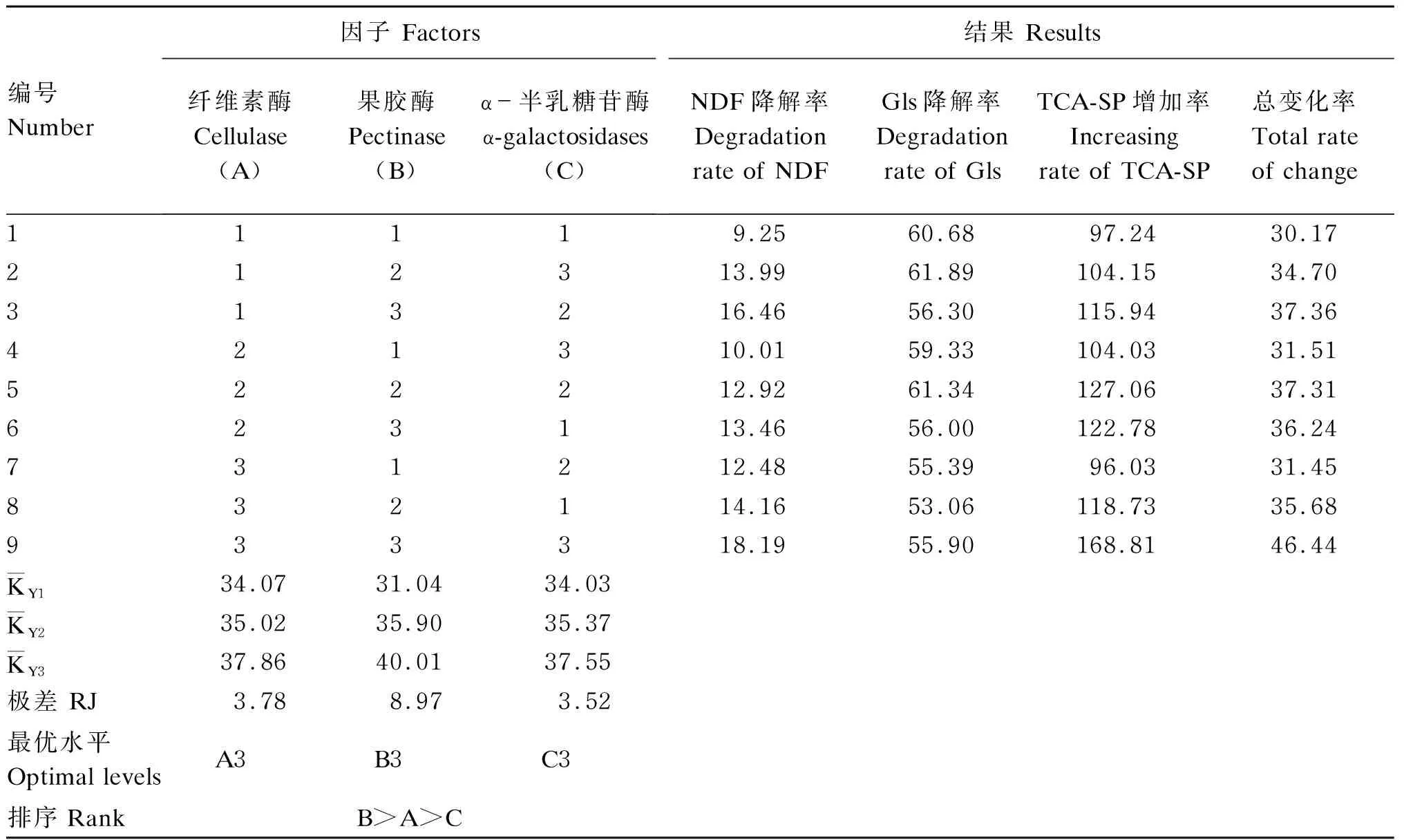
表3 酶菌协同发酵菜籽饼的正交试验结果(干物质基础)
2.3 酶菌协同发酵对菜籽饼pH及化学成分的影响
由表4可知,在最优配比组合酶菌协同发酵条件下,菜籽饼发酵后的pH显著低于发酵前(P<0.05),CP、TCA-SP含量显著高于发酵前(P<0.05),NDF、ADF、半纤维素、Gls含量显著低于发酵前(P<0.05),谷氨酸、丙氨酸、缬氨酸、亮氨酸及总氨基酸含量显著高于发酵前(P<0.05)。
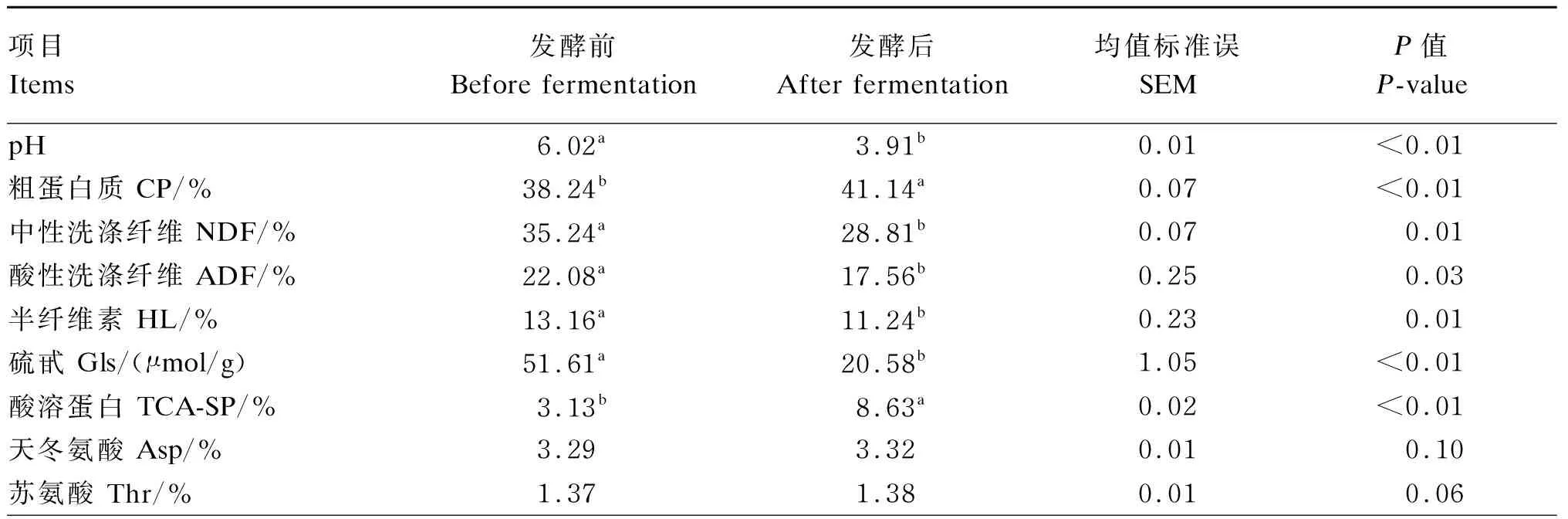
表4 酶菌协同发酵对菜籽饼pH及化学成分的影响(干物质基础)
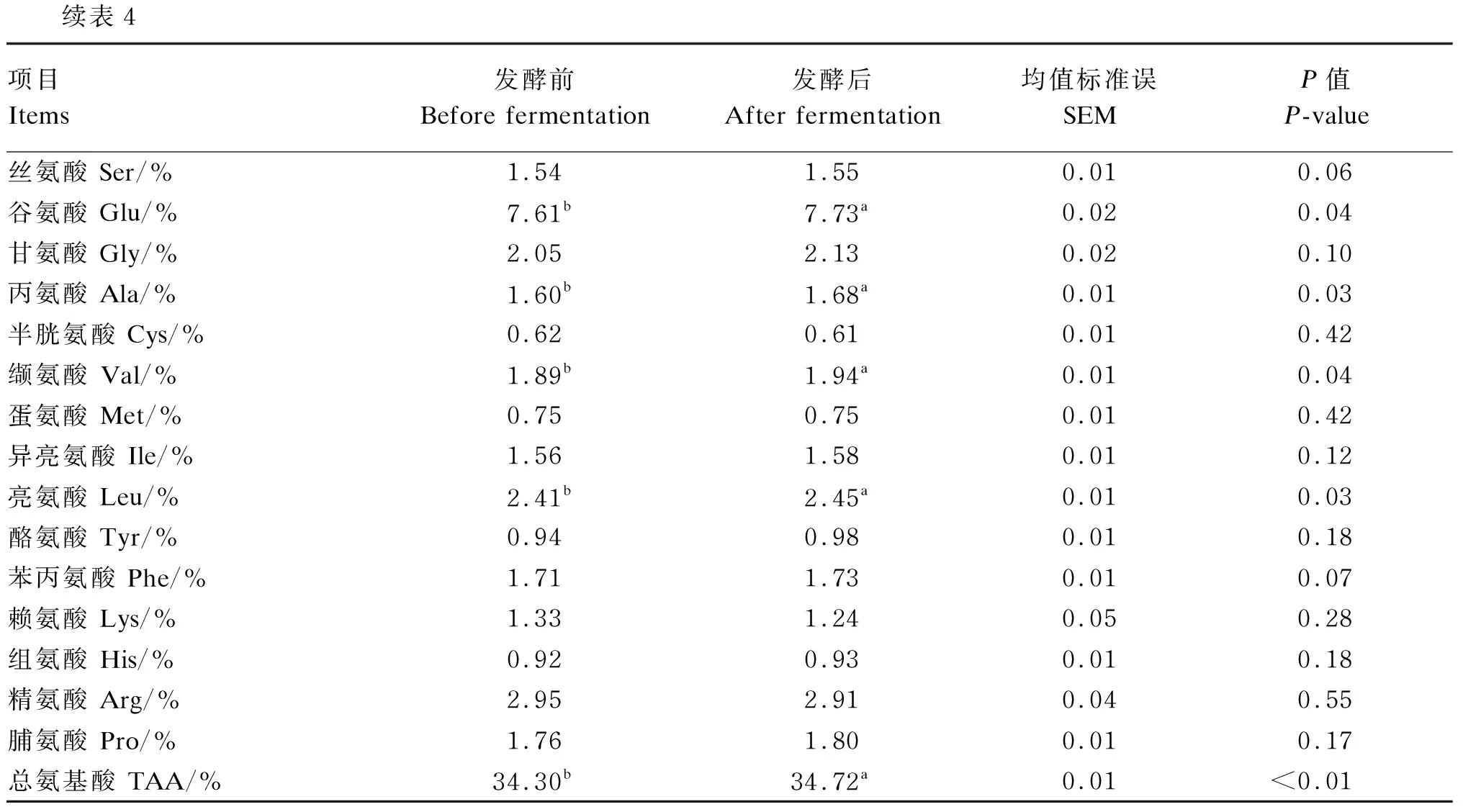
续表4项目Items发酵前Before fermentation发酵后After fermentation均值标准误SEMP值P-value丝氨酸 Ser/%1.54 1.550.010.06谷氨酸 Glu/%7.61b 7.73a 0.020.04甘氨酸 Gly/%2.05 2.13 0.020.10丙氨酸 Ala/%1.60b 1.68a 0.010.03半胱氨酸 Cys/%0.62 0.61 0.010.42缬氨酸 Val/%1.89b 1.94a 0.010.04蛋氨酸 Met/%0.75 0.75 0.010.42异亮氨酸 Ile/%1.56 1.58 0.010.12亮氨酸 Leu/%2.41b 2.45a 0.010.03酪氨酸 Tyr/%0.94 0.98 0.010.18苯丙氨酸 Phe/%1.71 1.73 0.010.07赖氨酸 Lys/%1.33 1.24 0.050.28组氨酸 His/%0.92 0.93 0.010.18精氨酸 Arg/%2.95 2.91 0.040.55脯氨酸 Pro/%1.76 1.80 0.010.17总氨基酸 TAA/%34.30b 34.72a 0.01<0.01
2.4 酶菌协同发酵对菜籽饼表面形态的影响
在最优配比组合酶菌协同发酵条件下,电镜观察发酵前后菜籽饼表面形态见图1。在400倍电镜观察下,发酵前的菜籽饼颗粒大,形态不规则;发酵后的菜籽饼颗粒较小,结构较松散。在3 000倍电镜观察下,发酵前的菜籽饼表面光滑平整,呈较大的片状,无小孔隙且结构较为完整;发酵后的菜籽饼表面粗糙,呈现不规则分布,孔隙较多。
2.5 酶菌协同发酵对菜籽饼蛋白质分子质量的影响
在最优配比组合酶菌协同发酵条件下,发酵前后菜籽饼的蛋白质分子质量结果见图2。A区域蛋白质分子质量为30~75 ku,B区域蛋白质分子质量为12~23 ku,发酵前菜籽饼的蛋白质在A区域分布有多个条带,发酵后菜籽饼的蛋白质主要分布在B区域。
2.6 酶菌协同发酵对不同来源菜籽饼粕养分含量的影响
在最优配比组合酶菌协同发酵条件下,对9个不同来源菜籽饼粕进行酶菌协同发酵。
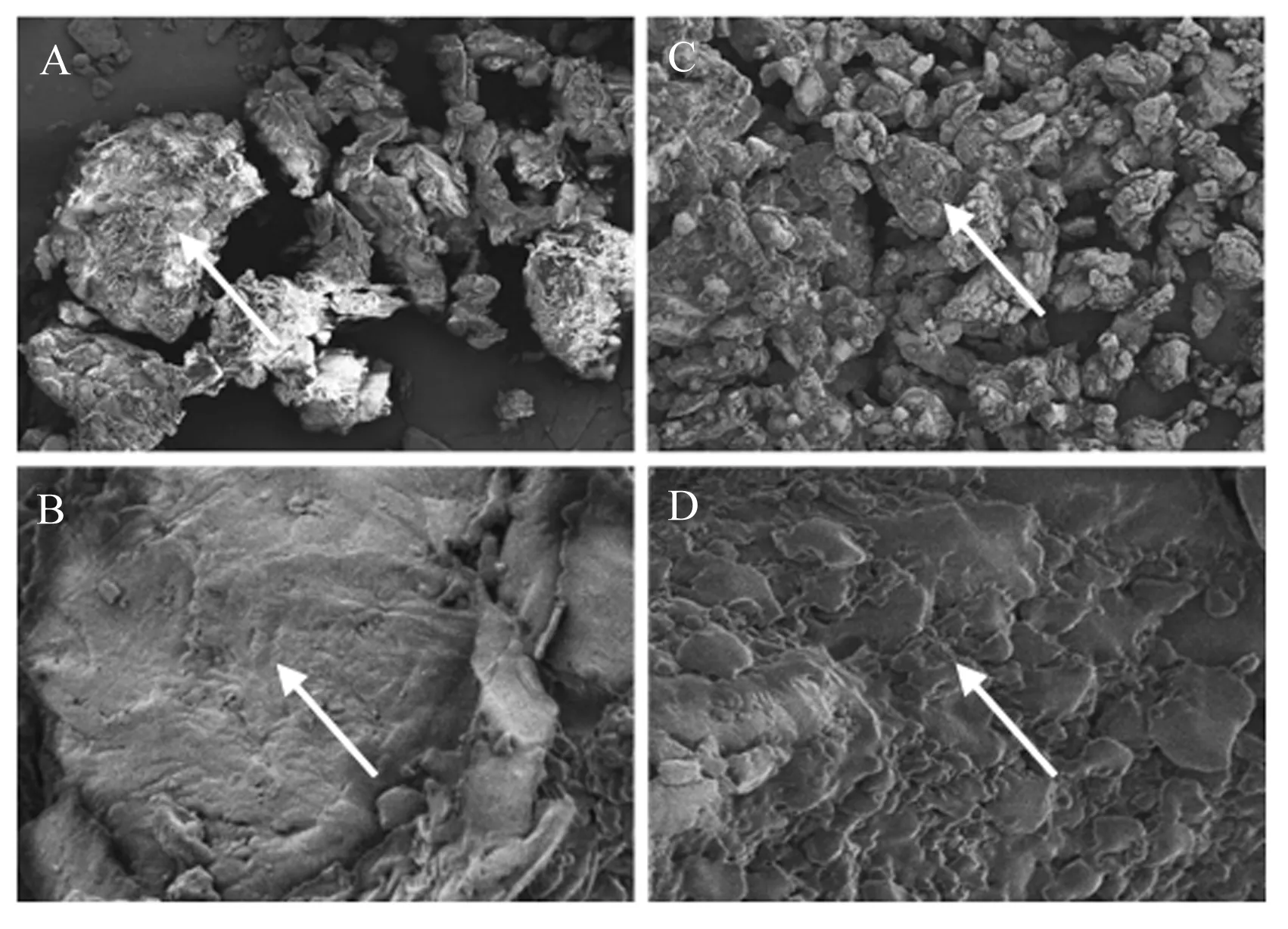
A:发酵前(400×);B:发酵前(3 000×);C:发酵后(400×);D:发酵后(3 000×)。
2.6.1 pH及CP、TCA-SP含量
由表5可知,发酵前,9个菜籽饼粕的pH在5.46~5.95,平均值为5.75;发酵后,9个菜籽饼粕的pH在3.92~4.27,平均值为4.06。9个菜籽饼粕发酵后的pH均显著低于发酵前(P<0.05),其中变化幅度最高的为菜籽饼9,降低幅度为1.98。
由表6可知,发酵前,9个菜籽饼粕的CP含量在37.07%~45.56%,平均值为43.00%;发酵后,9个菜籽饼粕的CP含量在38.85%~46.15%,平均值为43.57%。菜籽粕1、3及菜籽饼9发酵后的CP含量显著高于发酵前(P<0.05)。发酵前,9个菜籽饼粕的TCA-SP含量在2.11%~3.19%,平均值为2.42%;发酵后,9个菜籽饼粕的TCA-SP含量在5.35%~6.52%,平均值为5.95%。9个菜籽饼粕发酵后的TCA-SP含量显著高于发酵前(P<0.05),其中变化幅度最高的为菜籽粕1,提高幅度为192.38%。
2.6.2 NDF和Gls含量
由表7可知,发酵前,9个菜籽饼粕的NDF含量在27.03%~40.39%,平均值为32.61%;发酵后,9个菜籽饼粕的NDF含量在20.62%~34.76%,平均值为26.08%。9个菜籽饼粕发酵后的NDF含量显著低于发酵前(P<0.05),其中变化幅度最高的为菜籽粕7,降低了26.00%;变化幅度最低的是菜籽饼1,降低了13.95%。
由表8可知,发酵前,9个菜籽饼粕的Gls含量在18.10~57.64 μmol/g,平均值为29.99 μmol/g;发酵后,9个菜籽饼粕的Gls含量在13.62~46.23 μmol/g,平均值为22.80 μmol/g。9个菜籽饼粕发酵后的Gls含量显著低于发酵前(P<0.05),其中变化幅度最高的为菜籽粕1,降低了31.30%;变化幅度最低的是菜籽粕4,降低了16.20%。
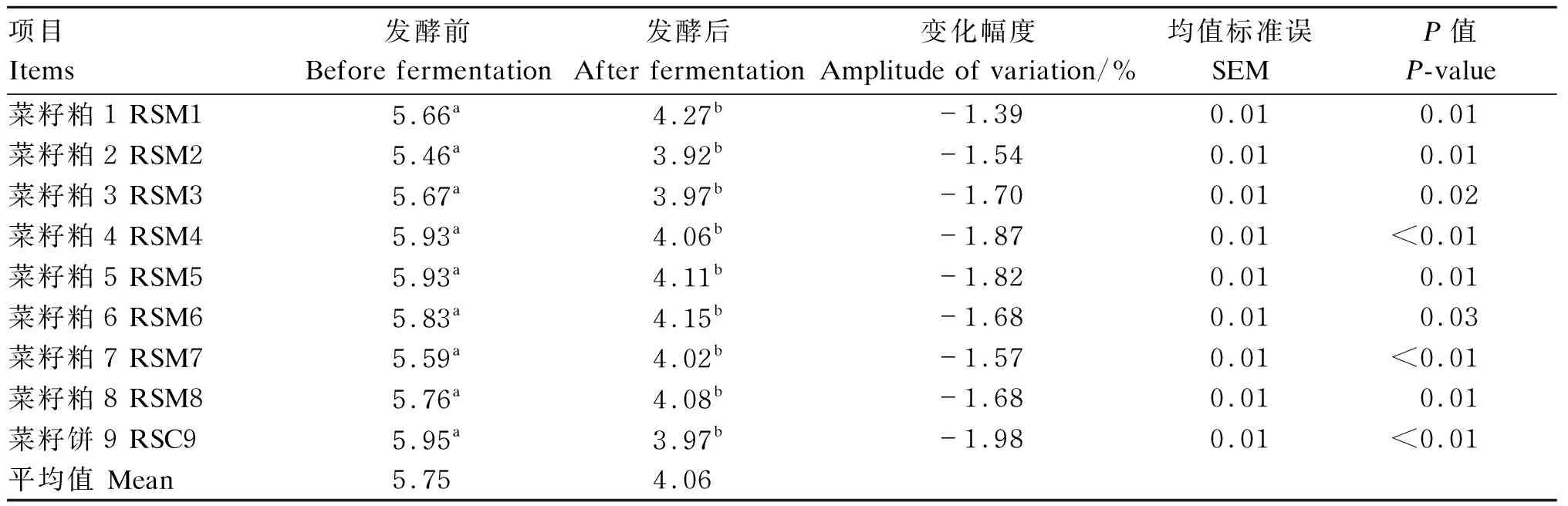
表5 酶菌协同发酵对不同来源菜籽饼粕pH的影响
3 讨 论
3.1 酶菌协同发酵对菜籽饼pH、化学成分及氨基酸含量的影响
本试验前期工作中发现,混菌发酵菜籽饼对NDF降解作用较小,部分处理出现NDF含量升高的现象,这可能是由于寡糖及Gls含量的降低所形成的“浓缩效应”[26];也可能是发酵后产生中性不溶洗涤物或酸性洗涤不溶物等,使NDF的表征值被高估了[27]。针对这一结果,本试验在混菌发酵体系中加了纤维素酶、果胶酶和α-半乳糖苷酶,采用酶菌协同发酵的方式,结果发现,NDF降解率、TCA-SP增加率及总变化率在纤维素酶、果胶酶和α-半乳糖苷酶添加量分别为60、60和10 U/g时最高,分别为18.25%、175.72%和60.12%。Zhu等[28]研究发现,用非淀粉多糖酶前处理再进行乳酸菌发酵,菜籽粕的NDF、ADF及半纤维素含量显著降低,而乳酸菌单独发酵后其含量没有显著变化。王卫卫[29]筛选出的可降解羧甲基纤维素钠的枯草芽孢杆菌,发酵后对棕榈仁粕纤维降解效果较小,在进行酶菌协同发酵后可显著降低棕榈仁粕的纤维含量。本试验结果与上述研究结果相近。
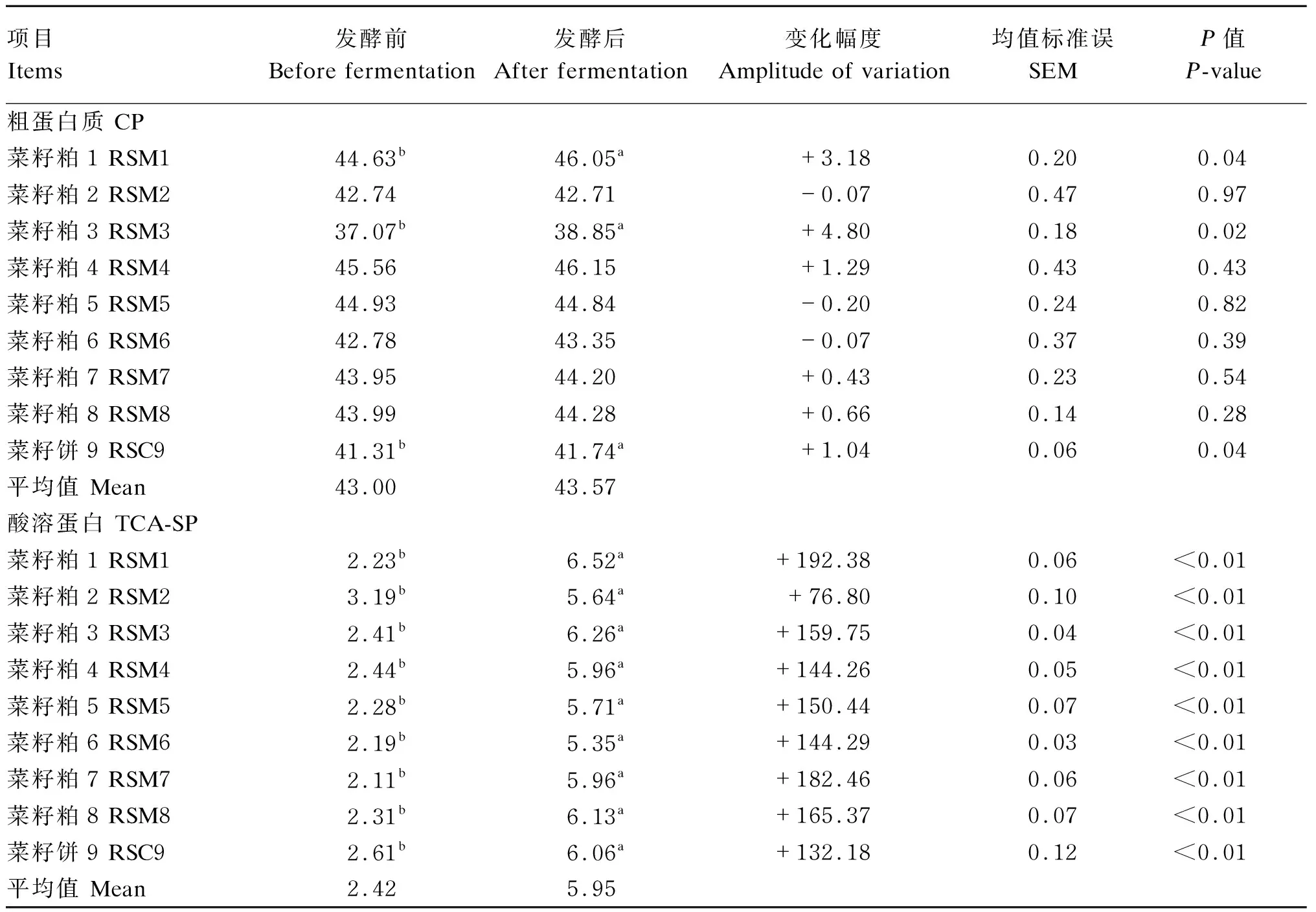
表6 酶菌协同发酵对不同来源菜籽饼粕CP和TCA-SP含量的影响(干物质基础)
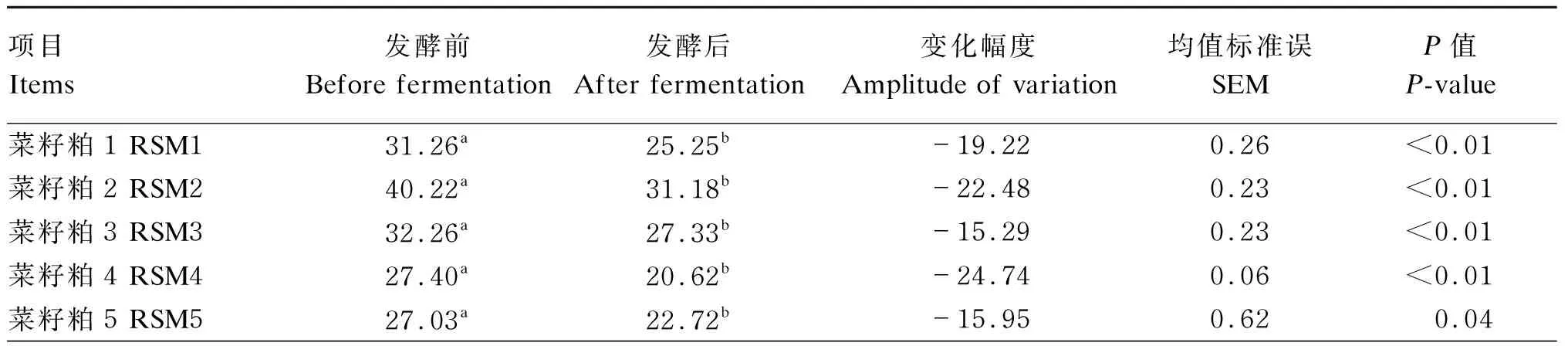
表7 酶菌协同发酵对不同来源菜籽饼粕NDF含量的影响(干物质基础)
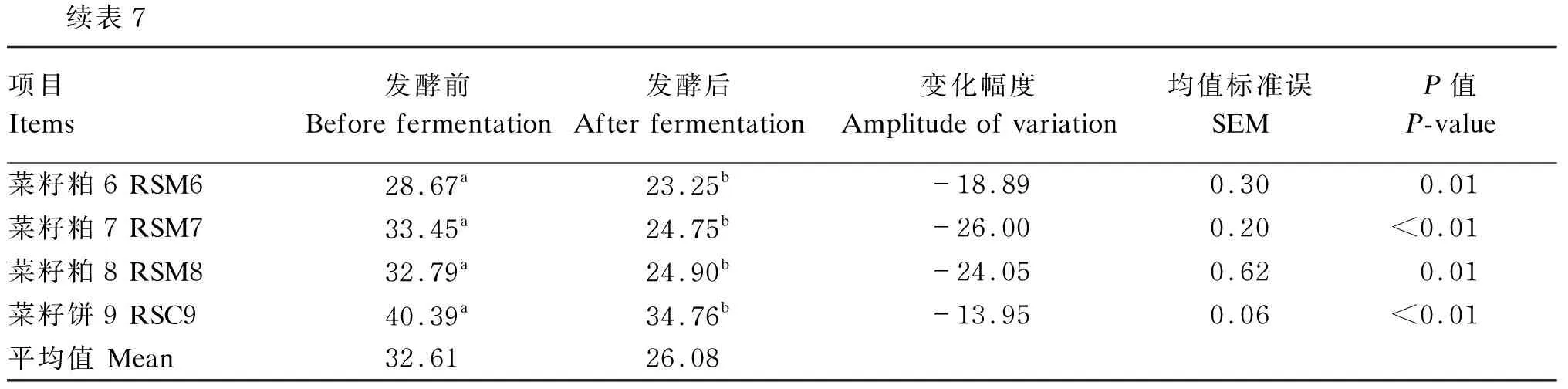
续表7项目Items发酵前Before fermentation发酵后After fermentation变化幅度Amplitude of variation均值标准误SEMP值P-value菜籽粕6 RSM628.67a23.25b-18.890.300.01菜籽粕7 RSM733.45a24.75b-26.000.20<0.01菜籽粕8 RSM832.79a24.90b-24.050.620.01菜籽饼9 RSC940.39a34.76b-13.950.06<0.01平均值 Mean32.6126.08
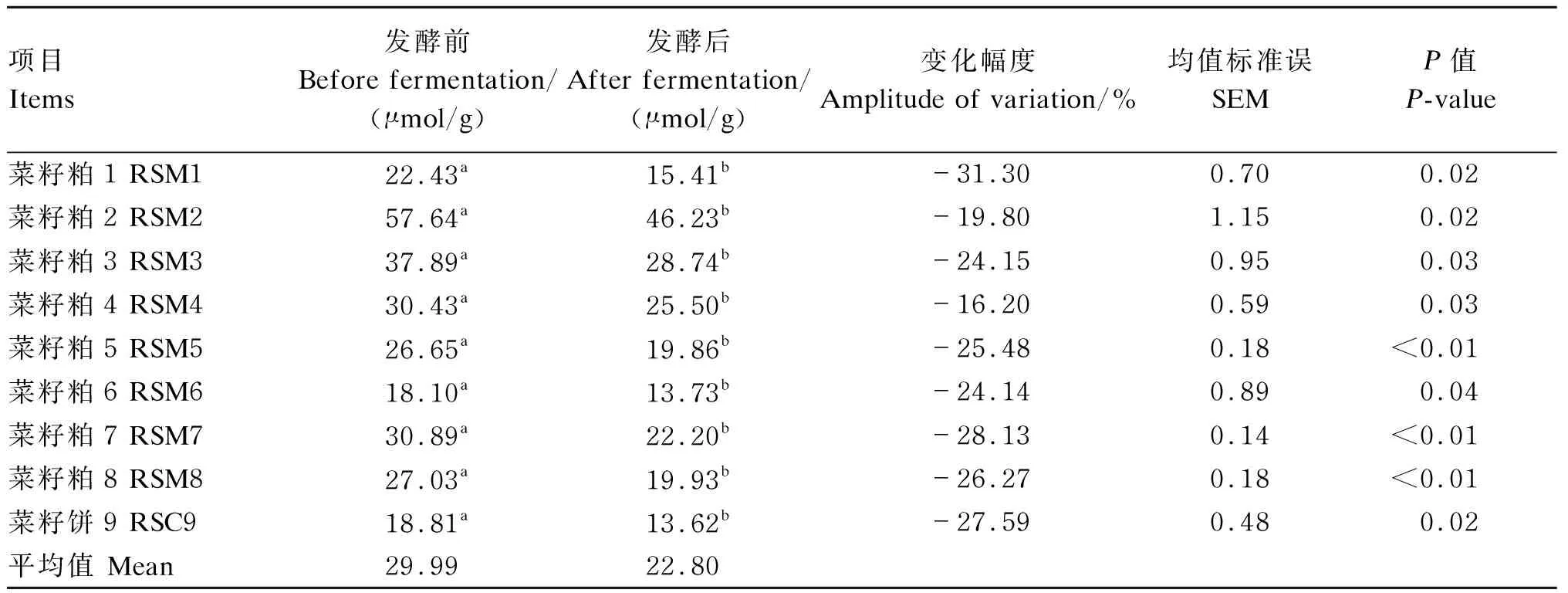
表8 酶菌协同发酵对不同来源菜籽饼粕Gls含量的影响(干物质基础)
本试验在最优配比组合酶菌协同发酵条件下进行酶菌协同发酵菜籽饼后,菜籽饼发酵后pH显著低于发酵前,CP含量显著高于发酵前。在菜籽饼发酵过程中微生物利用底物中的养分并形成菌体蛋白,并将菜籽饼中大分子蛋白质分解为小肽,也因“浓缩效应”使得发酵后CP含量升高。酶菌协同发酵菜籽饼后TCA-SP增加率为175.72%,TCA-SP主要是由氨基酸残基组成的寡肽,动物吸收寡肽的能力要高于游离的氨基酸[30]。本试验结果表明,酶菌协同发酵菜籽饼的纤维(NDF、ADF和半纤维素)含量均降低,这与Zhu等[28]的报道相似。酶菌协同发酵后Gls降解率为60.12%,表明所用菌株对Gls有较强的降解能力。朱晓峰等[11]用乳酸杆菌发酵后,菜籽粕Gls含量降低了43.43%。郝怡宁等[31]用枯草芽孢杆菌和雅致放射毛霉发酵后,菜籽粕Gls含量降低了45.26%。本试验所用菌株降解Gls能力高于上述文献菌株。
与豆粕相比,菜籽饼的氨基酸消化率较差[32]。本试验结果显示,菜籽饼总氨基酸含量较发酵前显著提高,但各氨基酸组成及含量变动较小。在郝怡宁等[31]的报道中,菜籽粕发酵3 d时总氨基酸含量显著高于未发酵时,然而,发酵5 d后各氨基酸及总氨基酸含量出现下降的趋势。本试验中,氨基酸含量的变化可能受到菌株及发酵时间的影响,在发酵过程中的具体变化趋势还需要进一步进行探讨。
3.2 酶菌协同发酵对菜籽饼表面结构和蛋白质分子质量的影响
扫描电镜结果表明,发酵前菜籽饼颗粒大形态不规则,经酶菌协同发酵后,颗粒变小,表面粗糙出现凹凸不平的孔状。这与Shi等[17]利用黑曲霉发酵菜籽粕的结果一致。菜籽饼完整细胞壁上的纤维网状结构会保护其他养分如蛋白质和多糖免于被降解或转化[33],当对菜籽饼进行堆肥发酵后,其纤维素网状结构大部分被破坏[34]。本试验结果显示,酶菌协同发酵后菜籽饼表面结构被破坏,使得表面积增加,在畜禽采食后,增加了肠道消化酶接触饲料的面积,更有利于消化饲料。
菜籽蛋白主要有2种类别,主要是大分子的12S球蛋白(α亚基和β亚基)和小分子的2S白蛋白,其中12S球蛋白的分子质量在30~60 ku,2S白蛋白分子质量约为6、12和21 ku[18]。本试验结果显示,未发酵菜籽饼主要为30~75 ku的大分子蛋白质,而酶菌协同发酵增加了小分子肽(<15 ku)的分布,这表明酶菌协同发酵在一定程度上将大分子蛋白质分解为小分子肽,进而增加菜籽饼小肽含量,这与Shi等[17]研究结果相似。与大分子蛋白质相比,小分子肽更易被家禽肠道所捕获吸收[18],有利于提高饲料养分消化率。
3.3 酶菌协同发酵对不同来源菜籽饼粕养分含量的影响
与豆粕相比,菜籽饼粕的纤维含量较高,含有Gls等抗营养因子,极大限制了菜籽饼粕在畜禽中的应用。而不同菜籽饼粕的营养价值与养分消化率因来源不同、榨油工艺不同而有差异[35]。油菜籽加工副产物可根据加工方式的不同分为高温热榨菜籽饼、低温冷榨菜籽饼以及压饼后浸提的菜籽粕,不同榨油过程中的温度处理生产的菜籽饼粕的养分和抗营养因子含量均有所不同[16]。Toghyani等[36]研究发现,菜籽粕NDF含量与肉鸡代谢能之间存在极显著的负相关关系。Eklund等[37]研究发现,猪的标准回肠氨基酸消化率随着菜籽粕中NDF等纤维含量的升高而显著降低。因此,提高菜籽饼粕的可消化蛋白质含量,降低Gls及纤维含量,是提高菜籽饼粕营养价值的重要手段。
本试验通过酶菌协同发酵,发现对9个不同来源的菜籽饼粕养分含量均有一定的改善作用。经过酶菌协同发酵后,菜籽饼粕的pH均显著降低。pH的降低可提高畜禽肠道内胃蛋白酶的活性,从而促进对养分的消化吸收[38]。经过酶菌协同发酵后,9个菜籽饼粕的CP含量略有升高,提高的幅度非常有限,在1%左右,说明本试验酶菌协同发酵改善菜籽饼粕的营养价值,对CP含量的提高并不明显,而发酵体系蛋白质含量的变化可能与菌体蛋白的产生有关。菌体蛋白产生量与发酵菌种有关,Wang等[18]利用乳酸菌和地衣芽孢杆菌发酵菜籽粕,CP含量提高了12.09%;Shi等[17]用黑曲霉发酵菜籽粕后,CP含量提高了16.90%。本试验采用的枯草芽孢杆菌具有分泌酸性蛋白酶的活性,可促进饲料大分子蛋白质的进一步降解,这与蛋白质分子质量的结果相一致的。本试验测定的9个菜籽饼粕的TCA-SP含量均有大幅度的提高,表明酶菌协同发酵对TCA-SP含量有明显的提高作用。如前所述,TCA-SP主要是由氨基酸残基组成的寡肽,动物吸收寡肽的能力要高于游离的氨基酸[30]。Tie等[20]研究发现,菜籽粕经蛋白酶酶解及黑曲霉发酵后TCA-SP含量增加了629.66%。本试验结果与上述结果相似。
菜籽饼粕的纤维含量高于豆粕3倍以上[39],且饲料原料纤维含量与代谢能存在负相关[36]。与饲喂玉米-豆粕基础饲粮相比,饲喂菜籽饼粕原料的肉鸡代谢能呈线性下降。相对较低的代谢能是限制菜籽饼粕在肉鸡中饲喂效果的主要因素,这通常与纤维含量高有关。本试验中,经过酶菌协同发酵后,9个菜籽饼粕的NDF含量均显著降低,表明本试验选用混合微生物并联合使用适宜的非淀粉多糖酶,均有一定降解9个不同来源菜籽饼粕NDF的效果。植物源性细胞壁结构错综复杂,纤维互相缠绕形成坚固的网状结构[4],而菜籽饼粕纤维主要是难以被家禽利用的结构性多糖[40],这种纤维网状结构会将蛋白质及淀粉等可消化养分进行包裹,不容易被分解利用[34]。而非淀粉多糖酶结合微生物进行酶菌协同发酵可对菜籽饼粕中纤维等抗营养因子或大分子结构更具针对性和特异性,这种策略延长了酶与底物的接触与作用时间,能同时提高微生物发酵效果和酶解的效率。有研究表明,细胞壁的纤维网状结构在纤维素酶、果胶酶的相互作用下得以打开并降解[41],此时微生物进入细胞内部发挥作用,将大分子化合物转变为小肽等小分子化合物,并利用纤维降解产生的可溶性糖转化为乳酸等代谢产物[42]。朱晓峰等[11]报道了在纤维素酶及果胶酶的酶解作用下,菜籽粕的NDF含量由38.12%降至18.68%;王卫卫[29]研究发现,用甘露聚糖、α-半乳糖苷酶协同植物乳杆菌和酵母菌发酵可显著降低棕榈仁粕的NDF含量,本试验结果与上述研究结果相似。
菜籽饼粕中的Gls可在芥子酶作用下分解为硫氰酸酯、异硫氰酸酯、噁唑烷硫酮及氰类化合物,可引起甲状腺肿,导致生长迟缓,故降低了畜禽生长性能[7]。本试验中,经过酶菌协同发酵后,9个菜籽饼粕的Gls含量显著降低。降低菜籽饼粕中抗营养因子Gls的含量,是双低菜籽饼粕育种技术的目标[39],也是采用发酵技术改善菜籽饼粕营养价值的主要目标之一[8],如利用枯草芽孢杆菌与毛菌混合发酵降解Gls[31],利用乳酸菌、酵母菌、地衣芽孢杆菌混合发降解Gls[18]。Gls含量的降低缓解了其对畜禽肠道、肝脏及甲状腺的损伤,进而提高畜禽对养分的消化吸收能力。此外,Gls含量的高低与生物发酵菌种有关,本试验结果表明选用酶菌协同发酵不同来源的菜籽饼粕,均可以起到降解Gls的效果。
4 结 论
① 在混菌固态发酵体系中加入纤维素酶、果胶酶和α-半乳糖苷酶,添加量分别为60、60、10 U/g,发酵体系菜籽饼Gls降解率为60.12%,TCA-SP增加率为175.72%,NDF降解率为18.25%,总变化率为48.15%。
② 酶菌协同发酵改变了菜籽饼的表面结构和蛋白质结构,使蛋白质分子质量变小。
③ 酶菌协同发酵降低了不同来源菜籽饼粕中NDF、Gls含量,增加了TCA-SP含量,改善了菜籽饼粕的营养价值。
