茶叶氨基酸合成代谢及转化研究进展
2023-09-20黄芳钱佳佳傅秀敏
黄芳 钱佳佳 傅秀敏

摘要:氨基酸是茶叶重要的呈味物质,其在茶叶加工过程中也会发生各种酶促或非酶促反应而转化为茶叶香气及色素的重要组成物质。文章主要综述了近年来不同茶树种质资源、环境条件和加工工艺对氨基酸合成代谢及转化影响的相关研究进展,以为茶叶品质形成与调控提供一定参考。
关键词:茶叶;茶叶加工;环境;氨基酸合成代谢;氨基酸转化
中图分类号:S571.1;Q517 文献标识码:A 文章编号:1000-3150(2023)09-10-9
Research Advances in Amino Acid Metabolism and
Transformation of Tea Plants (Camellia sinensis)
HUANG Fang1,2,3, QIAN Jiajia1,2, FU Xiumin1,2,3*
1. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China;
2. South China National Botanical Garden, Guangzhou 510650, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
Abstract: Amino acids are important flavor substances in tea, and they are converted into important constituents of tea aroma and pigment by various enzymatic or non-enzymatic reactions during tea processing. This paper mainly summarized the recent research progress of amino acid metabolism and transformation in different tea germplasm resources, production environments and post-harvest processing of tea. It would provide a reference for the formation and regulation of tea quality in the future.
Keywords: Camellia sinensis, tea processing, environment, amino acids metabolism, amino acids transformation
氨基酸是貢献茶叶风味的一类重要物质,如茶氨酸具有鲜爽味,谷氨酸、丙氨酸具有类似花香气味。目前已在茶叶中发现26种氨基酸,包括20种蛋白源氨基酸和6种非蛋白源氨基酸,其中非蛋白源氨基酸茶氨酸的含量可占总氨基酸含量的50%以上,其次为谷氨酸、精氨酸、丝氨酸和天冬氨酸等[1]。茶叶氨基酸的含量在不同茶树品种中存在差异,在不同组织中分布也不同,一般嫩叶组织中含量较高,老叶中则较低,且受外界环境影响[2-3]。由于氨基酸本身就是滋味物质和香气物质,因此茶鲜叶中氨基酸的组成和含量不同,影响着茶叶品质。另外,大量研究表明,氨基酸在茶叶不同加工工艺中会发生一系列的氧化、降解等转化反应,产生的一些化合物可影响茶叶品质(色、香、味)[4]。本文主要对茶树种质资源、环境条件及不同加工技术对茶叶氨基酸的合成代谢及其转化影响的相关研究进展进行综述。
1 茶树种质资源对茶叶氨基酸合成代谢及转化的影响
研究发现氨基酸尤其是茶氨酸的含量在不同的茶树品种中存在差异,主要是黄化或白化品种中茶氨酸含量比绿色叶品种高[5-8]。Liu等[8]比较茶氨酸代谢途径基因表达量,发现CsTS2、CsGS1和CsGDH2基因的表达量与品种间茶氨酸的含量呈正相关。Cheng等[6]通过稳定同位素示踪试验表明较弱的茶氨酸水解能力可能导致了黄化或白化品种中高茶氨酸的积累。Fu等[9]首次鉴定了在体外具有水解茶氨酸功能的酶CsPDX2.1,并发现其基因的表达量在黄化或白化品种中较低。近年来,随着测序技术的发展,研究者可通过群体基因组学分析挖掘不同种质资源间关联氨基酸含量差异的位点。如Fang等[10]分析了春、夏、秋三季191份茶树种质资源的茶氨酸含量,并进行了全基因组关联分析,发现7个候选的SNP位点与茶氨酸含量存在显著相关性。Huang等[2]以正常氨基酸含量的龙井43品种和高氨基酸含量的白鸡冠品种为亲本,得到了198份茶树F1人工分离群体,并构建了茶树的高密度遗传图谱,通过两年的数据分析鉴定到了4个与茶叶游离氨基酸(茶氨酸、谷氨酸、谷氨酰胺、天冬氨酸和精氨酸)相关的QTL位点。有趣的是,上述发现的SNP位点和QTL位点均不是目前已发现的氨基酸代谢途径相关基因。
此外,茶氨酸含量在组织部位的分布也有差异,一般在绿色叶品种茶树中,茶氨酸含量在根组织中最高,其次是茎、嫩芽、嫩叶和老叶,但Xu等[7]发现中黄3号和白叶1号嫩芽的茶氨酸含量最低,随着一叶、二叶、三叶叶位递增,芽叶中茶氨酸含量与绿色叶品种呈现相反的趋势。因此建议在加工和利用黄化或白化品种茶树资源时,可以考虑采摘成熟度稍高的芽叶,如一芽三叶或一芽四叶,以达到最佳利用效果,同时也可避免资源浪费。另外,茎组织中茶氨酸含量要比叶组织高,所以根据不同部位开发不同的茶叶产品,可以充分利用资源。
2 产地环境对茶叶氨基酸合成代谢及转化的影响
氨基酸的代谢和积累除了在不同的茶树种质资源上存在差异外,还受外界环境的影响,尤其受不同季节的影响。如春季采摘的茶叶中氨基酸含量要明显高于夏季茶叶。Li等[11]收集了从冬季到春季的4个时间点的13个茶树品种的根和嫩梢组织,并对其进行了茶氨酸含量的分析。发现在茶树根组织中,12月12日、3月1日的样品茶氨酸含量比较稳定,但在3月23日样品中突然显著上升,然后在4月13号的样品中检测到含量下降。进一步通过转录表达共分析,发现茶氨酸合成酶与茶氨酸的含量负相关,而氨基酸转运蛋白CsLHT与茶氨酸含量呈正相关,说明从冬季到春季对茶氨酸含量的变化部分受茶氨基酸转运能力的影响。Ma等[12]研究了春、夏、秋3个不同采收季节对茶叶代谢物的影响,发现采收季节对大部分氨基酸含量影响显著,与以往研究类似,其中春季鲜叶氨基酸含量最高。
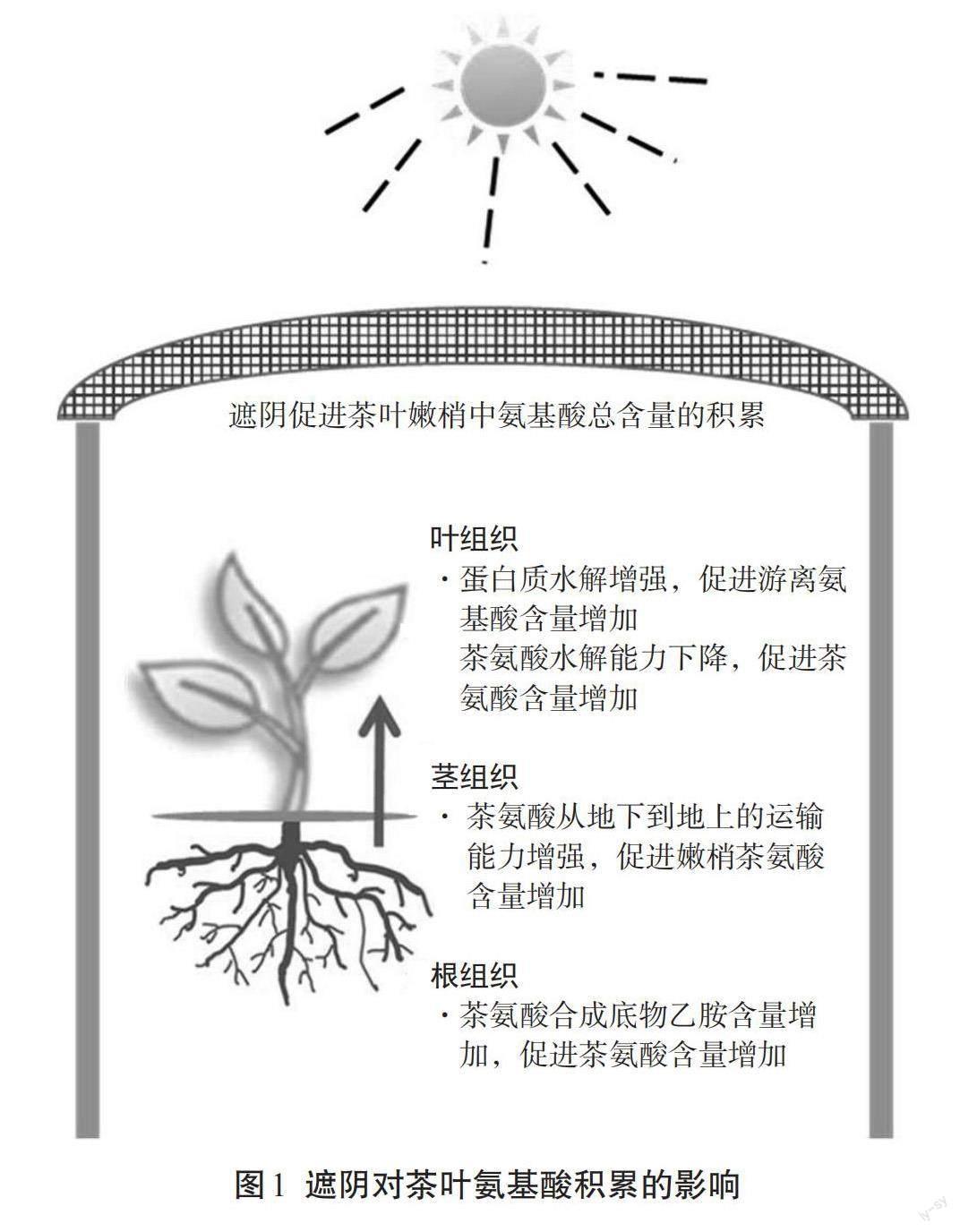
光照强度和温度往往是不同季节中变化幅度较大的环境因子。光环境因子对茶鲜叶氨基酸代谢影响的研究较多,并主要集中于研究遮阴对氨基酸含量的影响。大部分研究发现茶叶经过长时间的遮阴或黑暗处理后,游离氨基酸含量会显著增加[13-14](图1)。Chen等[13]发现在长期黑暗环境诱导下,叶绿体蛋白酶水平升高且叶绿体蛋白含量显著降低。因此,他们推断茶叶经黑暗处理后,叶片内的叶绿体蛋白被叶绿体蛋白酶水解从而生成大量游离氨基酸。Ji等[14]发现随着游离氨基酸的增多,葡萄糖含量大量减少,说明发生了碳饥饿;同样得出长时间黑暗处理后叶绿体内蛋白质发生水解导致游离氨基酸增多的结论。
除了上述蛋白源氨基酸的增加,非蛋白源氨基酸茶氨酸的含量也在遮阴环境下显著增加[15-17](图1)。Fu等[16]对遮阴处理后的茶树组织(叶、茎、根)进行单独分析,发现叶片中的茶氨酸含量并没有显著变化,根组织和嫩茎中茶氨酸含量显著增加。根组织中主要的茶氨酸合成酶基因CsTSⅠ 的表达量却在遮阴后下降,进一步对茶氨酸合成底物谷氨酸和乙胺含量进行分析,发现根中乙胺含量显著增加。因此推测遮阴处理后茶树根中茶氨酸含量增加可能是由于底物乙胺含量增加导致,而嫩茎中茶氨酸含量的增加可能与茶氨酸的转运相关。Yang等[17]对遮阴条件下茶氨酸转运蛋白的表达水平进行了研究,发现在嫩茎中转运蛋白基因CsAAP2/4/5/8的表达水平被遮阴显著诱导,也说明了遮阴可促进茶氨酸从地下组织向地上组织转运的能力。综上,遮阴可以促进茶树根组织中乙胺含量的增加从而使根组织中茶氨酸含量增加,茎组织中茶氨酸含量的增加则与其转运蛋白基因表达增强相关。总体而言,茶树遮阴条件下嫩梢中总游离氨基酸的增多,一方面来自于叶绿体蛋白水解产生的蛋白源氨基酸;另一方面则是减少了叶中茶氨酸的水解,并且加强了来自于根组织中茶氨酸的运输。相对于光因素试验,目前温度对茶鲜叶中氨基酸的合成和代谢方面的研究较少,Li等[18]研究发现持续高温(35 ℃)可显著降低茶氨酸的含量,并与高温下茶氨酸合成酶相关基因CsGS和CsGOGAT的表达水平及酶活水平被显著抑制相关。
3 加工工艺及贮藏对茶叶氨基酸合成代谢及转化的影响
茶叶氨基酸含量在加工过程中的变化主要受两方面作用的影响,一方面是在湿热作用下一些小分子蛋白质或多肽等物质发生局部水解、热解使氨基酸得到积累;另一方面则是氨基酸被氧化、水解、转化,以及与糖、多酚结合形成色、香、味物质而减少。
3.1 厌氧处理
γ-氨基丁酸(GABA)在茶鲜叶中含量非常低,但经过胁迫处理,尤其是在摊放或萎凋前期进行厌氧处理后,其含量会急剧增加。研究发现GABA形成的主要途径来自于谷氨酸的脱羧反应,在厌氧过程中一方面通过激活了谷氨酸脱羧酶酶活CsGAD1,另一方面促进CsGAD2基因成员转录水平,导致了GABA含量的提升[19]。厌氧除了可以影响GABA含量外,也会促进丙氨酸和组氨酸的积累,而谷氨酸、天冬氨酸、瓜氨酸和磷酸乙醇胺的含量则会显著降低。张金玉等[20]研究了不同厌氧时间(0、2、4、8 h)对福鼎大白茶、黄金芽和白叶1号的一芽二叶鲜叶中GABA积累的影响。发现随着厌氧时长增加,3个品种的茶叶游离氨基酸含量均显著增加,且黄金芽和白叶1号的GABA富集效果更好。另外,茶叶组织不同部位厌氧处理后GABA含量的增加也不一样。杨高中等[21]分析了厌氧处理后一芽五叶不同叶位及茎梗的氨基酸含量,发现第一叶和第二叶增幅较大,可达到6倍,而三、四、五叶则增幅较小。
3.2 摊放和萎凋
摊放一般为绿茶、黄茶、黑茶制作工序的第一道工艺,经过摊放后一般氨基酸总量呈上升趋势。张娇等[22]在茶叶摊放2~6 h过程中,发现氨基酸质量分数从1.58%增长至1.73%。许伟等[23]同样发现碧香早春季一芽二叶在摊放阶段,氨基酸含量呈上升趋势。另外,Liu等[24]研究了摊放时不同温度对茶氨酸含量的影响,发现随着摊放温度的升高,茶氨酸含量呈现下降的趋势,并推测高温下谷氨酸合成酶基因CsGOGAT的低表达水平与低茶氨酸含量有一定相关性。
萎凋是白茶、红茶和乌龙茶等茶类制作工序的第一道工序,其主要目的是降低含水量,在萎凋过程中,叶片的质地由脆变软,叶中所含酶类物质的活性增强,产生复杂的化学反应,如淀粉、低分子蛋白质或一些肽类等成分发生分解、转化,生成葡萄糖和氨基酸等化合物,同时多酚类物质也会被不同程度氧化[25]。宋振硕等[25]对10份茶树资源以萎凋失水减重率为跟踪检测点分析主要品质成分含量的变化,发现游離氨基酸的总含量随着萎凋程度的加重而增加,推测与萎凋过程中小分子蛋白质降解相关。陈佳佳等[26]则根据萎凋时间点对萎凋叶中氨基酸及相关酶进行检测和分析,发现游离氨基酸总量在萎凋前期(0~12 h)显著上升,推测主要来源于蛋白质的水解;而在萎凋后期(12~30 h)游离氨基酸总量下降至萎凋前水平,推测氨基酸发生了转化或水解形成了其他物质。对于非蛋白源氨基酸,杨高中等[21]发现经过厌氧处理后富含高GABA的鲜叶在经过萎凋后,GABA含量显著下降,推测在萎凋过程中GABA在转氨酶的作用下转化为琥珀酸半醛,再脱氢后进入三羧酸循环,导致降解。对于茶氨酸,研究发现在萎凋过程中其含量出现明显下降[27]。Cheng等[27]通过加权基因共表达网络分析(WGCNA)发现,茶氨酸水解酶基因CsPDX2.1与茶氨酸的含量呈显著负相关,推测在茶叶萎凋过程中茶氨酸水解能力的增强导致了茶氨酸含量的下降。上述研究说明茶叶萎凋过程中氨基酸含量变化是一个动态变化的过程。
光质对萎凋过程中氨基酸含量的变化也会产生影响。柯茜[28]在萎凋过程中利用不同光质对茶叶进行处理,发现经红光处理后的萎凋叶,其氨基酸含量显著高于自然萎凋,并在萎凋处理前期,发现天冬氨酸合成酶(谷草转氨酶)、谷氨酸合成酶和茶氨酸合成酶的相关基因在红光处理下表达水平上调,而天冬氨酸相关的分解转化酶(精氨琥珀酸裂解酶、天冬氨酸氨基转移酶、天冬氨酸氧化酶)基因和谷氨酸分解酶基因的表达量被红光抑制,从而促进萎凋叶在红光处理下天冬氨酸、谷氨酸和茶氨酸含量的积累。
3.3 发酵
茶叶的发酵过程实质上是茶叶内化合物的氧化及各种化学反应的发生过程。红茶的发酵与普洱熟茶的发酵有所不同,红茶发酵主要通过茶叶内多酚氧化酶来发酵,时间一般只需要几个小时,最长一般不超过12 h。普洱熟茶是在制作毛茶完成之后,再进行人工渥堆发酵,主要通过微生物种群参与发酵,发酵时间相对较长(20~50 d)。
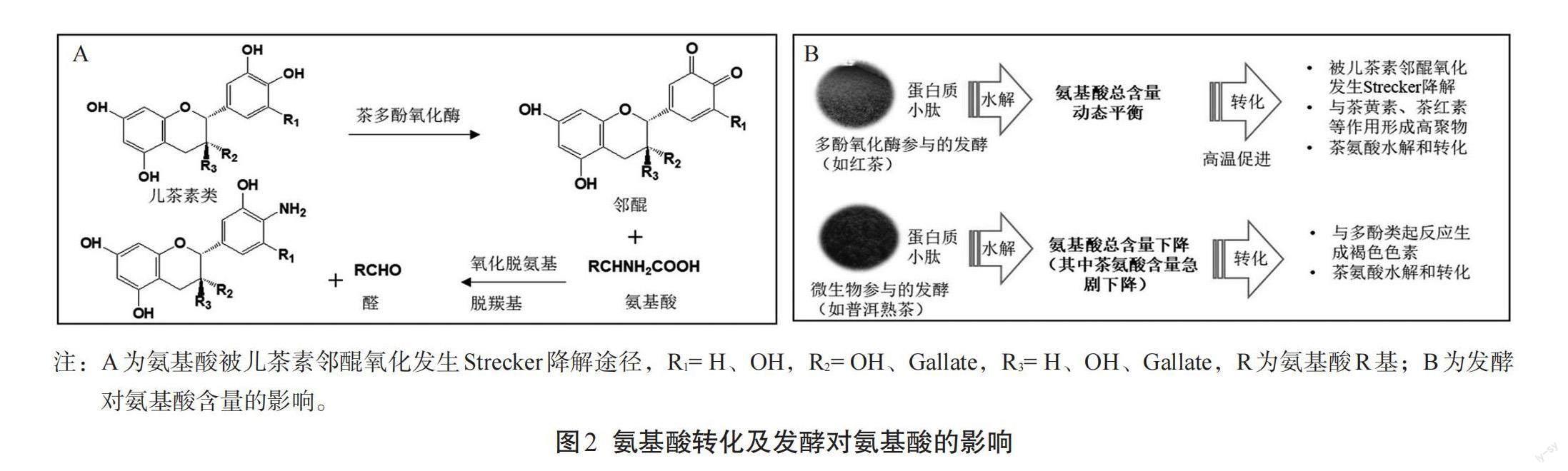
在正常的植物体内,醌和氨基酸存在不同的细胞空间,不易发生反应;而当茶叶在发酵过程中,尤其是红茶发酵过程中,经揉捻后细胞破损,细胞中的酚类物质被多酚氧化酶氧化,由此产生的醌类化合物就有机会与氨基酸发生一些可逆或不可逆的化学反应(图2-A)。如这些醌类物质可以与蛋白质或氨基酸的官能团如酰胺、胺、巯基、吲哚和咪唑等发生取代反应,生成席夫碱、N-醌基衍生物、S-醌基衍生物,及Strecker降解产生的一些醛类化合物[29]。Sanderson等[30]发现部分红茶香气来自于氨基酸的Strecker降解,如甲醛来自于甘氨酸、乙醛来自于丙氨酸、异丁醛来自于缬氨酸、异戊醛来自于亮氨酸、苯乙醛来自于苯丙氨酸、2-甲基丁醛来自于异亮氨酸、甲二磺醛来自于蛋氨酸等。这些醛类有不少对红茶香气呈现具有重要作用,如苯乙醛具有玫瑰花香,已有相关报道其与优质红茶相关。
另外,在发酵过程中氨基酸也能与儿茶素的邻醌结合生成有色化合物,这与红茶汤色形成相关。Tan等[31]对红茶发酵不同阶段进行化合物的变化分析,发现茶氨酸含量显著降低,并伴随着一个茶氨酸衍生物的含量显著增加。因此,推测在红茶发酵过程中茶氨酸可能被糖基化生成了新的茶氨酸葡糖苷化合物。Chen等[32]研究发现蛋白总含量和氨基酸总含量在红茶发酵过程中均下降,并通过加入人工合成的二肽(Z-Glu-Tyr)验证了在红茶发酵过程中蛋白质会被水解成氨基酸,也通过添加同位素标记的苯丙氨酸验证了氨基酸会转化为香气物质。上述这些研究结果表明,发酵期间红茶中氨基酸的含量是氨基酸形成和降解转化之间一个动态变化的结果(图2-B)。
Qu等[33]比较了不同发酵温度对红茶中氨基酸含量的影响,发现31 ℃条件下氨基酸含量比35 ℃高。Salman等[34]研究了乌龙茶制作过程中发酵时间对氨基酸含量的影响,发现5月采摘的茶鲜叶,大部分氨基酸(除了谷氨酸和酪氨酸)含量随着发酵时间延长而增加;而对于7月和9月采摘的茶鲜叶,氨基酸含量与发酵时长则没有显著相关性。
相对于红茶有多酚氧化酶参与的发酵,普洱熟茶的发酵则是微生物参与的发酵,研究显示氨基酸总量呈下降趋势(图2-B)。如梁名志等[35]研究发现,氨基酸在普洱茶发酵的特殊温湿条件下可发生氧化、降解和转化,并与多酚类起反应生成褐色色素,因此在普洱茶渥堆发酵过程中氨基酸总含量呈减少趋势,第五翻混合茶坯中氨基酸含量仅为0.56%。Zhu等[36]研究了普洱茶发酵期间氨基酸组分的变化。他们发现茶叶经过了49 d的发酵后,氨基酸总量显著下降,其中茶氨酸、色氨酸和缬氨酸的含量分别降低了94.58%、20.54%和37.93%,而丙氨酸、精氨酸和亮氨酸的含量显著增加,其余氨基酸的含量则没有显著性变化。另外,国内研究团队还研究了来自茶叶的真菌菌株对普洱茶渥堆发酵的影响,也发现茶叶中大多数氨基酸含量在发酵期间含量显著降低[37-38]。
3.4 杀青和干燥
杀青是绿茶、黄茶、黑茶、乌龙茶和普洱茶的工序之一,其主要目的是通过高温破坏茶鲜叶中的氧化酶活性从而抑制茶多酚等的酶促氧化,同时将部分水分蒸发,使叶组织变软,便于进一步加工,另外还可散发青臭味,促进良好香气的形成。杀青过程中,高温高湿的环境既会促进蛋白质的水解,又会使氨基酸水解和转化。研究显示杀青温度越高,杀青叶氨基酸含量则越低,因此推测温度越高,氨基酸和羰基化合物(如还原糖)的美拉德反应越强烈,消耗了更多的氨基酸。另外,氨基酸在高温下也易发生脱羧、脱氨等降解反应,使氨基酸含量减少。研究发现当茶氨酸被加热到180 ℃时,会产生大量的N-乙基甲酰胺、乙胺、丙胺、2-吡咯烷酮、N-乙基琥珀酰亚胺、1-乙基-3,4-脱氢吡咯烷酮等物质。茶氨酸和葡萄糖一起加热到150 ℃时,则会形成1-乙基-3, 4-脱氢吡咯烷酮及少量吡嗪和呋喃[39]。
Y?lmaz等[40]发现红茶干燥结束时,氨基酸总量由原先的8 799.7 mg/kg 降至7 702.0 mg/kg,推測在干燥阶段,氨基酸含量的降低可能与发生糖-氨基酸相互作用有关。研究者还比较了不同干燥方法对不同茶类氨基酸的影响。Qu等[41]比较了常规热风干燥、微波干燥、远红外干燥、卤素灯干燥和卤素灯-微波组合干燥对红茶氨基酸含量的影响,发现传统热风干燥的茶叶氨基酸含量最高,卤素灯-微波组合干燥法氨基酸含量最低。推测特殊热辐射卤素灯或卤素灯结合微波可能会加速氨基酸和可溶性糖转化为其他化合物(如芳香化合物),或者是使用该方法的干燥时间比较短,使蛋白水解成氨基酸的量比较少,具体原因还有待进一步解析。综上研究,不同种类的茶叶使用不同干燥方法对氨基酸含量的影响均不同。为提升绿茶品质,一般在加工过程中要尽可能保留更多的氨基酸;而对于红茶,氨基酸含量的要求则不是非常高,因此在干燥过程中可以适当促进氨基酸和其他多种化合物的相互转化,不但能增进茶汤滋味,还有利于茶叶香气的形成。
3.5 贮藏
氨基酸在茶叶贮藏过程中会因环境和时间的变化发生进一步降解与转化(图3)。Ning等[42]发现白茶经过长时间储存,氨基酸含量显著下降,如1993年制成的茶叶贮藏20年后氨基酸含量只有1.536 mg/g,而2013新制的茶叶氨基酸含量为14.848 mg/g,接近1993年茶叶的10倍。推测在长时间贮藏期间氨基酸可能被儿茶素邻醌氧化发生Strecker降解,导致其含量下降。另外还发现短期贮藏对氨基酸含量影响不大,如2010年制的茶叶,经过3年贮藏后测得氨基酸含量为10.752 mg/g。Zhu等[43]分析了不同年份的雅安藏茶氨基酸的含量,发现贮藏5年与1年的差异不显著,但到贮藏10年氨基酸含量急剧下降。Dai等[44]发现随着白茶贮藏期的增加,其茶氨酸含量会随之减少,但其与儿茶素类物质生成的产物N-乙基-2-吡咯烷酮取代的黄烷醇类化合物含量则增加。同样,Peng等[45]发现,在乌龙茶贮藏15年后茶氨酸含量只有0.78 mg/g,只占原先茶氨酸含量的10%,其转化产物N-乙基-2-吡咯烷酮取代的黄烷醇类化合物的含量与上述研究类似,呈现增加的趋势。Huang等[46]比较了不同贮藏年份对祁门红茶氨基酸的影响,同样发现在10年后茶氨酸含量会急剧下降。
除了贮藏时间会影响茶叶中氨基酸的积累,不同贮藏条件如温度和湿度等也会影响其含量和组成。Dai等[47]比较了不同温度对黄山毛峰贮藏150 d后氨基酸的影响,发现氨基酸的含量在4 ℃和室温条件下下降程度显著高于-80 ℃和-20 ℃条件,说明低温可以维持茶叶氨基酸的含量。Zhou等[48]比较了贮藏环境对普洱生茶氨基酸含量的影响,发现相对于东莞高温潮湿的环境,低温和干燥的环境有利于保存大部分氨基酸的含量。综上研究,低温和干燥的环境可以降低贮藏期间氨基酸的降解。
4 总结与展望
4.1 采前氨基酸影响因素及积累规律与机制
影响采前茶叶氨基酸含量的主要因素为种质资源和外界环境。在种质资源方面,一般黄化、白化品种茶树的嫩梢氨基酸含量要显著高于绿色叶品种。研究显示茶氨酸在黄化、白化品种中较弱的降解能力,与其高茶氨酸的积累相关。由于茶氨酸可占游离氨基酸总量的50%以上,因此黄化、白化品种的嫩梢氨基酸总量较常规绿色叶品种高。在外界环境影响方面,研究发现由于强烈的光照和高温,导致茶树呈现较低的氮代谢及较强的茶氨酸水解和转化能力,使得夏季嫩梢中氨基酸含量较低。因此,通过遮阴措施来降低光照强度,可促进游离氨基酸的积累,包括蛋白源氨基酸和非蛋白源氨基酸茶氨酸。其中蛋白源氨基酸增多,与遮阴促进了叶绿体蛋白的水解有关;茶氨酸的增加,一方面由于遮阴降低了叶片茶氨酸的水解能力,另一方面则增强了来自根中茶氨酸运输的能力。
4.2 采后氨基酸影响因素及积累规律与机制
采后主要是研究不同的加工技术对叶片氨基酸含量的影响。本文主要分析了摊放、萎凋、发酵、杀青、干燥及贮藏阶段氨基酸含量的变化及其中涉及的相关代谢及转化。
4.2.1 摊放和萎凋阶段
摊放和萎凋阶段,氨基酸总量一般呈上升趋势,主要是由于蛋白质水解形成了游离氨基酸,但茶氨酸含量却在这一阶段呈下降趋势,主要是失水激活了茶氨酸水解酶的表达,使茶氨酸被水解。另外,如果在这个阶段前期对茶鲜叶进行厌氧处理,GABA含量会因为谷氨酸脱羧酶被激活及基因表达量上升而显著增加。
4.2.2 发酵阶段
发酵,主要分多酚氧化酶发酵和微生物发酵。多酚氧化酶发酵过程中氨基酸含量呈现一种动态平衡,取决于蛋白质的继续水解形成氨基酸和氨基酸的转化与降解(主要与儿茶素氧化生成的醌发生的各种反应)两方面的影响;微生物发酵由于时间较长,一般发酵后氨基酸含量,尤其是茶氨酸的含量都呈明显下降趋势。
4.2.3 杀青和干燥阶段
杀青和干燥阶段,高温起主导作用,氨基酸往往会与邻醌发生氧化聚合,或者与羰基化合物(如还原糖)发生美拉德反应。另外,氨基酸在较高温下也易发生脫羧、脱氨等降解反应。
4.2.4 贮藏阶段
长时间贮藏期间氨基酸会继续被儿茶素邻醌氧化发生Strecker降解而减少。温度越高、湿度越大,降解量则越大。另外,茶氨酸含量往往在贮藏10年后会显著降低,研究显示可能与儿茶素类物质生成的产物N-乙基-2-吡咯烷酮取代的黄烷醇类化合物相关。
综上,氨基酸不仅是呈味物质,还在茶叶不同加工与贮藏过程中发生各种酶促或非酶促的化学反应,是茶叶香气及色素的重要前体物质。因此,氨基酸对茶叶芳香、清醇及鲜爽的风味贡献了重要的作用。本文主要总结了近几年来采前和采后茶叶中氨基酸合成代谢及转化的相关研究进展。目前关于茶树品种、环境响应,以及采后加工与贮藏环境对氨基酸尤其是茶氨酸的合成代谢及转运机制已有较多且深入的研究。同时,采后GABA的合成代谢机制也取得了系列进展,但采前及采后不同加工工艺对茶叶中其他氨基酸的代谢调控机制研究尚比较笼统,有待后续进一步深入研究。综合了解大部分氨基酸的合成代谢和转化规律及其背后的调控机制,可为后期精准提高茶叶综合品质提供有效的研究基础。
参考文献
[1] YU Z M, YANG Z Y. Understanding different regulatory mechanisms of proteinaceous and non-proteinaceous amino acid formation in tea (Camellia sinensis) provides new insights into the safe and effective alteration of tea flavor and function[J]. Critical Reviews in Food Science and Nutrition, 2020, 60(5): 844-858.
[2] HUANG R, WANG J Y, YAO M Z, et al. Quantitative trait loci mapping for free amino acid content using an albino population and SNP markers provides insight into the genetic improvement of tea plants[J/OL]. Horticulture Research, 2022, 9: uhab029. https://doi.org/10.1093/hr/uhab029.
[3] LIN S J, CHEN Z P, CHEN T T, et al. Theanine metabolism and transport in tea plants (Camellia sinensis L.): Advances and perspectives[J]. Critical Reviews in Biotechnology, 2022, 43(3): 327-341.
[4] YU P H, HUANG H H, ZHAO X, et al. Dynamic variation of amino acid content during black tea processing: A review[J/OL]. Food Reviews International, 2022: 494. https://doi.org/10.1080/87559129.20
21.2015374.
[5] LIU G F, HAN Z X, FENG L, et al. Metabolic flux redirection and transcriptomic reprogramming in the albino tea cultivar 'Yu-Jin-Xiang' with an emphasis on catechin production[J/OL]. Scientific Reprots, 2017, 7: 45062. https://doi.org/10.1038/srep45062.
[6] CHENG S H, FU X M, LIAO Y Y, et al. Differential accumulation of specialized metabolite L-theanine in green and albino-induced yellow tea (Camellia sinensis) leaves[J]. Food Chemistry, 2019, 276: 93-100.
[7] XU Y X, YANG L, LEI Y S, et al. Integrated transcriptome and amino acid profile analyses reveal novel insights into differential accumulation of theanine in green and yellow tea cultivars[J]. Tree Physiology, 2022, 42(7): 1501-1516.
[8] LIU Z W, WU Z J, LI H, et al. L-Theanine content and related gene expression: Novel insights into theanine biosynthesis and hydrolysis among different tea plant (Camellia sinensis L.) tissues and cultivars[J/OL]. Frontiers in Plant Science, 2017, 8: 498. https://doi.org/10.3389/fpls.2017.00498.
[9] FU X M, CHENG S H, LIAO Y Y, et al. Characterization of L-theanine hydrolase in vitro and subcellular distribution of its specific product ethylamine in tea (Camellia sinensis)[J]. Journal of Agricultural and Food Chemistry, 2020, 68(39): 10842-10851.
[10] FANG K X, XIA Z Q, LI H J, et al. Genome-wide association analysis identified molecular markers associated with important tea flavor-related metabolites[J/OL]. Horticulture Research, 2021, 8(1): 42. https://doi.org/10.1038/s41438-021-00477-3.
[11] LI F, DONG C X, YANG T Y, et al. Seasonal theanine accumulation and related gene expression in the roots and leaf buds of tea plants (Camellia sinensis L.)[J/OL]. Frontiers in Plant Science, 2019, 10: 1397. https://doi.org/10.3389/fpls.2019.01397.
[12] MA B S, WANG J C, ZHOU B X, et al. Impact of harvest season on bioactive compounds, amino acids and in vitro antioxidant capacity of white tea through multivariate statistical analysis[J/OL]. LWT - Food Science and Technology, 2022, 164: 113655. https://doi.org/10.1016/j.lwt.2022.113655.
[13] CHEN Y Y, FU X M, MEI X, et al. Proteolysis of chloroplast proteins is responsible for accumulation of free amino acids in dark-treated tea (Camellia sinensis) leaves[J]. Journal of Proteomics, 2017, 157: 10-17.
[14] JI H G, LEE Y R, LEE M S, et al. Diverse metabolite variations in tea (Camellia sinensis L.) leaves grown under various shade conditions revisited: A metabolomics study[J]. Journal of Agricultural and Food Chemistry, 2018, 66(8): 1889-1897.
[15] YANG Z Y, KOBAYASHI E, KATSUNO T, et al. Characterisation of volatile and non-volatile metabolites in etiolated leaves of tea (Camellia sinensis) plants in the dark[J]. Food Chemistry, 2012, 135(4): 2268-2276.
[16] FU X M, LIAO Y Y, CHENG S H, et al. Nonaqueous fractionation and overexpression of fluorescent-tagged enzymes reveals the subcellular sites of L-theanine biosynthesis in tea[J]. Plant Biotechnology Journal, 2021, 19(1): 98-108.
[17] YANG T Y, XIE Y X, LU X, et al. Shading promoted theanine biosynthesis in the roots and allocation in the shoots of the tea plant (Camellia sinensis L.) cultivar Shuchazao[J]. Journal of Agricultural and Food Chemistry, 2021, 69(16): 4795-4803.
[18] LI X, WEI J P, AHAMMED G J, et al. Brassinosteroids attenuate moderate high temperature-caused decline in tea quality by enhancing theanine biosynthesis in Camellia sinensis L. [J/OL]. Frontiers in Plant Science, 2018, 9: 1016. https://doi.org/10.3389/fpls.2018.01016.
[19] MEI X, CHEN Y Y, ZHANG L Y, et al. Dual mechanisms regulating glutamate decarboxylases and accumulation of gamma-aminobutyric acid in tea (Camellia sinensis) leaves exposed to multiple stresses[J/OL]. Scientific Reports, 2016, 6: 23685. https://doi.org/10.1038/srep23685.
[20] 张金玉, 李美凤, 郜秋艳, 等. 不同厌氧时间对绿茶和红茶加工品质的影响[J]. 茶叶学报, 2021, 62(2): 78-84.
[21] 杨高中, 彭群华, 张悦, 等. 厌氧处理对不同类型茶叶的氨基酸组成及生物活性的影响[J]. 茶叶科学, 2022, 42(2): 222-232.
[22] 张娇, 梁壮仙, 张拓, 等. 黄茶加工中主要品质成分的动态变化[J]. 食品科学, 2019, 40(16): 200-205.
[23] 许伟, 彭影琦, 张拓, 等. 綠茶加工中主要滋味物质动态变化及其对绿茶品质的影响[J]. 食品科学, 2019, 40(11): 36-41.
[24] LIU Z W, LI H, WANG W L, et al. CsGOGAT is important in dynamic changes of theanine content in postharvest tea plant leaves under different temperature and shading spreadings[J]. Journal of Agricultural and Food Chemistry, 2017, 65(44): 9693-9702.
[25] 宋振碩, 王丽丽, 陈键, 等. 茶鲜叶萎凋过程中主要品质成分的动态变化[J]. 茶叶学报, 2016, 57(3): 138-141.
[26] 陈佳佳, 朱陈松, 朱文伟, 等. 白茶萎凋过程中氨基酸类物质代谢分析[J]. 茶叶科学, 2021, 41(4): 471-481.
[27] CHENG H Y, WU W, LIU X F, et al. Transcription factor CsWRKY40 regulates L-theanine hydrolysis by activating the CsPDX2.1 promoter in tea leaves during withering[J/OL]. Horticulture Research, 2022, 9: uhac025. https://doi.org/10.1093/hr/uhac025.
[28] 柯茜. 基于转录组的红茶红光萎凋过程中氨基酸含量变化的机理[D]. 武汉: 华中农业大学, 2016.
[29] BITTNER S. When quinones meet amino acids: Chemical, physical and biological consequences[J]. Amino Acids, 2006, 30(3): 205-224.
[30] SANDERSON G W, GRAHAMM H N. Formation of black tea aroma[J]. Journal of Agricultural and Food Chemistry, 1973, 21(4): 576-585.
[31] TAN J F, DAI W F, LU M L, et al. Study of the dynamic changes in the non-volatile chemical constituents of black tea during fermentation processing by a non-targeted metabolomics approach[J]. Food Research International, 2016, 79: 106-113.
[32] CHEN Y Y, ZENG L T, LIAO Y Y, et al. Enzymatic reaction-related protein degradation and proteinaceous amino acid metabolism during the black tea (Camellia sinensis) manufacturing process[J/OL]. Foods, 2020, 9(1): 66. https://doi.org/10.3390/foods9010066.
[33] QU F F, ZENG W C, TONG X , et al. The new insight into the influence of fermentation temperature on quality and bioactivities of black tea[J/OL]. LWT - Food Science and Technology, 2020, 117: 108646. https://doi.org/10.1016/j.lwt.2020.108646.
[34] SALMAN S, Y?LMAZ C, G?KMEN V, et al. Effects of fermentation time and shooting period on amino acid derivatives and free amino acid profiles of tea[J/OL]. LWT - Food Science and Technology, 2021, 137: 110481. https://doi.org/10.1016/j.lwt.2021.110481.
[35] 梁名志, 夏丽飞, 陈林波, 等. 普洱茶渥堆发酵过程中理化指标的变化研究[J]. 中国农学通报, 2006, (10): 321-325.
[36] ZHU Y C, LUO Y H, WANG P P, et al. Simultaneous determination of free amino acids in Pu-erh tea and their changes during fermentation[J]. Food Chemistry, 2016, 194: 643-649.
[37] MA C Q, LI X H, ZHENG C Q, et al. Comparison of characteristic components in tea-leaves fermented by Aspergillus pallidofulvus PT-3, Aspergillus sesamicola PT-4 and Penicillium manginii PT-5 using LC-MS metabolomics and HPLC analysis[J/OL]. Food Chemistry, 2021, 350: 129228. https://doi.org/10.1016/j.foodchem.2021.129228.
[38] WANG Z C, ZHENG C Q, MA C Q, et al. Comparative analysis of chemical constituents and antioxidant activity in tea-leaves microbial fermentation of seven tea-derived fungi from ripened Pu-erh tea[J/OL]. LWT - Food Science and Technology, 2021, 142: 111006. https://doi.org/10.1016/j.lwt.2021.111006.
[39] HO C T, ZHENG X, LI S M. Tea aroma formation[J]. Food Science and Human Wellness, 2015, 4(1): 9-27.
[40] Y?LMAZ C, ?ZDEMIR F, G?KMEN V. Investigation of free amino acids, bioactive and neuroactive compounds in different types of tea and effect of black tea processing[J/OL]. LWT - Food Science and Technology, 2020, 117: 108655. https://doi.org/10.1016/j.lwt.2020.108655.
[41] QU F F, ZHU X J, AI Z Y, et al. Effect of different drying methods on the sensory quality and chemical components of black tea[J]. LWT - Food Science and Technology, 2019, 99: 112-118.
[42] NING J M, DING D, SONG Y S, et al. Chemical constituents analysis of white tea of different qualities and different storage times[J]. European Food Research and Technology, 2016, 242(12): 2093-2104.
[43] ZHU C L, YANG Z B, HE L, et al. The longer the storage time, the higher the price, the better the quality? A 1H-NMR based metabolomic investigation of aged Ya'an Tibetan tea (Camellia sinensis)[J/OL]. Foods, 2022, 11(19): 2986. https://doi.org/10.3390/foods11192986.
[44] DAI W D, TAN J F, LU M L, et al. Metabolomics investigation reveals that 8-C N-Ethyl-2-pyrrolidinone-substituted flavan-3-ols are potential marker compounds of stored white teas[J]. Journal of Agricultural and Food Chemistry, 2018, 66(27): 7209-7218.
[45] PENG J K, DAI W D, LU M L, et al. New insights into the influences of baking and storage on the nonvolatile compounds in Oolong tea: A nontargeted and targeted metabolomics study[J/OL]. Food Chemistry, 2022, 375: 131872. https://doi.org/10.1016/j.foodchem.2021.131872.
[46] HUANG A, JIANG Z D, TAO M, et al. Targeted and nontargeted metabolomics analysis for determining the effect of storage time on the metabolites and taste quality of keemun black tea[J/OL]. Food Chemistry, 2021, 359: 129950. https://doi.org/ 10.1016/j.foodchem.2021.129950.
[47] DAI Q Y, LIU S T, JIANG Y R, et al. Recommended storage temperature for green tea based on sensory quality[J]. Journal of Food Science and Technology, 2019, 56(9): 4333-4348.
[48] ZHOU B X, MA C Q, WU T T, et al. Classification of raw Pu-erh teas with different storage time based on characteristic compounds and effect of storage environment[J/OL]. LWT - Food Science and Technology, 2020, 133: 109914. https://doi.org/10.1016/j.lwt.2020.109914.
