黄土丘陵区撂荒年限和坡向对灌丛生物量及丰富度的影响
2023-10-08野起瑞李江文张晓曦裴婧宏
刘 冲, 野起瑞, 李江文*, 张晓曦, 邓 健, 裴婧宏, 樊 慧
(1.延安大学生命科学学院, 陕西 延安 716000; 2.陕西省红枣重点实验室(延安大学), 陕西 延安 716000)
黄土高原水土流失严重,由于长时间过度开垦和不合理的耕作,加之该地区降水少而集中,且黄土本身孔隙多且大,土质松散,具有很强的湿陷性,对流水的抵抗力弱,易受侵蚀,导致黄土丘陵区水土流失最为严重。自20世纪80年代开始,我国陆续在黄土高原丘陵沟壑区开展退耕还林(草)工程[1]。撂荒耕地作为其中主要的组成部分,在近几十年的撂荒过程中逐渐形成了区域沟壑纵横、植物群落极不稳定的草地生态系统,其中植物群落物种组成及生物量的变化对于解释不同撂荒年限草地生态系统过程至关重要。
地上生物量作为草地生态系统能量获取的主要表现形式,在草地生态系统的功能和结构形成过程中具有十分重要的作用[2]。地上生物量既受到光照、水分、海拔、地形、土壤等自然因素的影响,也会因群落物种组成及结构的不同而变化。早期的研究结果显示,随着撂荒年限的增加,撂荒草地植物群落地上生物量呈现先减少后增加的变化趋势[3],但也有观点指出,植物地上生物量总体表现为随撂荒年限的增加而增加,也会出现波动[4]。同时,近些年有很多学者从土壤学角度分析了撂荒草地群落生物量与土壤养分之间的交互效应[5],为植物群落物种组成及变化提供了诸多解释依据。物种丰富度可表征植物群落结构和功能的复杂性[6],体现了植物群落在组成、结构、稳定性及生境上的差异性[7-8]。目前对于植物群落物种丰富度的研究多集中于环境梯度和演替过程中的变化[9]。而坡向作为黄土丘陵区一项重要的地形因素,影响着土壤、温度、水分及光照的变化[10],同一区域不同坡向的微气候和土壤养分存在很大差异,从而导致草地植物群落物种组成、种群空间格局及生物量改变[11-12]。灌丛群落作为植物群落重要的组成部分,其增加可显著影响草地植物群落物种多样性及生产力[13]。因此,在撂荒年限和坡向因素的共同影响下,撂荒草地植物灌丛地上生物量及丰富度的变化规律尚需进一步讨论。基于撂荒年限和坡向变化的交互效应,研究草地灌丛植物群落生物量及物种丰富度的变化,对于黄土丘陵区植物多样性保护具有重要的指导意义。
黄土丘陵区草地生态环境脆弱、敏感,植物群落生产力低下、稳定性差,容易受到人类活动及自然环境的干扰。撂荒草地作为黄土丘陵区的典型代表,灌木和半灌木在草地生态系统中占据着重要的地位。本研究以黄土丘陵区不同撂荒年限草地作为研究对象,在植物群落调查的基础上,通过调差不同坡向草地群落、灌木和半灌木地上生物量,以及物种丰富度变化。采用巢式样方法确定撂荒草地最佳取样面积,探讨撂荒年限和坡向对植物群落物种丰富度及地上生物量的影响,为黄土丘陵区撂荒草地植物多样性保护和可持续发展提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市安塞区纸坊沟流域,采样点坐标如表1所示,平均海拔约1 239 m。该区域属温带半干旱气候,年平均气温约8.9℃,年平均降水量491.23 mm(数据来源于试验区域内设置的JD-QC5小型气象站,2022年监测数据),全年70%以上降水主要集中在6—9月。撂荒年限的界定主要依据为退耕荒草工程的实施年限,撂荒前为耕地,主要种植作物为糜子(Panicummiliaceum)和谷子(Setariaitalica),退耕前施肥水平较低。该地区属于黄土高原丘陵沟壑区第二副区,区域内沟壑纵横,地形起伏大,区域面积约为8.27 km2。土壤主要以黄绵土为主。该区域撂荒草地主要植物为狗尾草(Setariaviridis)、阿尔泰狗娃花(Asteraltaicus)、小花鬼针草(Bidensparviflora)、野菊(Chrysanthemumindicum)、香青兰(Dracocephalummoldavica)、早熟禾(Poaannua)、达乌里胡枝子(Lespedezadavurica)等。主要灌木和半灌木包括:铁杆蒿(Artemisiasacrorum)、茭蒿(Artemisiagiraldii)、柠条锦鸡儿(Caraganakorshinskii)、山莓(Rubuscorchorifolius)、黄刺玫(Rosaxanthina)等。
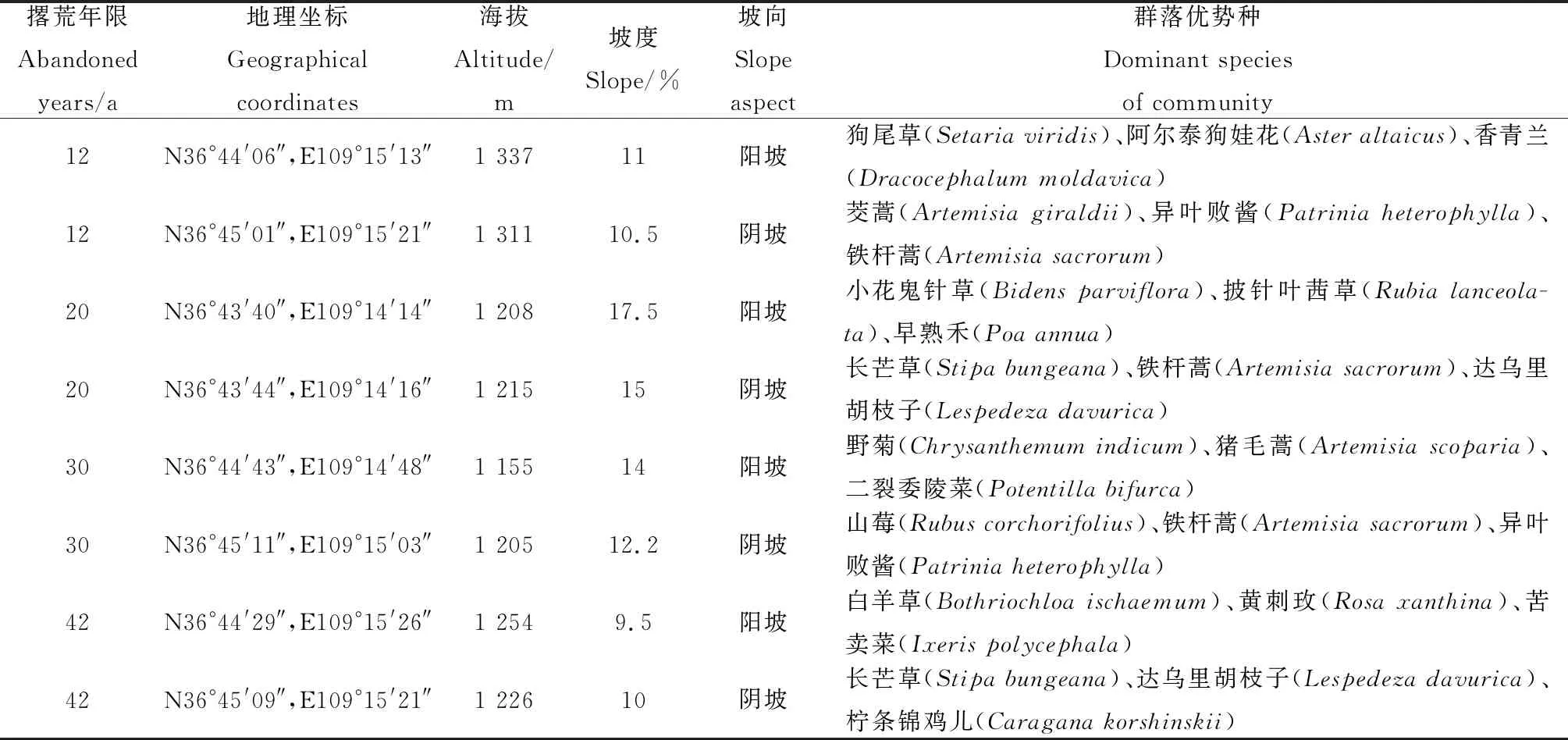
表1 调查样地基本概况
1.2 研究方法
于纸坊沟流域内选取不同撂荒年限(12 a,20 a,30 a和42 a)的阳坡和阴坡草地,要求不同撂荒年限阳坡和阴坡草地立地条件基本一致,且每个撂荒年限的阳坡和阴坡分别选取3个重复样地,共计调查样地24个(其中阳坡12个、阴坡12个),每个样地设置5个1 m×1 m的固定样方,主要用于调查草地植物群落物种组成和各物种地上生物量。同时,在每个调查样地内设置3个巢式样方(如图1),每个巢式样方分别包含0.25,0.5,1,2,4,8,16,32,64,128,256 m2,共计11级样方,在各级样方内统计植物群落物种数。
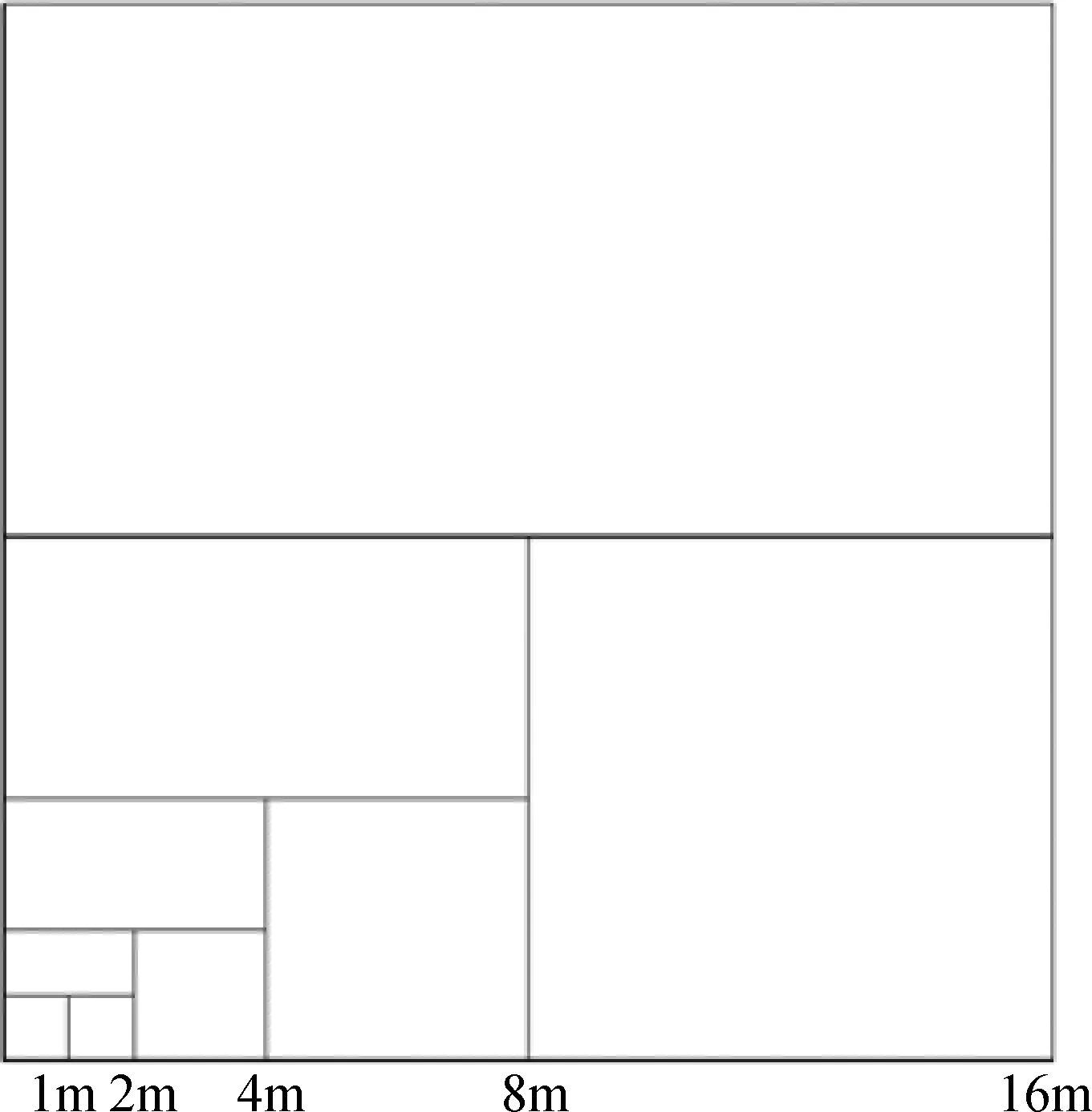
图1 巢式样方调查图
1.3 数据采集与分析
2022年8月,分种采集固定样方内的植物,登记样方内物种组成后,将样方内采集物种分种装于不同信封带内,做好标记后带回实验室。将采集好的植物样品置于65℃烘箱内,烘干24 h至恒重,分种称取重量后,计算固定样方单位面积地上生物量和灌木和半灌木的地上生物量。同时,在选取样地内设置巢式样方,统计巢式样方不同面积内的植物物种数量,用于核算种-面积关系,物种丰富度取单位面积内(1 m2)的物种类别数目。
利用SPSS 23.0统计软件对植物群落地上生物量及物种丰富度做差异性分析(包括(One-way ANOVA和Double factor variance analysis,P<0.01 &P<0.05),并分析阳坡和阴坡撂荒草地群落地上生物量、植物群落物种丰富度、撂荒年限之间的pearson相关系数(P<0.01 &P<0.05),作图软件使用SigmaPlot 12.5。
2 结果与分析
2.1 不同撂荒年限草地灌丛地上生物量
不同撂荒年限草地地上生物量差异性分析如图2所示,30 a和42 a撂荒草地地上生物量显著高于12 a和20 a(P<0.05),30 a撂荒草地地上生物量最高,阳坡和阴坡分别达到133.20 g·m-2和100.18 g·m-2,且12 a至30 a阳坡草地地上生物量随撂荒年限的增长而增加,且30 a显著高于20 a,20 a显著高于12 a(P<0.05,图2左)。20 a,30 a和42 a撂荒草地阳坡地上生物量显著高于阴坡草地(P<0.05)。整体而言,撂荒草地地上生物量随撂荒年限的增长而增加,但峰值出现在30 a。不同撂荒年限草地灌木和半灌木地上生物量差异性分析显示,阳坡30 a撂荒草地灌木和半灌木地上生物量最高,达到50.15 g·m-2,且显著高于其他撂荒年限草地灌木和半灌木地上生物量(P<0.05)。而阴坡42 a撂荒草地灌木和半灌木地上生物量最高,达到42.97 g·m-2,且30 a和42 a撂荒草地灌木和半灌木地上生物量显著高于12 a和20 a(P<0.05)。12 a,20 a和42 a撂荒草地阳坡灌木和半灌木地上生物量显著低于阴坡撂荒草地(P<0.05)。30 a撂荒草地阳坡和阴坡灌木和半灌木地上生物量百分比接近36%,除30 a外,自12 a至42 a撂荒草地阳坡和阴坡灌木和半灌木地上生物量百分比逐年增加,且阴坡草地42 a百分比最高,达到51.68%。总体而言,阴坡撂荒草地灌木和半灌木地上生物量随撂荒年限的增长而增加,但阳坡撂荒草地峰值出现在30 a。阴坡撂荒草地灌木和半灌木地上生物量百分比整体上高于阳坡草地。双因素方差分析结果显示,撂荒年限、坡向以及二者的交互效应对群落地上生物量、灌木和半灌木地上生物量均产生显著影响(P<0.01,P<0.05)。
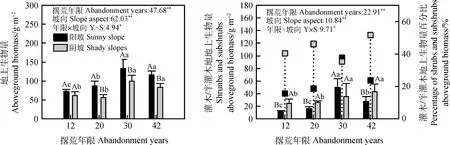
图2 不同撂荒年限植物群落地上生物量差异性分析
2.2 不同撂荒年限草地灌丛群落物种丰富度
分析不同撂荒年限草地植物群落物种丰富度变化,结果如图3所示,阳坡撂荒草地30 a植物群落物种丰富度最高,平均可达到10.5种·m-2,且30 a和42 a植物群落物种丰富度显著高于12 a和20 a(P<0.05)。阴坡撂荒草地植物群落物种丰富度随撂荒年限的增长而增加,42 a撂荒草地植物群落物种丰富度最高,平均约为8种·m-2,且12 a撂荒草地植物群落物种丰富度显著低于20 a,30 a和42 a(P<0.05)。30 a和42 a阳坡撂荒草地植物群落物种丰富度显著高于阴坡(P<0.05)。不同撂荒年限阳坡和阴坡撂荒草地灌木和半灌木物种数目均无显著性变化,但随着撂荒年限的增加,植物群落中灌木和半灌木物种数目呈逐渐增加的趋势,并且阴坡撂荒草地灌木和半灌木物种数目百分比均高于阳坡草地,平均占比30.47%;阳坡撂荒草地灌木和半灌木物种数目百分比随撂荒年限的增加而降低,12 a占比最高达28.57%。双因素方差分析结果显示,撂荒年限、坡向对群落物种丰富度产生显著影响(P<0.01),但二者间的交互效应对物种丰富度无显著影响。撂荒年限、坡向及二者的交互效应对灌木和半灌木物种数目无显著影响。
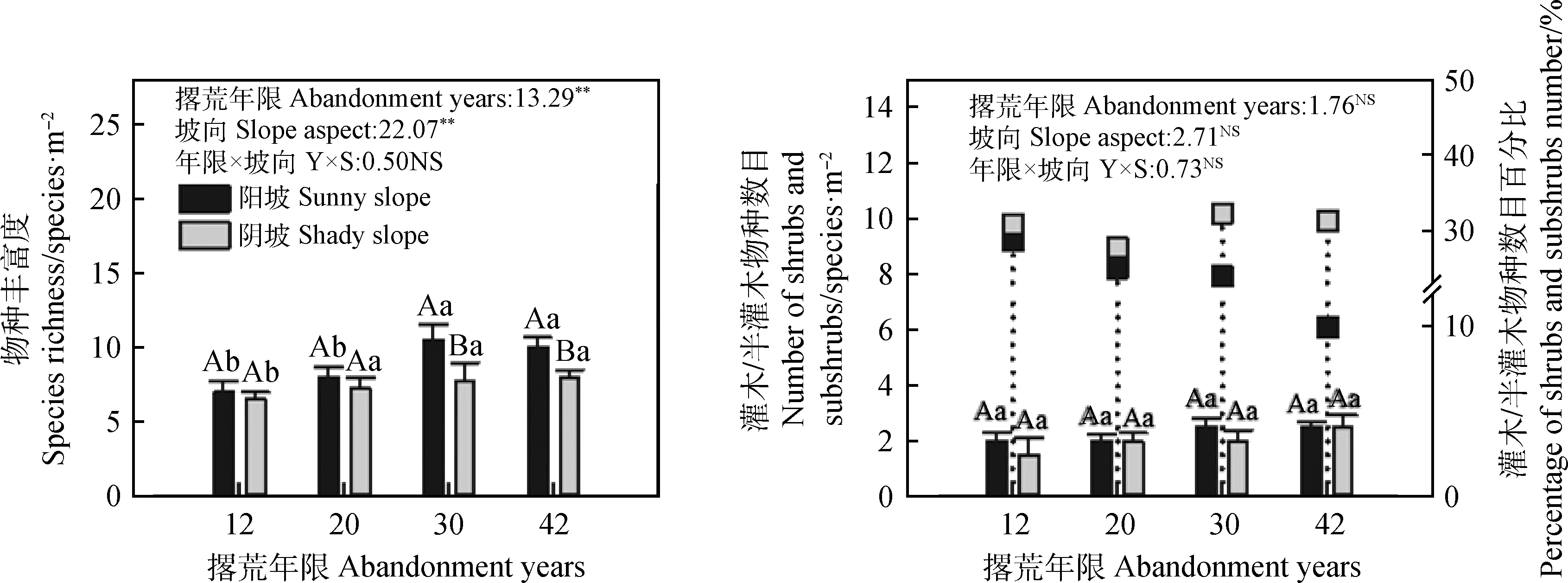
图3 不同撂荒年限植物群落物种丰富度及灌丛物种数目差异性分析
不同撂荒年限草地阳坡和阴坡植物群落物种数目均随取样面积的增大而增加,取样面积从0.25 m2到32 m2尺度上植物群落物种数据快速增加,32 m2以后物种数目增加的速度趋于平缓(图4)。不同撂荒年限之间物种数目进入平衡态的点位主要集中在128 m2上,但平衡态上的物种数不尽相同。12 a平衡态阳坡物种数为19,但阴坡仅为18;20 a平衡态阳坡物种数最大为22,阴坡物种数自128 m2后一致保持在21,30 a和42 a阳坡和阴坡平衡态物种数保持一致,分别为21和23。整体而言,随着撂荒年限的增加,进入平衡态的植物群落物种数目逐渐增多。但是,不同撂荒年限草地阳坡和阴坡灌木和半灌木物种数目进入平衡态的取样面积不尽相同。12 a和20 a阳坡和阴坡灌木和半灌木进入平衡态物种数目一致,但12 a阳坡进入平衡态3的取样面积为2 m2,而阴坡则需要达到16 m2;20 a阳坡灌木和半灌木物种数目进入平衡态4的取样面积为2 m2,而阴坡则需要达到16 m2。30 a阳坡灌木和半灌木物种数目进入平衡态5的取样面积为8 m2,而阴坡进入平衡态6则需要达到64 m2;42 a阳坡和阴坡灌木和半灌木物种数目进入平衡态的取样面积均为32 m2,但阳坡灌木和半灌木进入平衡态的物种数目为4,而阴坡为5。整体而言,群落中灌木和半灌木物种数目随取样面积的增大而增加,随着撂荒年限的增加,群落物种数进入平衡态的数目逐渐增加,但灌木和半灌木物种数目呈先增加后降低的趋势。
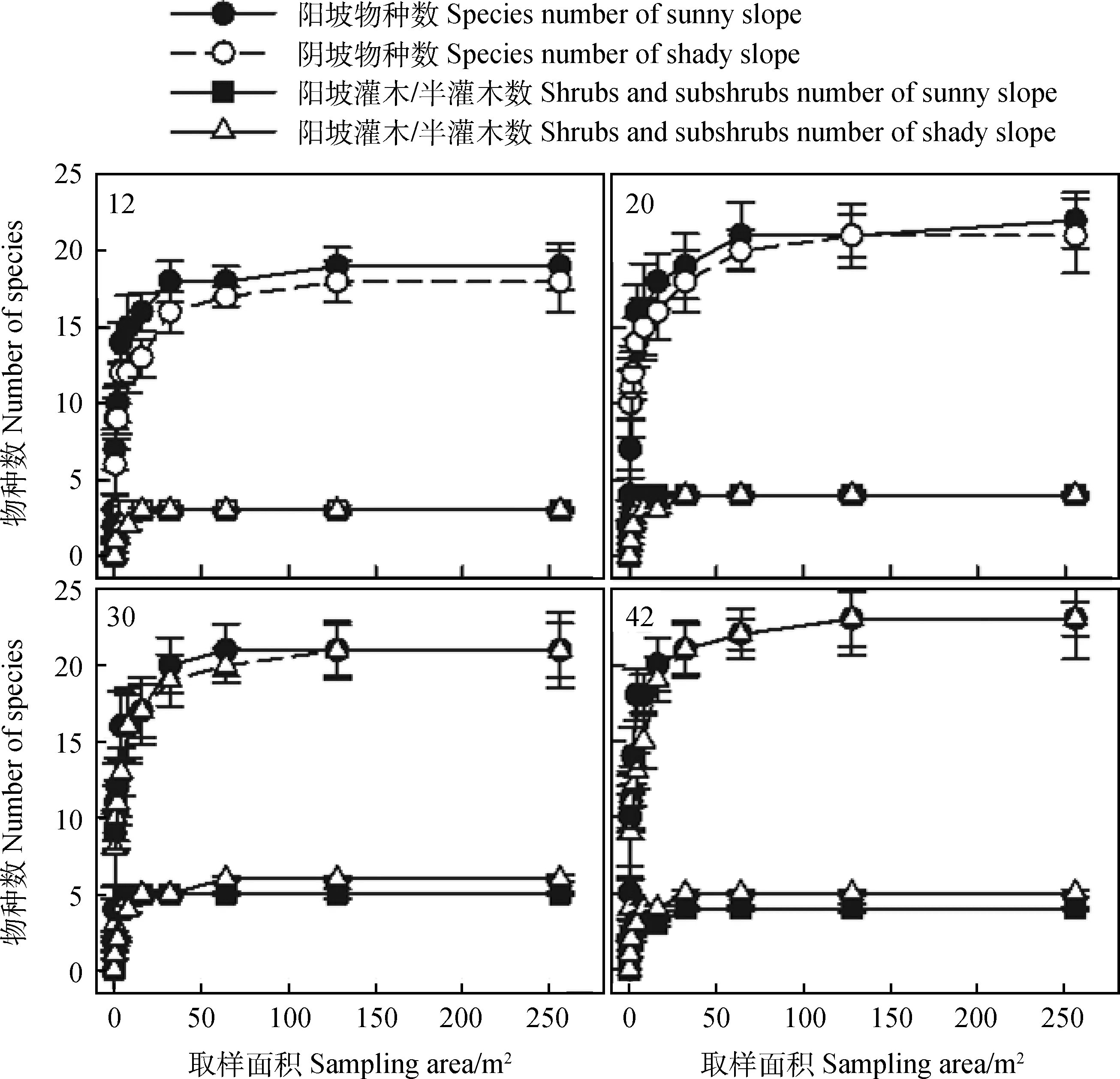
图4 不同撂荒年限植物群落种-面积关系
2.3 撂荒草地地上生物量与物种数目相关性分析
撂荒草地地上生物量与物种丰富度相关性分析结果如表2所示,地上生物量与灌丛地上生物量之间具有极显著正相关关系(P<0.01),阳坡地上生物量、群落物种丰富度与撂荒年限之间极显著相关(相关系数为0.69和0.74,P<0.01),而灌丛地上生物量与撂荒年限之间的相关系数为0.53(P<0.05);阴坡地上生物量和灌丛地上生物量与撂荒年限之间存在极显著相关性(相关系数为0.66和0.73,P<0.01),而群落物种丰富度与撂荒年限之间的相关系数仅为0.49(P<0.05)。阳坡植物群落物种丰富度与地上生物量和灌丛地上生物量极显著正相关(P<0.01),但阴坡植物群落物种丰富度与地上生物量和灌丛地上生物量之间均无显著性相关关系。

表2 撂荒草地地上生物量与物种丰富度pearson相关系数
3 讨论
本研究分析结果表明,草地撂荒年限较长,地上生物量越高,这与杜峰等[3]的研究结果基本一致。但是,本研究30 a撂荒草地群落和灌丛地上生物量最高,生物量最高的年份并不是42 a的撂荒草地,可能是群落新增物种生物量高于丢失物种的生物量,如本研究中30 a撂荒草地中出现的猪毛蒿(A.scoparia)和山莓(R.corchorifolius),因其个体生物量较高,导致该年份群落和灌丛的生物量明显增加。而阳坡撂荒草地地上生物量显著高于阴坡,主要归因于不同坡向水、热等条件存在差异,阳坡具有光照优势,阴坡则在水分含量上略占优势[14],进一步说明植物群落地上生物量积累过程中,植物光合作用优势较大。而不同坡向30 a撂荒草地灌丛地上生物量百分比接近,取决于灌木、半灌木与草本植物之间激烈的光照竞争,这弱化了坡向对光照强度的影响[15],造成生物量百分比接近。撂荒草地植物群落与地形因素关系密切,坡向与草地物种分布格局息息相关[16],植物因自身生长习性会天然的选择喜光、向阳、耐阴和湿润的生长环境[17]。本研究结果指出,阴坡撂荒草地灌丛地上生物量百分比高于阳坡草地,从植物群落分布来看,阴坡主要以铁杆蒿(A.sacrorum)、山莓(R.corchorifolius)、黄刺玫(R.xanthina)、柠条锦鸡儿(C.korshinskii)等灌木/半灌木为主。而阳坡则主要以狗尾草(S.viridis)、异叶败酱(P.heterophylla)、野菊(C.indicum)、白羊草(B.ischaemum)等草本植物为主,这体现了植物对不同生境的适应性,具体表现在灌丛地上生物量及其占比的变化上。阴坡撂荒草地灌丛地上生物量及百分比随撂荒年限的增长而增加,这与物种丰富度的变化规律保持一致。
物种丰富度分析结果表明,撂荒年限越长,物种丰富度越高。阳坡撂荒草地30 a植物群落物种丰富度最高,阴坡撂荒草地植物群落物种丰富度随撂荒年限的增长而增加,这与撂荒草地地上生物量的变化规律基本保持一致。而30 a和42 a阳坡撂荒草地植物群落物种丰富度显著高于阴坡,原因在于阳坡草地光照条件更好,草本层接受光照更充足,利于喜阳物种生存,因此物种丰度更高。撂荒年限对灌木和半灌木物种数目无显著性影响,但植物群落中灌木和半灌木物种数目随撂荒年限的增加整体上呈逐渐增加的趋势,可能是随撂荒年限增加,土壤养分富集,灌木和半灌木较草本植物能够获取更多的养分和光照,导致其数量及种类增多[18]。阳坡撂荒草地灌木和半灌木物种数目百分比随撂荒年限的增加而降低,可能是群落物种丰富度随撂荒年限的增加而增加,导致阳坡草地灌木和半灌木的生存空间受影响所致。另外,植物群落灌丛化通常被认为是草地退化的表征,一般会导致物种多样性降低,阴坡撂荒草地灌木和半灌木物种数目百分比高于阳坡草地,这与张荣等[17]的研究结果基本一致。造成这种结果的原因可能是冬季阴坡土壤冻融,水分充足,此时群落中优势物种的株高较低,使得群落内光照分布不均,正好有利于灌木层物种的生长。而阳坡撂荒草地因土壤风化,造成土壤养分难以富集,虽光照充足,但水分蒸发快,抑制了耐阴植物的生长和繁殖[19],导致植物群落灌木和半灌木的百分比较低。由于在一定尺度范围内,受资源和环境异质性的影响,植物群落物种数随取样面积的增加而增加[20],因此不同坡向和撂荒年限草地植物群落物种丰富度均随取样面积的增大而增加。而平衡态拐点出现在不同取样面积上,则归因于植物群落的斑块效应。随撂荒年限增加,群落物种数进入平衡态的数目逐渐增加,但灌木和半灌木物种数目呈先增加后降低的趋势,这与群落物种以及灌木和半灌木物种数目的变化规律基本保持一致。
地上生物量、物种丰富度及撂荒年限之间的相关性结果显示,地上生物量与灌丛地上生物量之间具有极显著正相关关系,灌丛作为植物群落的重要组成部分,其生物量的高低直接影响了群落地上生物量的变化,因此两者之间存在显著正相关关系。物种丰富度、群落及灌丛地上生物量与撂荒年限之间呈显著正相关关系,归因于撂荒年限的延长改变了土壤、小气候及植物群落环境。而物种丰富度增加有助于提高生物量[21],群落地上生物量与优势物种的多度呈正相关,优势物种的改变会导致植物群落生物量及稳定性的改变[22],这就造成了地上生物量和物种丰富度随撂荒年限的增加而呈现出相同的变化规律。有研究结果显示,撂荒年限的增加,优势种发生转变,物种丰富度减少,群落结构趋于简单[23]。但本研究结果指出,物种丰富度随撂荒年限的延长而增加,这可能是由于新出现的优势种受生物环境和非生物环境共同影响,竞争优势不明显,群落优势物种转变并未对群落其他物种间的竞争关系产生显著影响。因每个物种的适应策略不同,从而导致群落物种组成改变,影响到了群落中灌木和半灌木物种数目[24],这也是草地撂荒后群落环境整体向好的一种表现。另外,阳坡植物群落物种丰富度与地上生物量、灌丛地上生物量极显著正相关,原因可能在于灌木和半灌木物种数目百分比降低,但个体生物量增加,导致物种丰富度与灌丛生物量呈正相关关系;虽然灌木和半灌木物种数目百分比降低,但其物种数目随撂荒年限的延长而增加,这与群落物种丰富度随撂荒年限的延长而增加的变化规律一直,也说明了物种丰富度与灌丛生物量呈正相关关系。但阴坡植物群落物种丰富度与地上生物量和灌丛地上生物量之间均无显著性相关关系,可能是由于在植物群落灌丛化过程中,坡向变化未造成物种丰富度和地上生物量的显著变化,但却呈现出相同的变化规律,但在阴坡草地表现不显著[13]。
4 结论
撂荒草地植物群落、物种丰富度和灌丛地上生物量及其百分比随撂荒年限的增加而增加;阳坡草地群落地上生物量及物种丰富度高于阴坡草地;但阳坡草地灌木和半灌木地上生物量低于阴坡草地,且阳坡撂荒草地群落灌木和半灌木物种数目百分比随撂荒年限的增加而降低。撂荒年限与群落地上生物量、灌木和半灌木地上生物量、物种丰富度之间均存在正相关关系,坡向不同引起了植物群落中灌木和半灌木的数量和生物量的变化。
