合理施磷量提高水稻拔节期对大气增温的抵御能力
2023-09-26唐煜杰乔云发苗淑杰
唐煜杰,苗 欢,冯 茜,乔云发,苗淑杰
(南京信息工程大学应用气象学院,江苏南京 210044)
水稻是我国主要粮食作物之一,种植面积较大,约为2.94×107hm2[1],全国有一半以上的人口以稻米为主食。但我国人均耕地面积较少,因此,提高水稻单产是保障我国粮食安全的重要手段[2]。然而,全球变暖(IPCC 预测21 世纪末全球温度相比于工业革命时期升高1.5℃以上[3])对水稻生长的影响日益凸显。针对这一问题,国内外学者采用了不同方式研究增温对水稻生长发育的影响。研究结果表明,室内栽培条件下增温促进水稻生长,提高光合速率和养分含量[4-5]。然而,开放式田间增温系统下的研究结果存在分歧。张佳华等[6]发现,增温提高水稻叶绿素含量和生物量,而其他田间研究则表明增温可能会降低水稻的光合速率和生物量[7-8]。此外,Liu 等[9]通过作物模型的建模方式发现,增温可提高水稻生物量。因此,目前关于增温对水稻生长的影响仍无法给出明确结论。
磷素是水稻生长和发育所必需的营养元素之一。低磷环境下,水稻光合磷酸化速率下降,光合产物减少,同时光合产物运输也受到抑制[10]。而施用磷肥促进水稻根系的延伸和根毛的形成,增强水稻的生理活性和抗逆性,从而提高产量[11-12]。研究发现,施磷可以促进水稻对磷的吸收[13],同时有利于水稻吸收其他养分如氮和钾[14]。另一方面,有研究表明,一定程度的低磷处理对水稻养分积累更有效[15],而过量施磷则可能抑制水稻对养分的吸收[16]。此外,施磷后水稻的生长指标如叶面积比和有效分蘖数均有提高,同时水稻产量也显著增加[17]。然而,研究也表明过量施磷会降低水稻产量[18-19]。因此,优化施磷是促进水稻生长发育的有效措施。
拔节期水稻代谢旺盛,养分吸收量大,是承前启后的关键时期,拔节期合理的施肥措施是实现产量提高的基础[20]。已有关于增温或施磷单因素对水稻生产影响的研究,但缺乏双因素综合分析,尤其对拔节期水稻的磷肥需求还不清楚。因此,本研究选择‘陵两优268’ (籼型两系杂交稻)作为研究材料,采用红外增温模拟系统,使对照组大气增温1.5℃,通过设置不同的施磷水平,研究拔节期水稻的光合特性、养分吸收和生物量积累对增温的响应,以期为未来水稻生产应对全球变暖的磷肥施用技术提供理论支撑。
1 材料与方法
1.1 试验地点与材料
1.1.1 试验地点 本试验于2020 年5 月至8 月在南京信息工程大学农气试验站进行(32°12′23″N,118°42′47″E)。在地理区域上属于亚热带季风气候区,年降水量为1105 mm,相对湿度为78%,年平均气温为15.5℃,无霜期236 天。
1.1.2 供试土壤与水稻品种 供试品种为‘陵两优268’,为籼型两系杂交稻。全生育期112 天,株高87.7 cm,抗性强。供试土壤为江苏典型黄棕壤,试验前表层土壤(0—15 cm) 基本理化性质如下:pH 6.2、有机质19.4 g/kg、全氮1.45 g/kg、碱解氮80.3 mg/kg、速效磷16.2 mg/kg、速效钾113 mg/kg、粘粒含量 26.1%、田间持水量 27%、容重1.25 g/cm3。
1.2 试验设计
试验采用双因素随机区组设计,盆栽方式培养,容器为直径25 cm、高30 cm 的塑料桶,装土10 kg/盆。温度条件设为增温1.5℃ (eT)与对照大气温度(aT)。红外加热器(MR-2420)对增温处理进行全天24 h 增温,加热器附带的温度记录仪监测全天温度,日均增温效果如图1 所示。设置5 个P2O5施用水平:0、30、60、90、120 mg/kg,记为P0、P30、P60、P90和P120,处理P30、P60、P90和P120分别相当于农业农村部推荐施肥量(P2O575 kg/hm2)[15,21]的1、2、3 和4 倍。

图1 水稻生育期内平均日温度的变化Fig.1 Changes of average daily temperature during rice growth period
供试水稻种子经过温汤浸种以及适宜环境条件下催芽后,于5 月25 日使用基质土(蛭石∶田园土∶有机肥=1∶1∶1)在温室苗床育苗(温度22℃~28℃,湿度45%~55%)。在移栽前(6 月22 日)施用基肥,每桶添加尿素(含N 46.7%,分析纯) 0.2 g/kg,土;依据磷处理,用磷酸二氢钾(含P2O5≥52.0%,K2O 34.0%,分析纯)提供磷;以硫酸二氢钾(K2O≥50%,分析纯)补充钾含量至K2O 80 mg/kg, 土。施肥后压实达到田间自然容重水平,然后灌水,养土。待秧龄达到30 天后(6 月24 日)进行移栽,设置每桶3 穴,双本移栽方式,3 次重复,共计30 个样本。移栽7 天后,每桶施入0.5 g 尿素作为追肥,在水稻生长期间保持水层高度在2 ~ 3 cm。
1.3 测试项目与方法
1.3.1 光合参数的测定 在观察到第一株水稻进入拔节期后(基部第一个生长节长度达到1 cm),记录日期并加大观察频次(7 月24 日)。待水稻进入拔节中期后(50%以上水稻进入拔节期,7 月30 日),在天气晴朗的条件下(7 月31 日,8 月3 日,8 月6日),于日间8:00—11:00 用LI-6400 便携式光合仪测定水稻叶片的光合参数,主要包括净光合速率(Pn)、气孔导度 (Gs)、胞间CO2浓度(Ci) 和蒸腾速率(Tr)。测定时选择每株水稻叶片中最长的3 枚叶片进行测定,计算平均值作为一个重复。光合测定条件设置如下:内置光源光强为CO21000 μmol/(m2·s)、环境气温为 (22.3±0.5)℃、温度为26℃、叶室相对湿度为 (50.0±2.0)%、参比室CO2浓度为350 µmol/mol。其中8 月6 日可能由于风速较大导致测定数据出现异常波动 [净光合速率在CO230 μmol/(m2·s) 以上],故不做参考,而剩余两次测定的光合参数表现相同的变化趋势,故选择其中一次测定数据作为研究结果展示。
1.3.2 生物量的测定 于8 月7 日采集水稻样品,收获后将其分为地上部和根部,用自来水清洗3遍,蒸馏水润洗两遍,再用滤纸吸干表面水分,随后在105℃条件下杀青30 min,然后在70℃烘干至恒重。用天平对烘干样品进行称重。
1.3.3 氮磷钾含量的测定 烘干样品用万能粉碎机进行粉碎,经浓H2SO4-H2O2消煮后,氮含量用凯氏定氮法测定,磷含量用钼锑抗比色分光光度法测定,钾含量用火焰光度法测定[22],其中地上部和根部养分含量分开进行测定。部分参数计算方法如下:
根冠比=根生物量/地上部生物量
总生物量 (g)=地上部生物量+根生物量
养分总量 (g/kg)=(地上部养分含量×地上部生物量+根养分含量×根生物量)/总生物量
1.4 数据分析
用Microsoft Excel 2013 软件整理试验数据,对温度和施磷两因素分组,用SPSS 25.0 软件“描述统计”选项卡中的“探索”功能,对水稻各变量进行方差齐性检验(Levene-test) 和正态分布(Shapiro-Wilk test)检验,观察到两种检验结果P>0.05 后,认为满足符合方差齐性和正态分布。用 “一般线性模型”中的“单变量”功能进行双因素方差分析(twoway ANOVA, LSD 多重比较),用“相关”中的“双变量”功能进行Pearson 相关性分析。然后在不同温度条件下,用“回归”中的“线性回归”功能进行水稻生物量的通径分析,因素构成包括净光合速率、氮磷钾养分总量。在通径分析中剔除了P120处理,因为研究结果表明P120处理对水稻生长表现出负效应。
2 结果与分析
2.1 增温和施磷对水稻光合特性参数的影响
2.1.1 水稻净光合速率 增温和施磷对水稻净光合速率(Pn)有极显著影响(P<0.01,表1)。无论增温与否,水稻Pn随施磷量的变化均表现出单峰趋势,峰值出现在P90处理,但P90与P60水稻Pn无显著性差异(表1)。P0和P30处理下,增温显著提高水稻Pn,分别比常温处理提高9.7%和11.0% (P<0.05)。进一步增加磷水平,增温处理水稻Pn与常温无显著差异。

表1 不同磷素水平和增温下水稻光合特性参数Table 1 Photosynthetic characteristic parameters of rice under different phosphorus levels and temperatures
2.1.2 水稻气孔导度和蒸腾速率 增温和施磷对水稻气孔导度(Gs) 和蒸腾速率(Tr) 有极显著影响(P<0.01),且二者交互作用对水稻Tr有极显著影响(P<0.01,表1)。无论增温与否,水稻Gs和Tr随施磷量的增加也表现出单峰变化趋势,峰值出现在P90处理,除增温条件下,P90与P60处理之间水稻Gs无显著性差异外,其余处理组合之间Gs和Tr差异显著。除P90处理外,增温显著提高水稻Gs,且比常温处理提高5.6%~12.2% (P<0.05)。同时,增温显著提高除P120处理外水稻Tr,且比常温处理提高2.3%~8.3% (P<0.05)。
2.1.3 水稻胞间CO2浓度 增温和施磷对水稻胞间CO2浓度(Ci)有极显著影响(P<0.01),且二者交互作用对水稻Ci有极显著性影响(P<0.01,表1)。常温和增温条件下,施磷对水稻Ci的影响规律相反,分别在P90处理取得最小值和最大值,且P90与P60处理下水稻Ci无显著差异(表1)。除P120处理外,增温显著提高其余施磷水平水稻Ci,且比常温处理提高2.7%~6.0% (P<0.05)。
2.2 增温和施磷对水稻养分含量的影响
2.2.1 水稻磷含量 增温对水稻地上部和根部磷含量有极显著影响(P<0.01),施磷仅对水稻根部磷含量有极显著影响(P<0.01),二者交互作用分别对水稻地上部和根部磷含量有显著(P<0.05) 和极显著影响(P<0.01,表2)。随着施磷量的增加,水稻地上部和根部磷含量均显著提高(P<0.05)。与常温相比,增温对P0、P30和P60处理地上部含磷量无显著影响,但显著提高了P90和P120处理地上部磷含量(P<0.05);增温显著提高P0和P120处理下水稻根部磷含量,分别比常温处理增加5.6%和5.8% (P<0.05)。
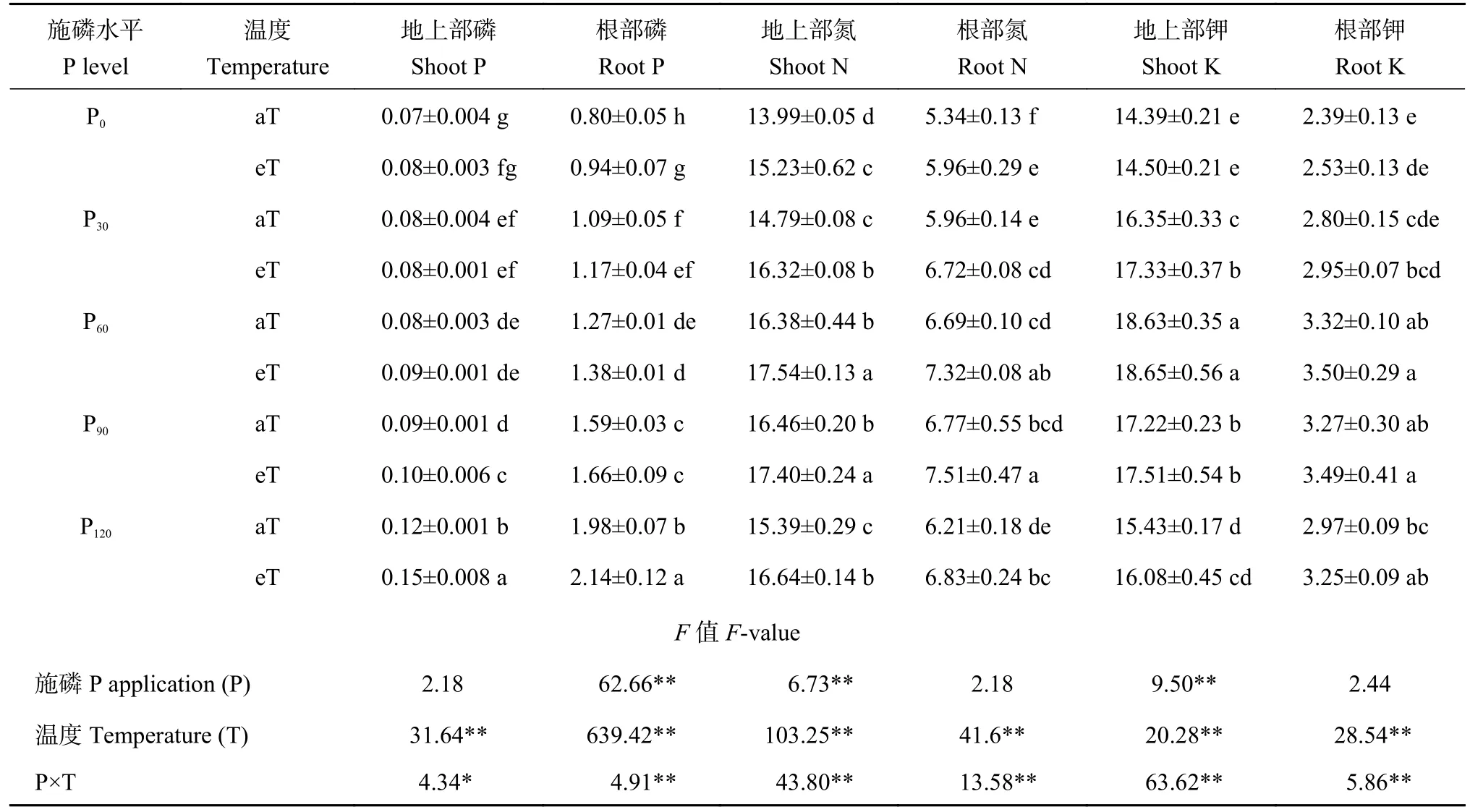
表2 不同磷素水平和温度下水稻地上部和根部养分含量(g/kg)Table 2 Nutrient contents in shoots and roots of rice under different phosphorus levels and temperatures
2.2.2 水稻氮含量 增温对水稻氮含量有极显著性影响(P<0.01),施磷仅对水稻地上部氮含量有极显著影响(P<0.01),二者交互作用对水稻氮含量有极显著影响(P<0.01,表2)。无论是否增温,水稻地上部和根部氮含量随着施磷水平增加呈现单峰曲线变化趋势,其中常温条件下水稻地上部和根部氮含量均在P90处理达到峰值,增温条件下水稻地上部和根部氮含量分别在P60和P90处理达到峰值,且P60与P90处理下水稻地上部和根部氮含量无显著差异(表2)。与常温相比,增温显著提高各施磷水平下水稻地上部和根部氮含量,且分别比常温处理提高5.7%~10.3%和9.4%~12.8% (P<0.05)。
2.2.3 水稻钾含量 施磷对水稻根部钾含量无显著影响,对水稻地上部钾含量影响显著而增温对水稻地上部和根部钾含量影响均极显著(P<0.01),施磷与增温对植株地上部和根部钾含量也有显著交互效应(P<0.05,表2)。不论是否增温,水稻地上部和根部钾含量均随着施磷水平的增加呈现单峰曲线变化趋势,常温条件和增温条件下,水稻地上部和根部钾含量均在P60处理达到峰值,且增温条件下,P60与P90处理下水稻根部钾含量无显著差异(表2)。
2.3 增温和施磷对水稻生长的影响
2.3.1 水稻生物量 增温和施磷对水稻生物量有极显著影响(P<0.01,表3)。无论是否增温,水稻生物量随着施磷水平的增加呈现单峰曲线变化趋势,在P90处达到最大值,且增温条件下,P60与P90处理下水稻地上部生物量无显著差异(表3)。与常温相比,增温显著降低P0和P90处理下水稻总生物量,分别比常温处理降低5.7%和4.2% (P<0.05)。增温显著降低各磷水平下水稻地上部生物量,比常温处理降低6.8%~10.9% (P<0.05),但显著提高了P30和P60处理下水稻根生物量,分别比常温处理提高14.3%和7.4%(P<0.05)。

表3 不同磷素水平和温度下水稻生物量和分蘖数Table 3 Biomass and tiller number of rice under different phosphorus levels and temperatures
2.3.2 水稻根冠比和分蘖数 增温和施磷对水稻根冠比和分蘖数有极显著影响(P<0.01,表3)。无论增温与否,随着施磷水平增加,水稻根冠比表现出不断提高的趋势,并在P120达到最大值。而水稻分蘖数在常温条件下表现为单峰曲线变化趋势,并在P90达到峰值,在增温条件下表现出先提高后在P90处理下趋于平稳的趋势(表3)。与常温相比,增温显著提高各施磷水平下水稻根冠比,比常温处理提高12.3%~22.1% (P<0.05)。而水稻分蘖数对增温不敏感,增温仅显著提高P120水平下的水稻分蘖数,且比常温处理提高8.5% (P<0.05)。
2.3.3 水稻净光合速率(Pn)和养分含量对总生物量的相关性分析与通径分析 从相关性分析(表4)可知,在P30~P90范围内,常温条件下水稻体内氮含量(TN)与钾含量 (TK)、Pn、生物量 (TB) 之间的相关系数分别为0.719、0.754、0.742,达到显著正相关水平(P<0.05);植株磷含量 (TP)与Pn、TB 之间的相关系数分别为0.816 和0.946,Pn与TB 之间的相关系数为0.942,均达到极显著正相关水平(P<0.01)。通径分析结果表明,Pn、TN、TP 对TB 具有直接促进作用,通径系数分别为0.554 (P<0.01)、0.309、0.308,而TK 对TB 具有抑制作用,但直接通径系数仅为-0.235,抑制作用不显著。此外,TN 和TP 通过Pn对TB 的间接通径系数较大,分别为0.417 和0.452。

表4 施磷30—90 mg/kg 范围内水稻净光合速率、养分含量与生物量的相关性分析 (n=12)Table 4 Correlation between Pn, nutrient contents and biomass of rice within P application rate of 30-90 mg/kg
增温背景下,水稻体内TN 与TP、TB 间的相关系数分别为0.689、0.765,另外Pn与 TB 间的相关系数为0.745,均达到显著正相关水平(P<0.05)。TP与TB 之间的相关性系数为0.931,达到极显著正相关水平(P<0.01)。与常温不同,增温背景下,TP 对TB 的直接通径系数为0.794 (P<0.01),其余因素TN、TK、Pn对TB 的直接通径系数较小,分别为0.126、0.104 和0.096。此外,TN、Pn通过TP 对TB 的间接通径系数分别为0.547 和0.528,说明TN、Pn通过TP 对TB 的间接影响不可忽略(表4)。
3 讨论
3.1 增温背景下施磷对水稻光合特性的影响
增温对水稻净光合速率(Pn) 的影响因供磷水平不同而有差异。本研究中,施磷量低于P2O590 mg/kg 时,红外增温1.5℃显著提高水稻Pn(P<0.05),这与袁嫚嫚等[23]利用热水增温管道增温1℃~2℃,发现灌浆期水稻Pn有下降趋势的结果不一致,而与谢立勇等[24]通过覆膜增温约1.5℃,发现拔节期水稻Pn有升高趋势的结果一致。因此,增温方式、增温幅度以及测定的生育期会影响水稻Pn对增温的响应。此外,增温也提高水稻气孔导度(Gs)蒸腾速率(Tr)和胞间CO2浓度(Ci),说明增温直接影响叶片气孔的开闭,并影响水分和CO2进出叶片,从而影响水稻Pn[25]。而随着施磷量的增加,增温对水稻Pn影响减小,仅表现出提高的趋势(P>0.05),这种变化可能是由于施磷显著提高水稻Gs和Tr,解除了气孔对水稻Pn的限制,从而使水稻Pn几乎不受增温影响。施磷量超过P2O590 mg/kg 后,水稻Pn显著下降(P<0.05),这与林诚等[26]冷浸田水稻施磷的研究结果一致。这是由于施磷过多降低水稻Gs、Tr以及蔗糖磷酸合成酶活性,使蔗糖合成受到抑制,从而使水稻Pn降低[27]。这些结果表明,施磷量低于P2O590 mg/kg 时,施磷协同增温共同提高水稻Pn,施磷量超过P2O590 mg/kg,施磷降低水稻Pn。
3.2 增温背景下施磷对水稻养分含量的影响
增温对水稻养分含量的影响因不同供磷水平有差异,并且不同养分响应规律不一致。研究结果表明,施磷量低于P2O590 mg/kg 时,增温提高水稻磷含量,这是由于一方面磷酸盐吸收速率与温度大小成正比[28],另一方面增温提高水稻净光合速率,进而提高磷酸盐吸收速率[29]。增温提高水稻氮含量, 这与常少燕等[30]研究结果一致,这要归因于增温促进土壤氮素矿化过程,提高了氮素供应能力,从而为水稻提供了更多的有效氮[31]。此外,大部分处理下水稻钾含量表现出对增温的正响应,原因可能是施磷增强水稻根系活性,促进水稻对养分的吸收[32]。施磷量超过P2O590 mg/kg 后,增温显著提高水稻磷含量(P<0.05),这可能是增温促进水稻对磷的奢侈吸收,并且这一结果在燕麦[33]以及大麦[34]中也有所体现。同时,正因为水稻根系奢侈吸收磷素,降低了其对氮钾元素的吸收。这与Salifu 等[16]过量施磷降低成熟期水稻磷含量的结论不同,可能是由于本研究水稻处于拔节时期,对磷的需求量较高[35],促进了水稻对磷的吸收。另外,超出一定施磷范围后,水稻氮磷钾含量不再呈现正相关关系,这一结果已被相关研究[14]证实。这些结果表明水稻养分含量对增温和施磷响应结果不同,增温促进水稻养分含量提高,施磷量超过P2O590 mg/kg,施磷协同增温提高水稻磷含量,而降低水稻氮、钾含量。
3.3 增温背景下施磷对水稻生长的影响
增温对水稻生长的影响因供磷水平不同而有差异。本研究结果表明增温不利于水稻生长,表现为总生物量降低。李克南等[36]认为增温提高北方地区水稻生物量,但林娇蓉等[19]认为增温缩短水稻生育期,进而降低生物量。这些结果表明,增温对水稻生长的影响因气候区域而有差别。本研究中增温抑制水稻生长,原因可能是增温背景下,水稻生育期内日均温度大于30℃天数占比较大,而水稻生育前期最适生长温度在25℃~30℃,从而表现对水稻生长的抑制。当施磷量低于P2O560 mg/kg 时,水稻总生物量仅有下降趋势,但下降不显著(P>0.05),这可能是磷参与水稻体内代谢过程,并提高水稻抗逆性从而减轻增温对水稻生长的抑制。而施磷量为P2O590~120 mg/kg 时,磷素的作用效果降低,这与张国盛等[37]在干旱环境下研究磷对水稻生长的影响结果一致,这可能是由于水稻过量吸收磷素提高水稻根冠比,减少光合面积,最终降低水稻总生物量。当施磷量达到P2O5120 mg/kg 时,虽然水稻总生物量仅有下降趋势(P>0.05),但此施磷处理下水稻总生物量较低,并且差异不显著的原因可能是过度施磷的抑制效果强于增温对水稻生物量的影响。这些结果表明,施磷量低于P2O560 mg/kg 时,施磷缓解增温降低水稻生物量的不利影响,超过该施磷量时,磷素作用效果下降。
3.4 常温和增温背景下水稻光合作用、养分含量对总生物量的贡献
常温和增温背景下,水稻光合参数、养分含量对总生物量的贡献不同。相关性与通径分析结果表明,常温条件下,Pn是影响水稻总生物量形成的主控因素,TN 和TP 的作用次之,且是通过Pn而产生的间接作用。增温条件下,水稻通过TP 的直接作用,以及TN 和Pn通过TP 的间接作用来增加水稻拔节期的生物量积累。两种温度条件下,影响水稻生物量积累的主控因子不同的原因可能在于,全天24 h增温加强日间光合作用,同时也加强夜间呼吸作用,加速光合产物消耗,最终致使日间光合作用增强对生物量积累的效果并不明显。这一结果与王小宁等[38]对水稻增温处理的研究结果一致,另外,增温背景下水稻蒸腾速率加强,养分转运加快,从而使水稻养分得到及时补充,进一步提高了磷的有效性[39]。这些结果表明,常温条件下,施磷时应注重提高水稻Pn,进而提高水稻的生物量。增温条件下,施磷时应注重磷素有效性,通过提高磷素有效性缓解增温对水稻生产的不利影响。
4 结论
合理施用磷肥是应对大气增温条件下水稻粮食安全问题的重要措施。常温条件下,施磷量在P2O560~90 mg/kg (约相当于P2O575~110 kg/hm2)时,显著提高水稻叶片净光合速率、气孔导度和蒸腾速率,有利于水稻根系对氮、磷、钾养分的吸收。增温显著提高相同磷水平下水稻叶片光合参数和氮含量,而磷和钾含量变化对增温不敏感,此外,增温在提高根生物量的同时却降低地上部水稻生物量和总生物量。本研究中,施磷量为P2O590 mg/kg (约相当于110 kg/hm2),水稻拔节期光合作用强,养分吸收量大,增加根部生物量,总生物量也达到最高值,还可抵消增温对光合效率的不利影响,为后期水稻生产打下基础。因此,为应对大气增温,江苏稻区在升温条件下应将磷肥(P2O5)施用水平保持在110 kg/hm2,充分利用升温提高水稻吸收利用磷能力的契机,提高磷肥利用效率。
