褪黑素调控草类植物生长发育及抗逆性功能研究进展
2023-10-08聂宇婷崔凯伦吕艳贞闫慧芳
张 昭, 聂宇婷, 崔凯伦, 吕艳贞, 闫慧芳*
(1. 青岛农业大学草业学院, 山东 青岛 266109; 2. 黄河三角洲草地资源与生态国家林业和草原局重点实验室, 山东 青岛 266109;3. 青岛市滩涂盐碱地特色植物种质创制与利用重点实验室, 山东 青岛 266109)
褪黑素即N-乙酰基-5-甲氧基色胺,是一种天然生物活性吲哚胺分子,广泛存在于植物的种子、叶、果实、花和根等组织中,具有调控植物生长发育的功能[1]。褪黑素是一种抗氧化剂,能够清除由非生物和生物胁迫诱导产生的过量活性氧(Reactive oxygen species,ROS),提高抗氧化防御能力,在植物应对干旱、盐、重金属、冷、热、紫外线等逆境胁迫中发挥着重要保护作用[2]。此外,褪黑素还是一种信号分子,能够通过激活胁迫应答基因的表达来增强植物对非生物和生物胁迫的抗性[3]。目前,关于褪黑素在传统农作物、蔬菜等植物生长发育中的生理功能及在抗逆性方面的调控作用研究已较为深入,而其在草类植物中的相关研究却非常有限。本文对近年来褪黑素调控草类植物生长发育及抗逆性的研究进行了总结,重点阐述了褪黑素在草类植物生长发育中的生理功能、非生物和生物胁迫响应中的调控作用,并对其在草类植物抗逆性功能研究和抗逆育种中的应用进行了展望,旨在为利用褪黑素提高草类植物抗逆性以及改善牧草品质提供参考依据。
1 褪黑素调控草类植物生长发育的生理功能
褪黑素作为一种生长调节剂和信号分子,参与调控植物生长发育涉及的多种生物过程,包括种子萌发、根系发育、叶片衰老、开花调节、果实成熟等[4]。
1.1 促进种子萌发
种子萌发是植物整个生命周期中最关键的阶段,也是对环境因子响应最敏感的时期[5]。研究表明,褪黑素在提高种子活力和促进种子萌发方面具有重要作用[6]。外源褪黑素有效缓解了干旱胁迫对无芒雀麦(BromusinermisLeyss.)以及盐胁迫对燕麦(AvenasativaL.)老化种子萌发造成的抑制,显著提高了种子的发芽率,增加了幼苗游离脯氨酸和可溶性蛋白质含量,提高了超氧化物歧化酶(Superoxide dismutase,SOD)活性,降低了丙二醛(Malondialdehyde,MDA)含量[7-9],表明褪黑素对干旱胁迫、盐胁迫和老化处理的种子发芽具有促进作用。外源褪黑素调控种子活力与萌发的生理功能具有浓度和时间依赖性。Yan等[10]采用不同浓度外源褪黑素引发燕麦老化种子,发现最适浓度200 μmol·L-1可通过抑制磷脂酶D表达和磷脂降解修复细胞超微结构,改善线粒体和细胞膜发育,进而增强抗坏血酸-谷胱甘肽(Ascorbic acid-glutathione,AsA-GSH)系统抗氧化能力,降低ROS含量,促进老化种子发芽和幼苗生长。而随后在褪黑素引发不同时间研究中则发现,燕麦种子中12-氧-植物二烯酸还原酶积累,可能通过间接促进茉莉酸(Jasmonic acid,JA)合成,以信号分子参与种子对老化胁迫的氧化防御反应[11]。因此,外源褪黑素改善逆境胁迫和老化对种子萌发的抑制作用主要通过增强抗氧化防御能力、清除多余ROS和修复细胞结构来完成。
1.2 促进根系发育
植物根系发育可直接影响其对土壤水分和养分的吸收能力,发育良好的根增加了与土壤的接触面积,有效地吸收营养物质。褪黑素作为一种植物激素或生长调节剂,以浓度依赖方式对根系结构发育起着重要调节作用,表现为低浓度促进初生根生长,而高浓度虽然抑制初生根生长,但促进侧根发育[12-13]。研究发现,低浓度褪黑素(0.01 μmol·L-1)对拟南芥(Arabidopsisthaliana(L.) Heynh.)初生根伸长的促进作用与吲哚-3-乙酸(Indole-3-acetic acid,IAA)诱导的激素、细胞壁和发育通路相关基因过度表达有关,取决于IAA极性转运的存在;而高浓度褪黑素(100~1 000 μmol·L-1)对初生根生长的抑制作用则与JA、油菜素类固醇(Brassinosteroids,BR)、细胞分裂素(Cytokinin,CTK)、生长素生物合成相关基因表达被抑制有关[14-15]。但是,高浓度褪黑素却促进了拟南芥根系的侧向发育,激活乙烯合成相关基因表达,并与其他激素结合调节锌指蛋白(ZFP)、类钙调蛋白(CML)和NAC基因表达,使IAA外排转运,以抑制初生根生长并促进侧根发育[16]。在拟南芥中,褪黑素可以使不定根增加2倍,侧根增加3倍,而对根毛密度没有影响[17]。褪黑素具有与IAA相似的结构和功能,两者均以色氨酸作为前体物质,诱导白羽扇豆(LupinusalbusL.)中柱鞘细胞产生根原基,促进不定根和侧根的形成[18]。因此,外源褪黑素可能被代谢为IAA或IAA促进剂,从而通过调节植物体内激素水平来促进根系发育[19]。
1.3 延缓叶片衰老
叶片是植物进行光合作用的主要场所,也是衰老比较敏感和容易发生的部位之一[20]。褪黑素作为一种强有力的抗氧化剂,具有延缓叶片衰老的重要作用[21]。Arnao等[22]研究外源褪黑素对缓解大麦(HordeumvulgareL.)叶片衰老的影响,发现叶片中叶绿素损失明显减少,叶片衰老程度得到有效缓解。外源褪黑素还抑制了黑暗诱导的黑麦草(LoliumperenneL.)叶片衰老,使叶绿素降解与衰老相关基因(LpSAG12.1,Lph36和Lpl69)转录水平降低,叶绿素含量和光化学效率提高,离体叶片内源O2-产生速率和H2O2含量显著降低,SOD、过氧化氢酶(Catalase,CAT)活性及转录水平增加,细胞膜稳定性保持良好[23]。由此可见,褪黑素主要通过抑制叶绿素降解、有效减缓光化学效率下降,调节ROS清除系统活性,并维持光系统完整性,从而延缓植物叶片衰老。
1.4 影响花发育及开花过程
植物开花是生命周期中的关键事件,也是营养生长向生殖生长过渡的重要标志[24]。褪黑素是一种黑暗信号分子,可通过叶片的光受体被传导到顶端分生组织,进而诱导植物成花过程[19]。Zhang等[25]研究发现,褪黑素参与了拟南芥花期转变时期独脚金内酯(Strigolactone,SL)下游作用,通过调控开花抑制基因FLOWERINGLOCUSC(FLC)表达来延迟开花,但SL如何调节褪黑素合成机理仍不清楚。此外,DELLA蛋白可以通过不同遗传途径抑制植物开花,而褪黑素处理增加了两种DELLAs蛋白RGA和RGL3的稳定性,并抑制开花基因SUPPRESSOROFOVEREXPRESSIONOFCONSTANT1(SOC1)和FLOWERINGLOCUST(FT)表达[26]。拟南芥中褪黑素水平降低与赤霉素合成减少有关,通过影响FT表达抑制开花,但褪黑素如何调节赤霉素合成还有待进一步研究[27]。Kolár等[28]研究则发现外源褪黑素参与了红叶藜(ChenopodiumrubrumL.)向开花过渡的早期事件,影响了茎尖分生组织中开花的光周期诱导和后续发育过程。目前,关于褪黑素调控草类植物花发育和开花过程的研究还非常有限。
褪黑素作为植物生长调节因子,在草类植物中促进种子萌发、促进根系发育、延缓叶片衰老、调控花发育方面的生理功能研究均已被报道。此外,褪黑素还广泛分布于植物果实中,对果实生长发育、采后成熟及调节昼夜节律等方面具有重要作用。然而,关于褪黑素调控草类植物果实(或种子)发育和昼夜节律的研究还未见报道(图1)。
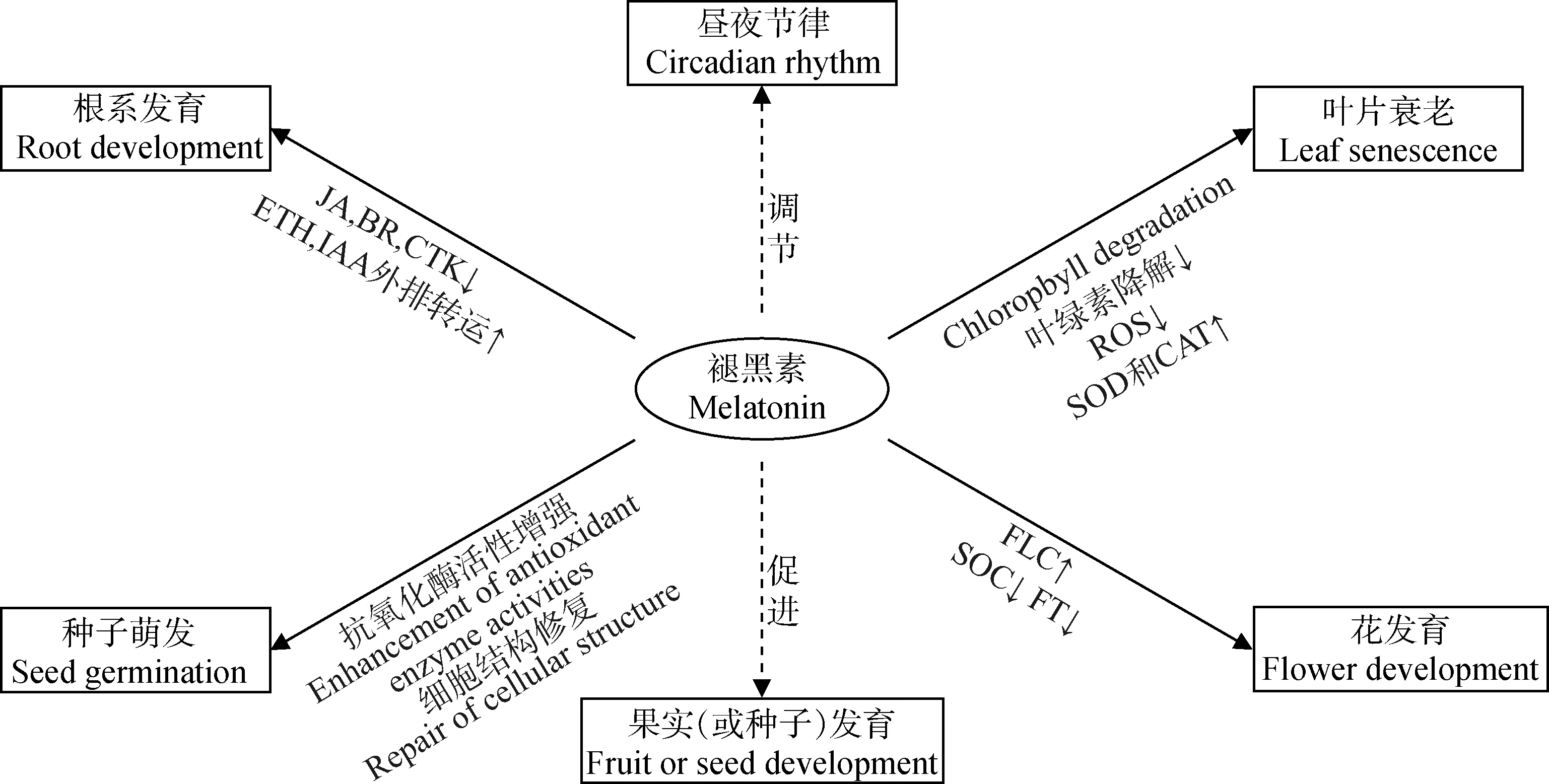
图1 褪黑素调控草类植物生长发育的生理功能
2 褪黑素参与调控草类植物非生物胁迫响应
非生物胁迫是影响植物生长发育的重要限制因素,如高温、寒冷、干旱、盐碱、重金属等,直接或间接影响了植物的生产潜力及营养品质。褪黑素不仅能调节植物的生长发育,还能参与调控植物对多种非生物胁迫的响应[13]。作为一种抗氧化剂,褪黑素通过有效清除胁迫诱导的大量ROS,提高抗氧化酶活性,从而增强植物抵抗逆境胁迫的能力[29]。研究表明,植物内源褪黑素含量在应对非生物胁迫时发生改变,并作为次级信使分子参与调控多种逆境胁迫应答[13]。
2.1 低温胁迫
植物生长发育过程遭遇低温寒冷条件将严重制约其物质代谢及生命活动事件,而褪黑素能够调节植物对低温胁迫的响应,在增强植物低温耐受性方面发挥重要作用。叶片喷施100 μmol·L-1褪黑素显著提高了低温胁迫下多年生黑麦草的幼苗鲜重,降低了叶片相对电导率、MDA含量和ROS积累,提高了CAT、抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)和谷胱甘肽还原酶(Glutathione reductase,GR)活性及脯氨酸和内源褪黑素水平,表明外源褪黑素缓解了低温对幼苗造成的损害,增强其抗寒性[30]。Irshad[31]研究外源褪黑素对蒺藜苜蓿(MedicagotruncatulaGaertn.)抗寒性影响,发现褪黑素提高了硝酸还原酶活性、养分吸收能力、渗透物和叶绿素含量,降低了MDA含量,并通过增强抗氧化能力缓解了低温胁迫造成的不利影响。褪黑素还提高了低温胁迫下狗牙根(Cynodondactylon(L.) Pers.)的抗寒性,降低了其MDA含量和电解质渗漏,提高了叶绿素水平、SOD和过氧化物酶(Peroxidase,POD)活性,增加了糖类化合物(阿拉伯糖、甘露糖、吡喃葡萄糖、麦芽糖和吐鲁糖)含量,并维持了细胞膜完整性[32]。由此可见,褪黑素在草类植物响应低温胁迫中通过增强抗氧化防御系统功能、提高叶片光合效率、维持细胞膜完整性等增强了植物的耐低温能力。
2.2 高温胁迫
随着全球气候变暖,高温胁迫也成为限制植物生长发育的重要非生物胁迫因子之一。植物生长季平均温度每升高1℃,产量将降低17%[33]。前人先后开展了外源脱落酸(Abscisic acid,ABA)、水杨酸(Salicylic acid,SA)、乙烯、CaCl2等对高温胁迫影响的缓解效果,证实了这些物质具有提高植物耐高温性的作用[34-35]。近年来,褪黑素也被证明在增强植物对高温胁迫抗性中发挥重要功能。Zhang等[36]研究发现,外源褪黑素抑制了多年生黑麦草衰老相关基因(LpSAG12.1和Lph36)、ABA生物合成基因(LpZEP,LpNCED1,LpABI3,LpABI5)表达,激活了CTK生物合成基因(LpIPT2和LpOG1)表达,使内源褪黑素和CTK含量升高,从而缓解了热胁迫诱导的生长抑制和叶片衰老。褪黑素还改善了匍匐翦股颖(AgrostisstoloniferaL.)的耐热性,降低了叶片脂质过氧化,提高了SOD,POD活性,改变了脂肪酸含量[37]。高羊茅(FestucaarundinaceaSchreb.)为冷季型草坪草,不耐高温,而褪黑素处理增加了其在热应激条件下的叶绿素含量、总蛋白和抗氧化酶活性,降低了ROS,MDA含量及电解质渗漏,转录组分析发现FaHSFA3,FaAWPM和FaCYTC2显著上调,表明这些基因可能是褪黑素的靶基因[38]。褪黑素通过提高抗氧化防御系统和光合作用系统效率,诱导高温胁迫相关基因表达,有效缓解了热应激对植物造成的损伤。
2.3 干旱胁迫
由全球气候变暖导致的季节性干旱和地域性降水分布不均严重影响了植物从幼苗到开花结果的整个发育阶段,引起植物生长受阻和发育迟缓等问题,进而制约农牧业的高效生产[39]。目前,干旱已成为全球范围内限制植物生长的重要非生物胁迫因子。
褪黑素作为一种抗氧化剂,缓解了干旱胁迫对草类植物幼苗和植株带来的损害。研究发现,叶片喷施褪黑素显著改善了高羊茅苗期根系生长指标(表面积、平均直径、长度、根系活力及侧根数等),提高根冠比和干鲜比,降低叶片相对电导率及MDA含量,提高了幼苗抗旱性[40]。叶片喷施褪黑素还显著提升了干旱胁迫下达乌里胡枝子(Lespedezadavurica(Laxm.) Schindl.)幼苗的地上鲜重、叶片相对含水量、可溶性糖、可溶性蛋白和游离脯氨酸含量,表明褪黑素通过改善叶片水分生理提高了幼苗的抗旱性[41]。同样,叶面喷施褪黑素使多年生黑麦草叶片相对电导率和MDA显著下降,SOD,POD,CAT活性及抗氧化能力显著提高,根和叶片中氮、磷含量增加,表明褪黑素对干旱胁迫下多年生黑麦草抗氧化和养分吸收能力具有一定调节作用[42]。紫花苜蓿(MedicagosativaL.)幼苗和植株的抗旱性也在施用褪黑素后得到明显改善,增强了抗氧化酶活性、叶绿素a、叶绿素b和类胡萝卜素含量,抑制MDA、ROS和活性氮(Reactive nitrogen species,RNS)积累,降低了膜脂过氧化程度,改变了ROS、RNS和脯氨酸代谢相关基因(MsGST7、MsGST17、MsNaDde、MsADH)的转录水平以调控氮氧平衡和脯氨酸代谢[39,43]。褪黑素还增强了草类植物种子萌发期的抗旱性,如褪黑素浸种显著提高了模拟干旱条件下多年生黑麦草种子的发芽势和发芽率[44]。此外,褪黑素对愈伤组织再生能力具有一定影响,如添加褪黑素诱发了模拟干旱胁迫下沟叶结缕草(Zoysiamatrella(L.) Merr.)愈伤组织再生过程中不同抗氧化酶的响应,提高了长期继代培养愈伤组织再生能力[45]。因此,褪黑素能够通过改善根系生长、改善叶片水分生理、提高抗氧化能力、调控氮氧平衡、调节营养物质吸收等过程增强草类植物对干旱胁迫的适应性。
2.4 水涝胁迫
水涝灾害严重威胁植物的生长及产量形成。植物体内,褪黑素与乙烯之间存在拮抗作用,通过抑制乙烯生物合成,从而提高植物对水涝胁迫的适应性[46]。紫花苜蓿在水涝胁迫下的碳水化合物和氨基酸代谢相关途径被激活,乙烯含量显著增加,而适宜浓度外源褪黑素缓解了植株的涝害症状,促进多胺合成,降低乙烯含量,增强细胞膜稳定性,提高光合效率,表明褪黑素通过调控乙烯合成减轻了水涝胁迫对紫花苜蓿的伤害,并增强了其耐涝性[47-48]。目前,关于褪黑素调节草类植物耐涝性的报道还非常有限,未来研究可以探索褪黑素在其他重要草类植物(如燕麦、黑麦草、狗牙根等)响应水涝胁迫中的调控作用。
2.5 盐胁迫
盐胁迫是限制全球农业生产的最主要环境因子,通过渗透胁迫、离子毒害、营养失衡、氧化还原稳态破坏、光合效能受阻等影响植物生长发育,而褪黑素具有改善盐胁迫下种子萌发、幼苗生长和植株发育的功能[49]。研究发现,褪黑素浸种提高了盐胁迫下紫花苜蓿种子的抗氧化酶活性和渗透调节物质含量,降低了脂质过氧化产物MDA含量,增强了盐胁迫下的种子萌发能力[50]。盐胁迫还降低了紫花苜蓿幼苗叶片抗氧化能力和光合作用效率,而外源褪黑素则显著提高了叶片POD、SOD活性,降低了相对电导率、MDA含量,增加了叶绿素a含量、叶绿素b含量、净光合速率、气孔导度和蒸腾速率,表明褪黑素通过提高抗氧化能力和光合作用效率增强了紫花苜蓿幼苗的耐盐性[51-52]。相似结果在褪黑素提高扁穗雀麦(BromuscatharticusVahl.)幼苗耐盐性研究中也被报道[53]。褪黑素还增强了盐胁迫下紫花苜蓿幼苗的1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)和1-氨基环丙烷-1-羧酸氧化酶(1-aminocyclopropane-1-carboxylate oxidase,ACO)含量,表明紫花苜蓿耐盐性增强可能与褪黑素调控乙烯生物合成密切相关[54]。此外,过表达褪黑素生物合成关键基因也提高了植物耐盐性,如过表达耐盐植物白颖苔草(Carexrigescens(Franch.) V.Krecz.)CrCOMT的转基因拟南芥中褪黑素含量增加,盐胁迫下植株的侧根数、脯氨酸含量及叶绿素含量均显著提高[55];分别过表达绵羊AANAT和HIOMT的转基因柳枝稷(PanicumvirgatumL.)植株中褪黑素含量显著提高,对盐胁迫的耐受性增强[56]。因此,褪黑素在一定程度上缓解了盐胁迫对草类植物造成的损害,通过调节抗氧化功能、光合性能及激素水平增强了植物抵抗盐胁迫的能力。
2.6 重金属胁迫
随着现代化城市和工业化进程的快速发展,土壤重金属污染问题已日趋严重。重金属胁迫抑制了植物体内的酶活性,破坏了蛋白质结构,从而使植物出现生长缓慢、植株矮小、产量降低及根系伸长受阻等现象,严重时甚至导致植物死亡[57]。研究表明,褪黑素及其前体物质能够与铝(Al3+)、铜(Cu2+)、镉(Cd2+)、铅(Pb2+)等重金属离子通过螯合作用形成螯合物,从而缓解或消除重金属离子对细胞和生物大分子的伤害[58]。Gu等[59]研究发现,Cd2+胁迫诱导紫花苜蓿根中积累内源褪黑素,而外源褪黑素处理则减少了Cd2+积累,诱导Cd2+转运相关基因表达,维持氧化还原平衡,缓解了Cd2+胁迫对紫花苜蓿生长的影响。同样,喷施褪黑素提高了多年生黑麦草的光合作用,降低了氧化损伤,增加了根中Cd2+螯合态,降低Cd2+向叶片中转移并有效缓解了Cd2+胁迫造成的伤害[60]。Xie等[61]研究褪黑素对Pb2+污染土壤中狗牙根光合性能和抗氧化能力的影响,发现叶片光合色素和叶绿体超微结构得到修复,光合作用和光化学效率得到提高,抗坏血酸(Ascorbic acid,AsA)、还原型谷胱甘肽(Glutathione,GSH)、GSH/GSSG氧化还原状态、AsA-GSH和乙二醛酶系统关键酶活性均被改善,生物量得到提升。此外,外源褪黑素还缓解了Pb2+胁迫对狗牙根种子萌发的抑制作用,提高了种子发芽率[62]。由此可见,褪黑素在草类植物响应重金属离子胁迫时可作为一种良好的保护剂。
2.7 黑暗胁迫
植物长期处于黑暗条件下会导致ROS过度积累和抗氧化清除系统功能减弱,严重影响植物的光合性能和生长发育,而褪黑素能够有效缓解黑暗诱导的植物叶片衰老。Zhang等[23]用外源褪黑素处理黑暗条件下的多年生黑麦草离体成熟叶片,发现叶片仍保持显著更高的内源褪黑素水平、叶绿素含量、光化学效率和细胞膜稳定性,增加了SOD、CAT酶活性和转录水平,而降低了叶绿素降解相关基因和衰老标记基因(LpSAG12.1,Lph36和Lpl69)的转录表达,表明褪黑素可能通过激活抗氧化酶活性和抑制叶绿素降解来缓解黑暗诱导的叶片衰老。目前,关于褪黑素调节草类植物黑暗胁迫响应的报道还非常有限,其相关调控作用及机制还有待进一步研究。
2.8 电离和紫外辐射胁迫
电离和紫外辐射等胁迫导致植物形态结构发生变化,尤其长期强紫外辐射会引发植物细胞膜脂过氧化,产生过量ROS,造成DNA损伤和有丝分裂异常,最终影响植物正常生长发育及生理生化代谢[63]。褪黑素作为一种抗氧化剂,无论内源合成还是外源施加,均能帮助植物清除ROS,减缓电离、紫外辐射诱导的损伤[64]。Afreen等[1]研究发现,高强度UV-B辐射刺激甘草(GlycyrrhizauralensisFisch.)根部合成褪黑素,在一定程度上缓解了UV-B辐射对植物造成的影响。过表达褪黑素合成基因NsSNAT和NsCOMT减缓了UV-B辐射对林烟草(NicotianasylvestrisSpeg. &Comes)的胁迫损伤[65]。此外,Yao等[66]研究发现,外源和内源褪黑素均可调节拟南芥的UV-B信号通路,通过改变通路中AtCOP1,AtHY5,AtHYH和AtRUP1/2基因表达以抵御UV-B诱导的胁迫,调节植物抗氧化防御功能。因此,作为一种抗氧化剂,褪黑素在植物抵御电离和紫外辐射胁迫中发挥了重要作用。
褪黑素是一种重要的吲哚分子,参与调控植物各种发育过程和胁迫响应[67]。前人研究报道了褪黑素参与草类植物的非生物胁迫响应,其相关调控作用包括4方面:①作为抗氧化剂清除自由基。褪黑素具有极强的清除能力,可以直接清除ROS,也可以通过增强其他抗氧化酶活性间接清除多余ROS,维持细胞氧化还原动态平衡[68]。②提高草类植物光合效率。褪黑素参与调控编码叶绿体转录机制基因、维持转录活性和叶绿体功能状态基因的表达,从而调节叶绿体功能以提供有效的光氧化应激保护[69]。③调控植物激素信号。褪黑素通过参与调节IAA,ABA,CTK,JA,乙烯等激素合成代谢在植物响应高温、盐等胁迫中发挥作用[36,54]。④调控胁迫相关基因表达。褪黑素调控草类植物非生物胁迫抗性相关基因包括衰老标记基因(LpSAG12.1,Lph36,Lpl69)、ABA生物合成基因(LpZEP,LpNCED1,LpABI3,LpABI5),CTK生物合成基因(LpIPT2,LpOG1)[36]、脯氨酸代谢相关基因(MsGST7,MsGST17,MsNaDde,MsADH)[42]、抗氧化基因(AtAPX1,AtCATs)[70]等。因此,褪黑素通过多种作用方式从多层面协调发挥功能,参与草类植物的非生物胁迫响应(表1)。

表1 褪黑素参与调控草类植物非生物胁迫响应的作用机制
3 褪黑素参与调控草类植物生物胁迫响应
自然界中多种病原菌以植物作为潜在寄主,包括细菌、真菌和病毒等,它们时刻威胁着植物的生长发育,严重的甚至导致减产,造成经济损失[72]。据报道,动物体内褪黑素有助于产生先天免疫,并调节多种生理功能[73]。近年来,随着植物褪黑素研究的不断深入,其在植物抵抗多种病原菌上也发挥重要功能[74]。
3.1 细菌侵染
细菌能够突破植物免疫监控而侵入宿主体内,通过分泌系统将效应蛋白运送到宿主细胞直接攻击植物或运送到环境中间接影响植物[75]。目前,已有研究证实褪黑素可以提高植物对细菌的局部耐受性。郭苗杰[75]研究褪黑素在丁香假单胞杆菌侵染拟南芥的抗菌功能中发现,褪黑素可通过促进AGD2样防御反应蛋白 1(ALD1)、SA诱导缺陷 2(SID2)、系统性获得性抗性缺陷 1(SARD1)和钙调素结合蛋白 60 g(CBP60 g)防御基因的表达,将防御信号传递给SA下游PR1、PR5基因,增强拟南芥局部防御细菌的能力。此外,褪黑素还通过MAPK级联反应、诱导NO产生来增强拟南芥抵御丁香假单胞杆菌侵染的抗性[76-77]。
3.2 真菌侵染
真菌种类繁多,常见的有灰霉菌、芸薹根肿菌、锈菌等[78]。近年来,褪黑素被报道具有提高植物对真菌耐受性的作用。祝颖[79]研究发现,灰霉菌侵染引起拟南芥ROS爆发,内源褪黑素含量增加并促进JA产生,从而保护植物免受灰霉菌的损伤,表明内源褪黑素提高了拟南芥对灰霉菌的防御能力。此外,研究还发现外源褪黑素处理降低了拟南芥根部芸薹根肿菌孢子囊数目和发病程度,提高了JA途径中PR3、PR4基因和SA途径中PR1、PR2、PR5基因表达,表明外源褪黑素提高拟南芥对芸薹根肿菌的抗性主要通过JA和SA信号途径完成[80]。因此,内源和外源褪黑素均能以激素信号形式调节相关基因表达,从而在植物抵抗生物胁迫中发挥重要作用。
3.3 病毒侵染
病毒是由核酸和蛋白质外壳构成的病原体,依靠寄生在宿主内存活。目前,能够侵染植物的常见病毒有烟草花叶病毒、苜蓿花叶病毒、黄瓜花叶病毒等,其中烟草花叶病毒是研究较多的一种植物病毒[81]。褪黑素被发现可以增强植物对病毒的抗性,如褪黑素通过提高抗氧化酶活性、减轻过氧化损伤增强了埃及茄科植物对苜蓿花叶病毒侵染的抵抗能力[82],褪黑素与NO协同作用提高了水稻(OryzasativaL.)对水稻条纹病毒的抗性[83]。然而,有关褪黑素增强草类植物对病毒抗性的研究还未见报道,尤其紫花苜蓿、燕麦、羊草(Leymuschinensis(Trin.) Tzvel.)、高羊茅、草地早熟禾(PoapratensisL.)等重要牧草和草坪草。因此,在未来研究中,可以关注褪黑素在这些草类植物抗病中的作用。
4 展望
植物褪黑素含量极低,但其在植物应对各种非生物胁迫(干旱、盐、低温、高温、重金属等)和生物胁迫中却发挥着至关重要的作用[30,36,45,50,61,80]。随着植物褪黑素研究的不断深入,其调控植物生长发育的生理功能及作用机制也被逐渐揭示。然而,相比于传统农作物和经济作物,褪黑素调控草类植物生长发育及抗逆性功能的研究还非常有限,仍存在一些需要深入探究和解决的问题。
4.1 褪黑素调控草类植物生长发育的生理作用需深入探究
作为植物生长调节剂,褪黑素在促进种子萌发、促进根系发育、延缓叶片衰老、影响花发育及开花过程等方面研究取得重要进展,但其调控草类植物昼夜节律、果实(或种子)发育等机制尚不明晰。此外,褪黑素与其他植物激素相互作用的信号途径仍不清楚。因此,未来研究仍需进一步探究并解析褪黑素与其他植物激素协同调控草类植物生长发育的作用机制。
4.2 褪黑素调控草类植物抗逆性的功能及信号通路需进一步明确
褪黑素参与调控草类植物应对极端温度[30-32,36-38]、干旱[39-43,45]、水涝[46-47]、盐[50-54,71]、重金属[61]、黑暗[23]、电离和紫外辐射[1,64-66]等非生物胁迫以及细菌[75-77]、真菌[79-80]感染等生物胁迫的研究均已被报道,其作用机制也得到部分揭示。然而,褪黑素在增强草类植物调节营养失衡、防御病毒感染方面的研究还未见报道,尤其在紫花苜蓿、燕麦、高羊茅等重要牧草和草坪草中。植物应对生物胁迫时,褪黑素能够通过水杨酸、茉莉酸、MAPK、NO信号途径诱导防御相关基因表达,未来可重点开展其在植物应对非生物胁迫时诱导抗性相关基因表达的具体信号通路方面的研究。
4.3 褪黑素合成基因的功能研究与其在抗逆新品种培育中应用需融合发展
过表达褪黑素生物合成基因可显著提高植物内源褪黑素水平,增强转基因植株对多种逆境胁迫的抗性[55-56,65]。随着测序技术的快速发展,利用现代分子育种手段调控草类植物褪黑素生物合成基因表达,适度范围内提高褪黑素含量,改善草类植物品质及对重要环境因子的抗性。因此,结合传统育种理论和经验,培育高褪黑素含量的新品种是一个具有挑战性的新方向,将对草类植物抗逆新品种培育具有重要意义。
