油莎豆种质资源低温胁迫生理响应与耐寒性评价
2023-10-15钟鹏苗丽丽王建丽刘杰王晓龙
钟鹏, 苗丽丽, 王建丽, 刘杰, 王晓龙
(1.黑龙江省农业科学院畜牧兽医分院,黑龙江 齐齐哈尔 161005; 2.黑龙江省黑土保护利用研究院,哈尔滨 150086; 3.黑龙江省农业科学院草业研究所,哈尔滨 150086)
中美贸易摩擦不断升级使我国食用油和饲料安全问题日益凸显,替代进口大豆、降低食用油和饲料对外依存度成为公众关注的热点和焦点。解决油料需求增长与耕地面积有限之间的矛盾,必需寻找单产和品质均高于大豆的食用油新油源作物开发利用。油莎豆被证明是能够部分替代进口大豆、最为现实可行的粮油饲兼用型作物[1]。油莎豆属莎草科一年生植物,耐旱、耐贫瘠、防风固沙能力强,比较适宜沙质土壤和沙化边际土地生长,不与粮争地[2-4]。在我国东北农牧交错区适时发展油莎豆产业,将是补充优质食用油供给、保护农牧交错区生态环境、补齐农业发展短板的有效途径。
低温是植物生长发育的限制因子之一,低温冷害会抑制植物生长发育、破坏细胞结构并影响正常生理代谢[5]。玉米、水稻、花生及果菜等耐低温胁迫研究较多,已经在耐低温性品种鉴定、低温胁迫生理响应及外源调节剂作用机理等方面取得一定进展[6-10]。尽管油莎豆引进我国已经70余年,但对其研究相对滞后。目前,国内外关于油莎豆的研究主要集中在栽培技术、营养成分分析、加工工艺、功能性食品开发等方面[11-14],国内关于油莎豆萌发期及苗期耐低温全面系统的评价尚未见报道。黑龙江省十年九春寒,且近几年来倒春寒现象频发,温度成为限制该地区油莎豆种植的主要因素。筛选和评价油莎豆耐低温种质资源,挖掘油莎豆耐低温基因对东北农牧交错区水土流失治理、黑土地保护利用及盐碱地改良具有重要意义。本研究在低温胁迫下对不同生态区油莎豆种质资源耐低温性进行鉴定和评价,以期明确油莎豆萌发期和苗期耐低温的主要鉴定指标,并筛选出耐低温较强的油莎豆种质资源,为后续培育耐低温品种、克隆耐低温基因及功能验证提供材料。
1 材料与方法
1.1 试验材料
试验材料为2010 年以来引进国内外不同生态区的18 份油莎豆种质资源材料,由黑龙江省农业科学院农村能源研究所能源植物资源利用研究室提供。由于国内油莎豆只有1 个登记品种中油莎1 号(ZYS01),其余均为种质材料,均用代号表示。18 份油莎豆种质材料如下:HLJ105、RY126、BY156、HLJ106、RY102、BY141、RY114、NM103、XJ101、JL154、LN181、SX100、HB122、HN162、ZYS01、XY109、GD187和QY186。
1.2 试验设计
大田试验在黑龙江省大庆市杜尔伯特蒙古族自治县新店林场试验基地(46°32′ N,124°30′ E)进行,该区属中温带大陆季风气候。2018—2020年均于5 月1 日播种,每穴1 粒,随机区组设计,3次重复,小区4.5 m×10 m,行株距45 cm×20 cm。倒春寒规定及等级标准参照文献[15-16]。
室内试验在黑龙江省农业科学院农村能源研究所进行。油莎豆种子萌发试验使用人工气候箱,光照时间为6:00—18:00,湿度80%。将种子置于直径为20 cm 的培养皿中,45 ℃温水浸泡吸胀36 h,根据大田试验倒春寒日最低温度范围,以正常温度(25 ℃)为对照,将温度设置3、5、7、9 ℃共4 个温度处理,分别培养3 d,25 ℃复温后继续培养,记录发芽时间、发芽数、芽长等数据。每个试验组每个油莎豆种质材料50粒种子,重复3次。
将油莎豆种子45 ℃温水浸泡吸胀36 h 后播种于盛珍珠岩的营养钵中,浇灌1/2 经典霍格兰(Hoagland)营养液培养萌发后,当幼苗第3 片叶完全展开后,进行低温处理。将幼苗置人工气候箱中,分别于3 、5 、7 、9 ℃ 4个温度处理2 h,以正常温度(25 ℃)为对照。每个试验组的每个种质材料8株,3次重复。
1.3 测定目标及方法
1.3.1 大田试验出苗时间和出苗率统计 2018—2020 年播种后10 d,在18 个油莎豆种质材料小区分别随机挑选3条垄每天调查出苗情况,记录出苗时间(emergence date, ED)、出苗数(emergence plant,EP),直至6月3日。统计播种穴数和出苗穴数,根据以下公式计算出苗率(emergence rate, ER)。
1.3.2 室内发芽试验 以正常温度为对照,统计18 个油莎豆种质材料在不同处理后种子发芽数、发芽时间、芽长等数据。计算发芽率(germination rate, GR)、发芽指数(germination index,GI)和种子活力指数(seed vigor index,VI)等参数[17-18]。
式中,M指发芽种子数,N指供试样品的种子数;Gt指不同发芽天数的发芽率,Dt指对应的发芽天数;S指长出部分鲜重。
1.3.3 生化特性指标测定 采用张宪政[19]的丙酮乙醇混合液法测定计算叶绿素(a+b)含量[chlorophyll content,Chl(a+b)]。相对膜透性(relative membrane permeability, RMP)测定参考李锦树等[20]的方法。丙二醛(malondialdehyde, MDA)含量测定参照Guo 等[21]的方法并进行改进。可溶性糖(soluble sugar, SS)含量和游离脯氨酸(proline, Pro)含量测定参照常博文等[7]的方法。
1.3.4 抗氧化酶活性测定 超氧化物歧化酶(superoxide dismutase, SOD)活性的测定参照Giannopolitis 等[22]的方法。过氧化物酶(peroxidase,POD)活性的测定参照Klapheck等[23]的方法。
1.4 统计分析
使用SPSS 22.0 软件进行统计分析,用Duncan’s检验(P=0.05)进行显著性分析。根据隶属函数法,综合各指标数据计算18 份种质材料隶属函数值进行抗寒性评价[24-25]。
2 结果与分析
2.1 油莎豆大田出苗时间及出苗率统计
2018—2020 年4 月28 日至6 月3 日期间试验场地连续3年均发生倒春寒现象。2018—2020年5 月11 日开始调查出苗情况,由图1 可知,HLJ105、RY126、BY156、HLJ106、RY102 和BY141出苗相对较早,每年均在5 月11 日开始出苗,于5月20 日出苗结束,出苗率均达到85.0%以上;QY186、GD187、XY109、ZYS01、HB122 和HN162出苗相对较晚,5 月14 日开始出苗,5 月22 日出苗结束,出苗率均低于65.0%。
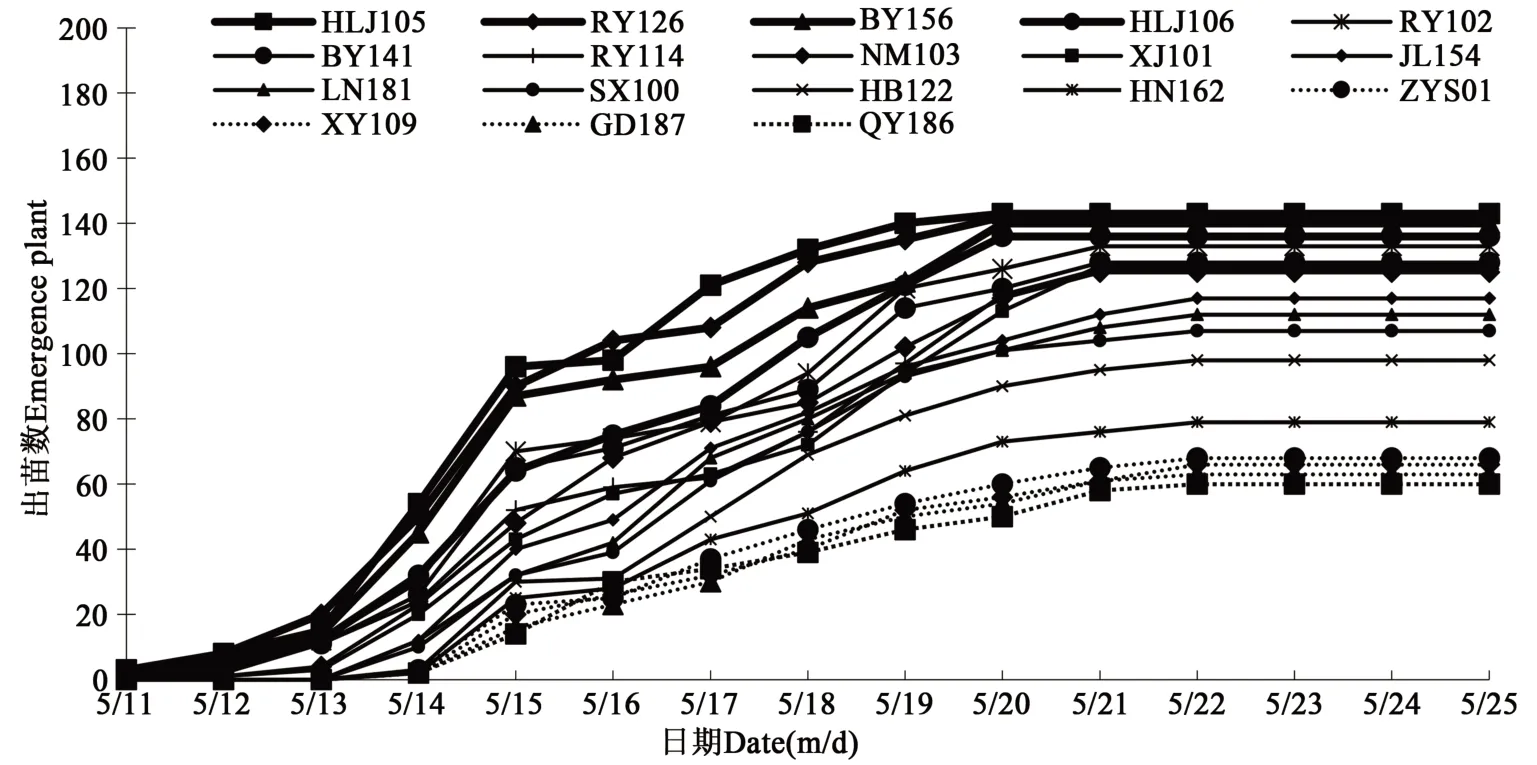
图1 2018—2020年18份种质材料出苗情况Fig. 1 Emergence of 18 germplasm materials in 2018-2020
2018—2020 年均在6 月3 日统计出苗率,由表1 可知,3 年出苗率较高的种质材料为HLJ105、RY126、BY156、HLJ106、RY102 和BY141,平均出苗率为95.21%、94.63%、93.32%、90.50%、88.96%和85.93%;而出苗率较低的种质材料为QY186、GD187、XY109、ZYS01、HN162 和HB122,平均出苗率为40.13%、41.88%、43.84%、45.34%、52.98%和65.56%。从3年气温变化、出苗情况、出苗率综合比较,HLJ105、RY126、BY156 和HLJ106 耐低温性较强,QY186、GD187、XY109 和ZYS01 耐低温性较弱。
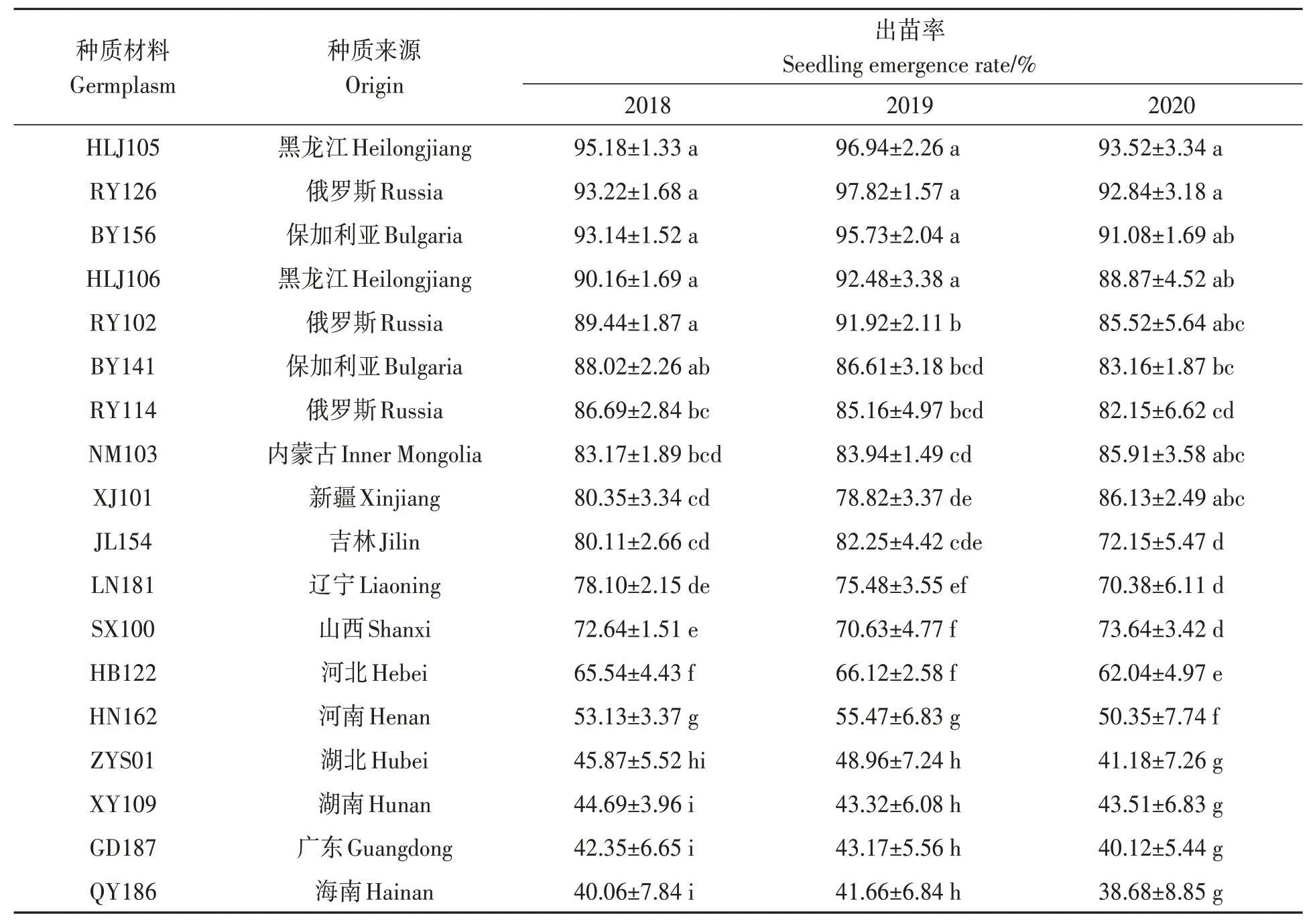
表1 油莎豆种质材料出苗率Table 1 Seedling emergence rates of all Cyperus esculentus germplasms
2.2 室内低温胁迫对油莎豆种子发芽指数和活力指数的影响
由表2 可知,随着处理温度的降低,不同油莎豆种质材料发芽指数均逐渐下降。3 ℃处理时,HLJ105、RY126、BY156 和HLJ106 种子发芽指数相对较高,与其他14 份材料差异均达显著水平(P<0.05);QY186、GD187、XY109和ZYS01种子发芽指数相对较低,除与HN162差异不显著外,与其余13 份材料差异均达显著水平(P<0.05)。5 ℃处理时,HLJ105、RY126、BY156 和HLJ106 种子发芽指数显著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 种子发芽指数相对较低,除与HN162、SX100 和JL154 差异不显著外,显著低于其余11 份材料。7 ℃处理时,HLJ105、RY126、BY156、HLJ106、RY102、BY141、NM103、JL154 和HN162共9份材料之间差异不显著,但均显著高于除LN181外的其他8份材料。9 ℃处理时,与对照组发芽指数基本一致,可达到对照组的97.6%。
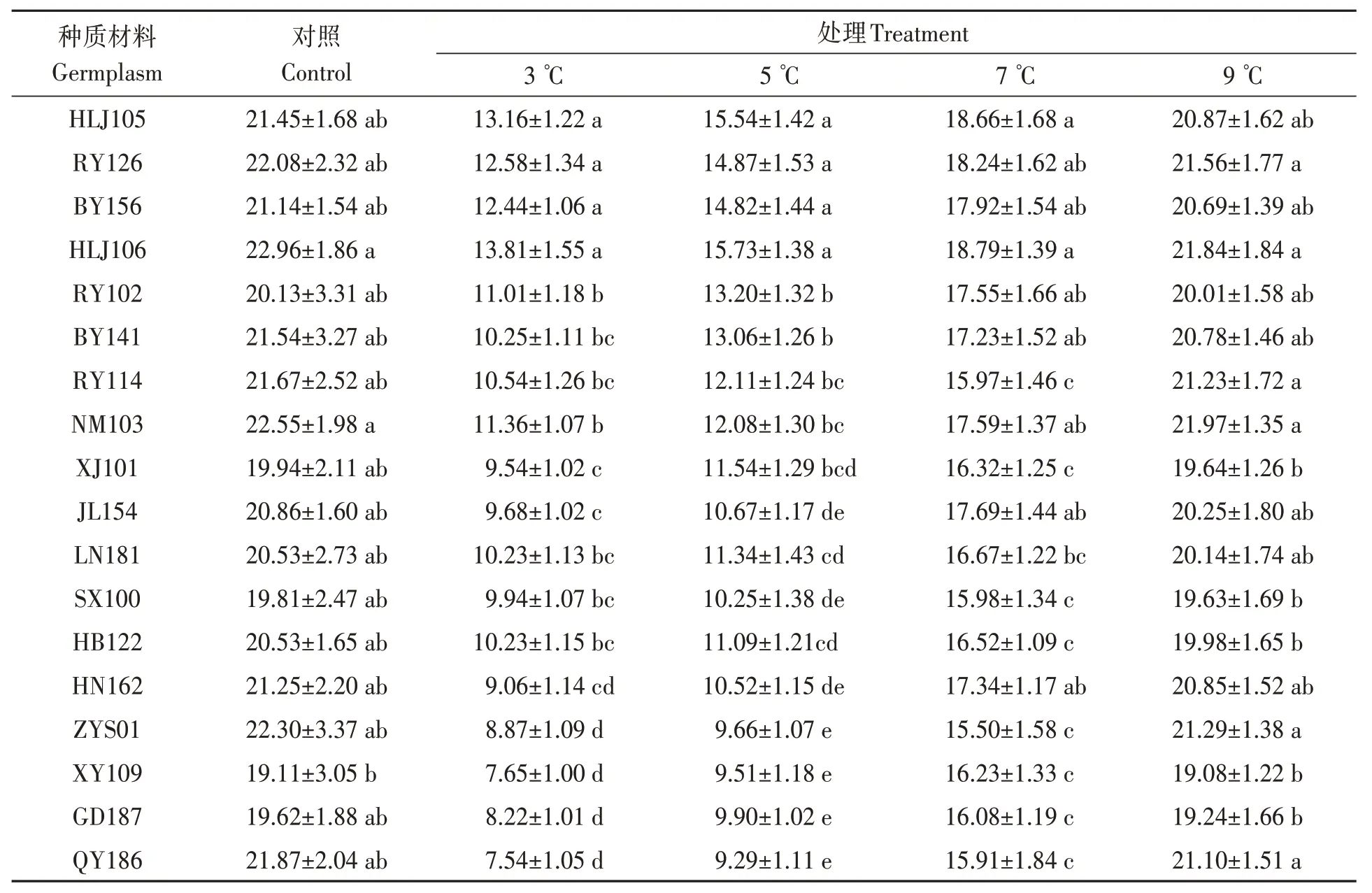
表2 低温胁迫对油莎豆种子发芽指数的影响Table 2 Effects of low temperature on germination index of Cyperus esculentus germplasms
由表3可知,随着处理温度的降低,除NM103外其他油莎豆种子活力指数逐渐下降。3 ℃处理时,HLJ105、RY126、BY156 和HLJ106 种子活力指数相对较高,与其他14 份材料差异均达显著水平(P<0.05);QY186、GD187、XY109 和ZYS01 种子活力指数显著低于其他14 份材料(P<0.05)。5 ℃处理时,HLJ105、RY126、BY156、HLJ106、RY102和BY141 种子活力指数相对较高,与其他12 份材料差异均达显著水平;QY186、GD187、XY109 和ZYS01 种子活力指数相对较低,与其他14 份材料差异均达显著水平。7 ℃处理时,HLJ105、RY126、BY156、HLJ106、BY141、ZYS01、RY102 和LN181 这8 份材料之间差异不显著,均与GD187之间差异显著。9 ℃处理时,处理组较对照组相比活力指数下降幅度不大,可达到对照组的96.8%。
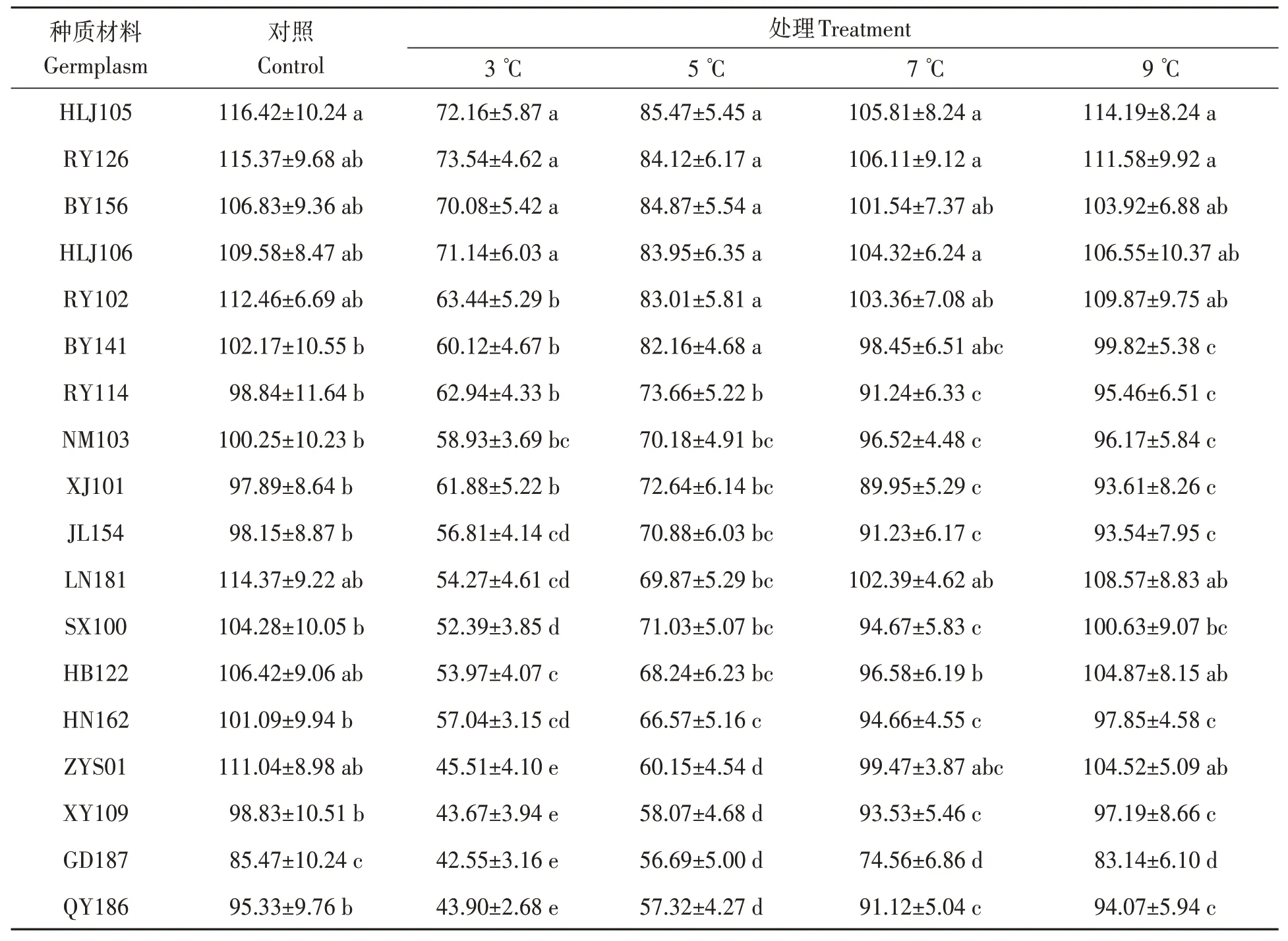
表3 低温胁迫对油莎豆种子活力指数的影响Table 3 Effects of low temperature on seed vigor of Cyperus esculentus germplasms
2.3 低温胁迫对油莎豆幼苗叶绿素含量的影响
随着温度的降低,油莎豆幼苗叶片叶绿素(a+b)含量逐渐降低(图2)。3 ℃处理时,HLJ105、RY126、BY156 和HLJ106 叶绿素(a+b)含量较高;QY186、GD187、XY109 和ZYS01 叶绿素(a+b)含量较低。5 ℃处理时,HLJ105、RY126、BY156 和HLJ106叶绿素(a+b)含量相对较高。7 和9 ℃处理时所有材料之间叶绿素(a+b)含量差别不明显。
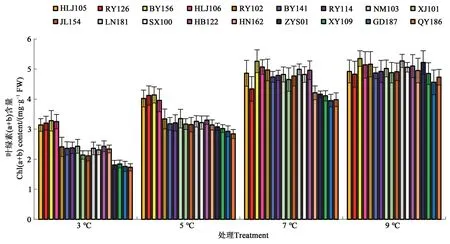
图2 低温胁迫对油莎豆幼苗叶片叶绿素含量的影响Fig. 2 Effects of low-temperature on the Chl(a+b) contents of Cyperus esculentus germplasms leaves
2.4 低温胁迫对油莎豆幼苗叶片质膜损伤程度的影响
随着处理温度的降低,油莎豆叶片相对膜透性逐渐增大(表4)。3 和5 ℃处理时,HLJ105、RY126、BY156 和HLJ106 与对照相比相对膜透性均增加,但显著低于其他14 份种质材料。3 和5 ℃处理,QY186、GD187、XY109 和ZYS01 与对照相比相对膜透性均增加,但显著高于其他14 份材料。7 ℃处理下,QY186、GD187、XY109、HN162、JL154 和ZYS01 相对膜透性较大。9 ℃处理时,QY186 和GD187 相对膜透性与XJ101、BY141、RY102、BY156 和HLJ105 差异显著,与其他材料之间差异不明显。
3和5 ℃处理,丙二醛含量变化与相对膜透性基本相似,HLJ105、RY126、BY156 和HLJ106 丙二醛含量较对照均增加,但丙二醛含量相对较少,显著低于其他14 份种质材料;QY186、GD187、XY109 和ZYS01 丙二醛含量较对照均增加,但丙二醛含量相对较大,显著高于其他14 份材料。7 ℃处理时,HLJ105、RY126、BY156、RY114、SX100 和HLJ106 丙二醛含量相对较低,与除HN162 外的其他11 份材料差异显著。9 ℃处理时,HLJ105、RY126、BY156、RY102、RY114、LN181、HLJ106 的丙二醛含量与QY186、GD187、XY109、ZYS01 之间差异显著,与其他材料之间差异不显著。
2.5 低温胁迫对油莎豆幼苗叶片可溶性物质含量的影响
除RY114 外,油莎豆叶片游离脯氨酸含量随着温度的降低急剧升高(表5)。当处理温度为3 ℃时,HLJ105、RY126、BY156 和HLJ106 游离脯氨酸含量显著高于其他14 份材料;QY186、GD187、XY109和ZYS01显著低于其他14份材料。处理温度为5 ℃时,HLJ105、RY126、BY156、RY102、HLJ106 和BY141 游离脯氨酸含量显著高于其他12 份材料。处理温度为7 ℃时,HLJ105、RY126、BY156、RY102、BY141、RY114和HLJ106游离脯氨酸含量显著高于其他11 份材料;处理温度为9 ℃时,HLJ105、BY156 与HB122、QY186、GD187、XY109之间游离脯氨酸含量差异显著。
除HN162 外,油莎豆叶片可溶性糖含量随着温度的降低而增加(表5)。处理温度为3 ℃时,HLJ105、RY126、BY156 和HLJ106 可溶性糖含量显著高于其他14 份材料;QY186、GD187、XY109和ZYS01 显著低于其他14 份材料。在处理温度5 ℃时,HLJ105、RY126、BY156、HLJ106、RY102、BY141 和RY114 可溶性糖含量显著高于其他11份材料。在处理温度7 ℃时,QY186、GD187、XY109、HN162、XJ101 和ZYS01 可溶性糖含量显著低于其他12份材料。
2.6 低温胁迫对油莎豆幼苗叶片抗氧化酶活性的影响
由表6 可知,SOD 活性随着温度的降低而升高(HN162 除外)。当温度降至3 ℃时,HLJ105、RY126、BY156 和HLJ106 叶片中SOD 活性显著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 显著低于其他14 份材料。当处理温度为5 ℃时,HLJ105、RY126、BY156、HLJ106 和RY102材料叶片中SOD 活性显著高于其他13 份材料;QY186、GD187、XY109 和ZYS01 显著低于其他14份材料。7 和9 ℃时,HLJ105、RY126、BY156、HLJ106、RY102、BY141 和RY114 材料SOD 活性显著高于其他11 份材料,其余材料之间差异不显著。
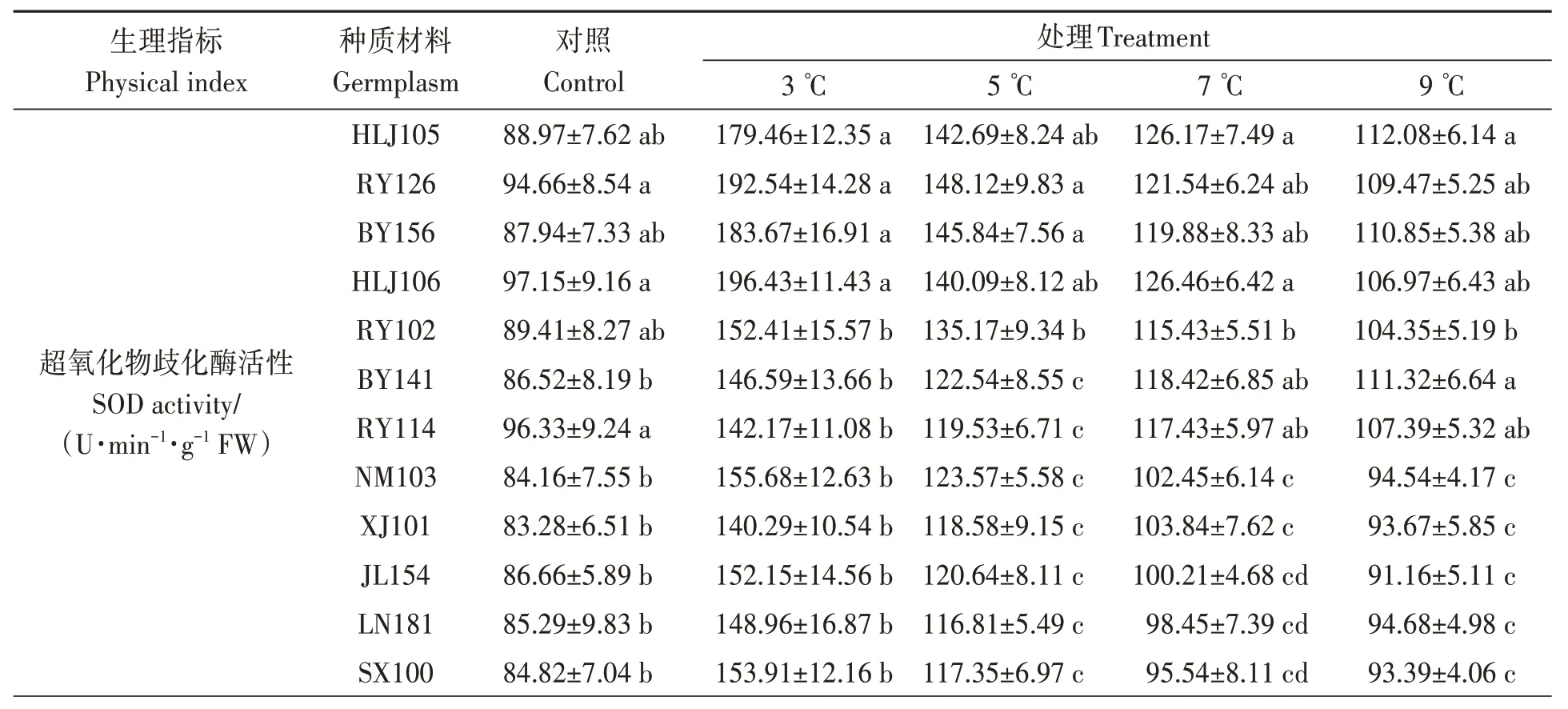
表6 低温胁迫对油莎豆幼苗叶片抗氧化酶活性的影响Table 6 Effects of low-temperature on anti-oxidase activity of Cyperus esculentus germplasms leaves
POD 活性变化趋势与SOD 大致相同。当温度降至3 ℃时,HLJ105、RY126、BY156 和HLJ106叶片中POD 活性显著高于其他14 份材料;QY186、GD187、XY109 和ZYS01 显著低于其他14份材料。当温度为5 ℃时,HLJ105、RY126、BY156、HLJ106、RY102、BY141和RY114材料叶片中POD 活性显著高于其他11 份材料。当温度为7和9 ℃时,18份材料之间POD活性差异不显著。
2.7 18份油莎豆种质材料抗寒性综合评价
18份油莎豆种质材料抗寒性综合评价情况见表7。参照刘慧民等[24]、张文娥等[25]的方法,将各材料综合抗寒能力平均隶属度分为5 级。抗寒性综合评价值(cold-resistance integrated assessment value, IAV)≥0.70为强抗寒性,定为Ⅰ级;抗寒性综合评价值在0. 60~0. 69为抗寒性,定为Ⅱ级;抗寒性综合评价值在0.40~0. 59为中抗寒性,定为Ⅲ级;抗寒性综合评价值在0. 30~0. 39为弱抗寒性,定为Ⅳ级;综合评价值< 0.30 为不抗寒性,定为Ⅴ级。经大田、室内种子发芽和幼苗抗寒性试验综合评价HLJ105、RY126、BY156和HLJ106抗寒性综合评价值分别为0.708 6、0.712 4、0.715 3 和0.705 4,为强抗寒性种质材料;RY102和BY141抗寒性综合评价值分别为0.616 0 和0.601 5,为抗寒性材料;RY114、NM103、XJ101、JL154、LN181 和SX100 抗寒性综合评价值在0.40~0. 59,为中抗寒性材料;HB122 和HN162 抗寒性综合评价值在0. 30~0. 39之间,为弱抗寒性材料;QY186、GD187、XY109 和ZYS01抗寒性综合评价值均小于0.30,为不抗寒性材料。
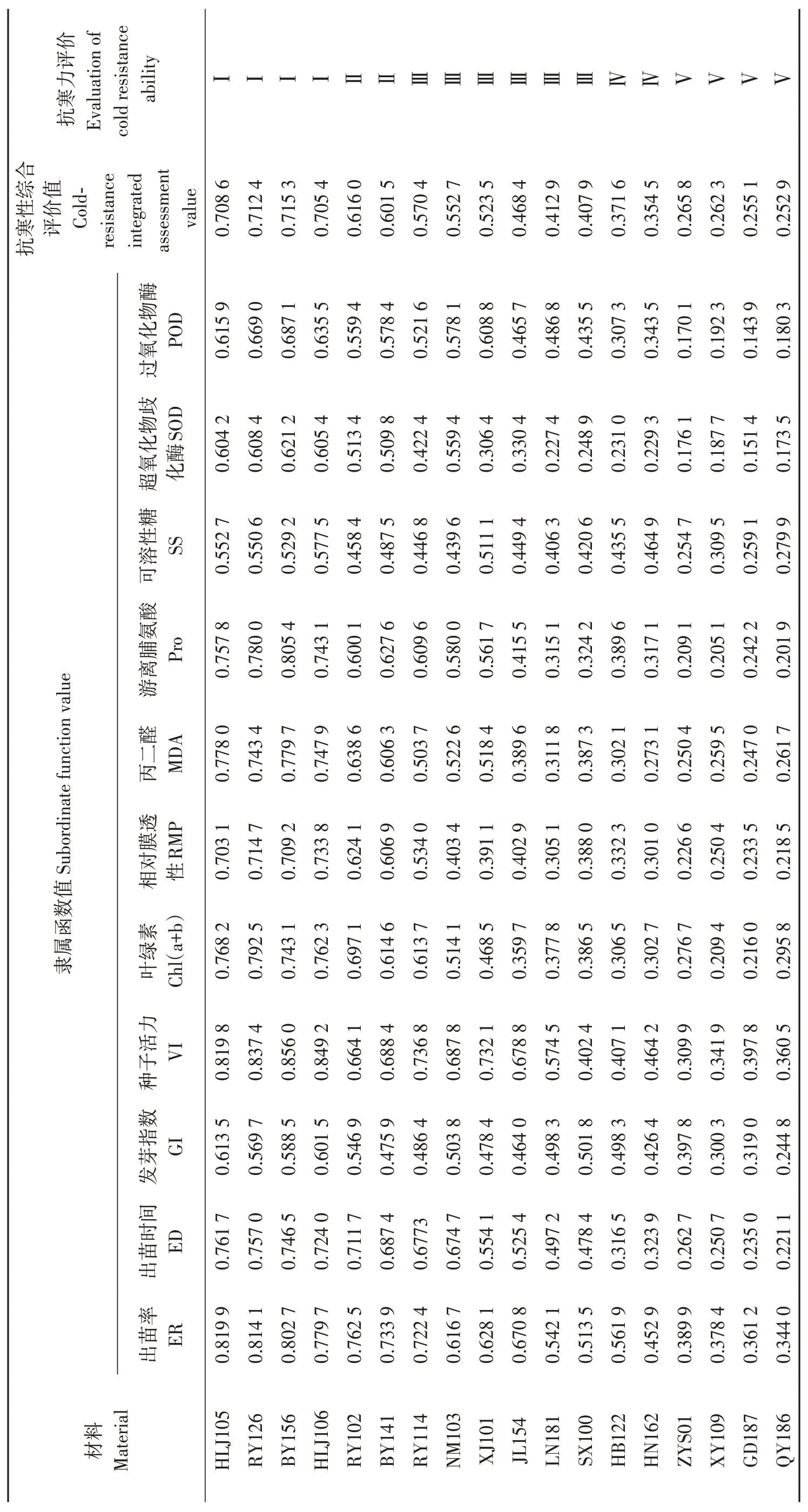
表7 油莎豆各指标隶属函数值、权重及抗寒性综合评价值Table 7 Subordinate function values,weight function and cold-resistance integrated evaluation values of all indexes in Cyperus esculentus germplasms
2.8 各测定指标抗寒性相关分析
低温胁迫下出苗率(ER)、发芽指数(GI)和超氧化物歧化酶(SOD)活性与抗寒性综合平均值(IAV)相关系数分别为0.978 2、0.953 6和0.970 1,呈显著正相关(P<0.05),出苗时间(ED)和丙二醛(MDA)含量与抗寒性综合平均值(IAV)相关系数分别为-0.912 8 和-0.923 3,呈显著负相关(P<0.05)(表8)。种子活力(VI)、叶绿素[Chl(a+b)]、游离脯氨酸(Pro)、可溶性糖(SS)含量和过氧化物酶(POD)活性与抗寒性综合评价值(IAV)呈正相关,但相关性均不显著;相对膜透性(RMP)与抗寒性综合评价值(IAV)呈负相关,相关性不显著。出苗率(ER)与发芽指数(GI)、种子活力(VI)呈正相关,相关性达到极显著水平(P<0.01)。
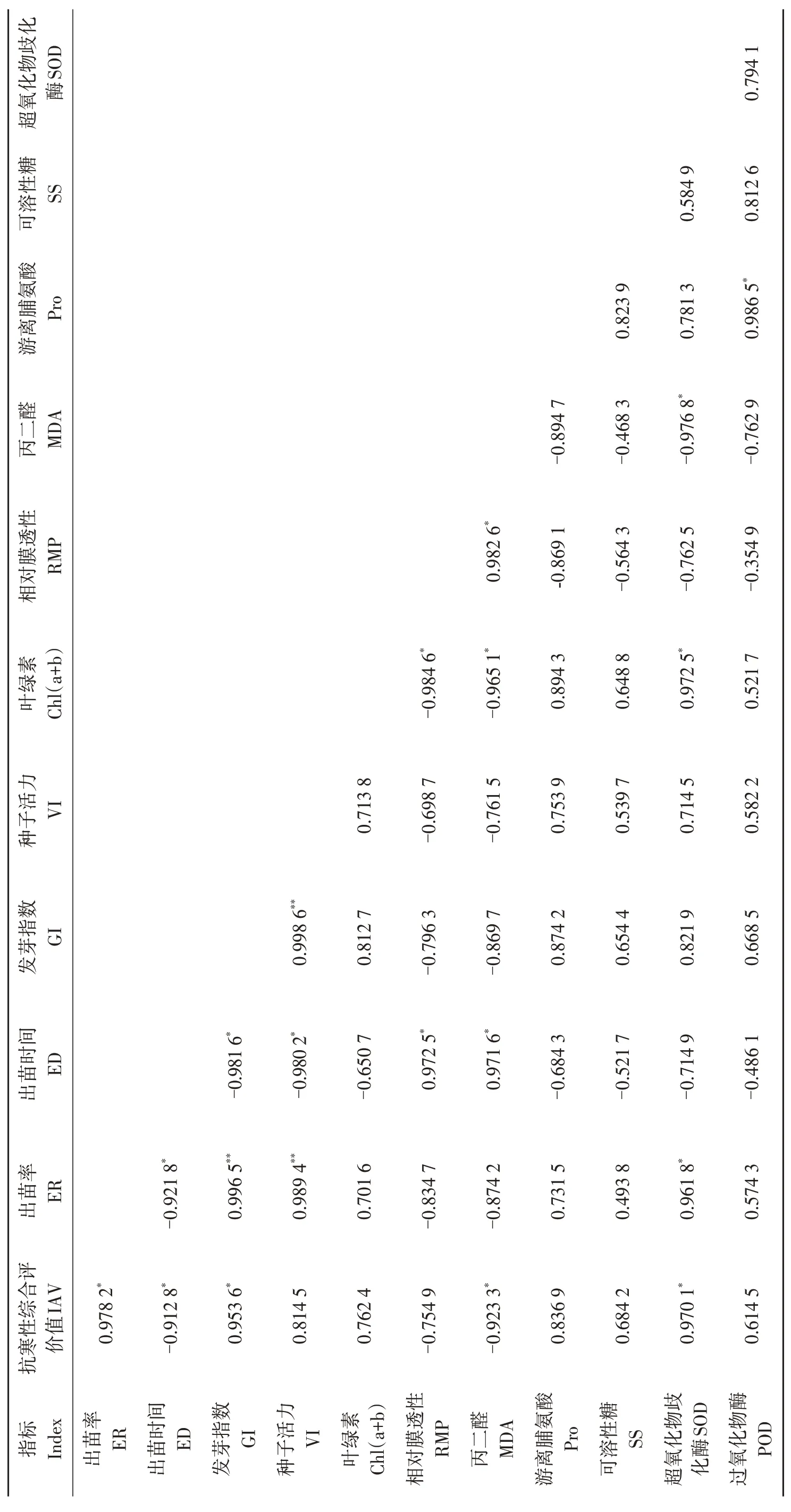
表8 供试油莎豆材料各测定指标及苗期抗寒相关性分析Table 8 Correlation analysis of subordinate function values of cold resistance index for tested varieties in Cyperus esculentus germplasms
3 讨论
低温不仅影响植物生长发育,而且限制其地理分布,当受到低温胁迫时植物可通过调节自身生理代谢和基因表达来增强低温耐受性[26]。近年来,倒春寒现象在我国发生频率逐渐上升,尤其东北地区更为严重,轻者补苗,重者毁种,给农业生产带来不可估量的损失[27]。种子在吸胀萌动和初始发育阶段对低温最为敏感[28]。本团队前期通过低温胁迫筛选出耐低温和不耐低温的花生品种,明确了耐低温品种发芽率、发芽指数、种子活力和芽长均高于不耐低温品种[7,29]。本试验以油莎豆为研究对象,大田试验中连续3 年经历倒春寒,HLJ105、RY126、BY156 和HLJ106 出苗时间较其他材料早3 d,出苗率均大于90.0%,并且与其他材料相比差异显著,说明其耐低温能力较强,低温对其种子萌发出苗影响较小;而QY186、GD187、XY109 和ZYS01 出苗时间较其他材料晚,出苗率均小于50.0%,并且与其他材料相比差异显著,说明其耐低温能力较弱,倒春寒对其种子萌发出苗影响较大。室内试验发芽率、种子活力等指数与大田结果一致。
细胞膜对维持植物细胞内环境稳定性起着重要作用。低温胁迫时,丙二醛含量激增,导致质膜透性增大,胞内电解质外渗,引起相对电导率升高[30]。本研究显示,随着温度的降低,18份种质材料的质膜透性和丙二醛含量均显著升高,说明温度越低油莎豆叶片细胞膜受冷害程度越严重;当温度降至3和5 ℃时,材料HLJ105、RY126、BY156和HLJ106 的相对膜透性和丙二醛含量上升幅度低于其他材料,而QY186、GD187、XY109 和ZYS01 上升幅度高于其他材料,说明耐低温性强的油莎豆材料细胞膜受低温伤害较小,抗寒性优于其他材料。质膜透性和丙二醛含量呈显著正相关,说明油莎豆叶片质膜损伤大部分是由丙二醛含量积累所导致,与之前花生抗寒性评价研究结果不一致[29],有待进一步深入研究。
