芋淀粉分支酶(SBE)基因的鉴定、生物信息学及表达分析
2023-08-14董伟清刘莉莉蒋慧萍邱祖杨何芳练
董伟清 刘莉莉 蒋慧萍 邱祖杨 何芳练


摘 要:淀粉分支酶(starch branching enzyme, SBE)在支链淀粉生物合成中发挥关键作用,直接影响淀粉的含量和结构。芋(Colocasia esculenta)是一种主要的块茎类作物,在世界上热带和亚热带地区广泛栽培。目前,SBE 在芋中的研究很少,对SBE 基因在芋中的数量、分子结构特征和表达模式还不清楚。本研究首次对芋SBE 基因进行了全面分析,鉴定了3 个SBE 基因(CeSBE1、CeSBE2 和CeSBE3)。CeSBE1、CeSBE2 和CeSBE3 蛋白氨基酸数量分别为828、845和598,分子质量分别为92 956.71、95 625.13、69 169.16 Da,等电点分别为5.22、5.41 和7.36。系统进化分析显示3个CeSBE 蛋白分别在3 个不同的亚群。基因结构分析显示,CeSBE1、CeSBE2 和CeSBE3 外显子数量分别为16、22、10;保守结构域分析表明,CeSBE1 和CeSBE2 均具有alpha-amylase_C 和alpha-amylase 结构域及7 个motif,而CeSBE3具有alpha-amylase 和CBM_48 结构域及3 个motif。CeSBE 基因启动子区域顺式作用元件分析表明,共预测到55 个顺式作用元件,其中29 个具有功能注释,涉及光响应、激素响应、植物生长发育及环境压力等相关元件。在不同组织中,3 个CeSBE 基因均能在所有组织中表达,其中CeSBE2 在球茎和叶片显著表达(P<0.05);在球茎不同发育阶段中,CeSBE2 在所有的发育阶段均有较高的表达量,呈现先升高后降低的表达趋势,在球茎发育120 d 的表达量达到峰值。
球茎不同发育阶段总淀粉和支链淀粉含量增加与CeSBE2 表达量趋势一致,说明CeSBE2 可能是芋支链淀粉生物合成的关键基因。本研究结果可为芋的产量、品质和营养性状的遗传改良提供基础。
关键词:芋;淀粉分支酶;生物信息学分析;表达分析
中图分类号:S632.3 文献标识码:A
淀粉是高等植物主要的储存多糖,由直链淀粉和支链淀粉构成,是人们日常饮食中碳水化合物的主要来源[1-2]。淀粉通常存在于谷类作物和块茎类作物中, 如水稻( Oryza sativa ) 、小麦(Triticum aestivum)、玉米(Zea mays)、马铃薯(Solanum tuberosum)、木薯(Manihot esculenta)和芋(Colocasia esculenta)等[3-8]。支链淀粉是淀粉的主要组分,占总淀粉的75%~85%[9]。支链淀粉生物合成涉及的酶有淀粉合成酶(starch synthase,SS)、淀粉分支酶(starch branching enzyme,SBE)和淀粉脱分支酶(starch debranching enzyme,DBE)[10],其中SBE 在淀粉含量、结构以及物理特性方面起着关键作用[1, 11]。SBE 属于α-淀粉酶家族,具有α-淀粉酶催化结构域(A 结构域)、氨基N-末端和羧基C-末端结构域,主要分类为3种类型(SBE1、SBE2 和SBE3),其中SBE1 和SBE2 的功能研究得较为清楚[11]。SBE1 将直链淀粉浓缩成长的葡聚糖链,SBE2 主要利用短葡聚糖链为底物合成支链淀粉,而SBE3 的功能仍然不明确[12]。
许多SBE 基因表达和功能研究揭示了其在支链淀粉生物合成和植物生长发育中的作用。例如,板栗(Castanea mollissima)SBE 基因在果实发育过程中表达量显著增加,与支链淀粉的合成和果实发育密切相关[13] ; 在缺乏SBEIIa 的玉米(Z. mays)中,叶片淀粉的分支急剧减少,叶片表现出严重的类似衰老的表型[14] ; 在水稻(O. sativa)中,通过抑制SBEI 和SBEIIb 基因的表达,降低了支链淀粉含量,将直链淀粉含量从25%提高到约60%,但同时由于淀粉结构的改变,抑制了水稻幼苗的生长[9, 15];在木薯(M. esculenta)中,通过抑制SBE1 和SBE2 的表达,降低了支链淀粉含量,增加了直链淀粉和抗性淀粉的含量[16-17]。
芋是一种主要的块茎类作物,在热带和亚热带地区广泛栽培[18]。淀粉是芋球茎的主要碳水化合物,占干物质含量的70%~80%[19],而支链淀粉又是总淀粉的主要组成部分[8, 20]。芋支链淀粉的生物合成和积累对芋球茎的产量、品质以及营养价值具有重要的意义。因此,有必要在全基因组范围内鉴定与支链淀粉生物合成有关的关键候选基因,以促进芋产量的提高和品质改良。本研究从芋基因组中鉴定了3 个SBE 基因,对其分子特征、系统发育关系、保守结构、基因结构、顺式作用元件和时空表达模式进行分析,为芋的产量、品质和营养性状的遗传改良提供基础。
1 材料与方法
1.1 材料
供试芋材料为芋新品种荔浦芋1 号,该品种为魁芋类型(槟榔芋),以母芋为主要食用器官,具有产量高、品质优和耐储藏等特点。选择播种90 d 的叶片、叶柄、球茎和根用于CeSBE 基因的空间表达分析;选择播种30 d(S1)、60 d(S2)、90 d(S3)、120 d(S4)、150 d(S5)、180 d(S6)、210 d(S7)、240 d(S8)的球茎用于CeSBE 基因的时序性表达分析。所有样品均设置3 次生物学重复,液氮速冻后于–80 ℃冰箱保存备用。
1.2 方法
1.2.1 CeSBE 蛋白的鉴定和系统发育分析 分别从TAIR 数据库(http://www.arabidopsis.org)和RGAP 数据库(http://rice.plantbiology.msu.edu)下载拟南芥和水稻的SBE 家族蛋白序列,使用HMMER 3.0 软件以拟南芥和水稻SBE 家族蛋白序列为模板构建隐马尔可夫模型。通过建立的模型搜寻芋参考基因组(Niue2, https://db.cngb.org/search/project/PRJNA328799/)的所有蛋白序列,找出潜在的CeSBE 蛋白。此外,使用BLASTp软件(版本:2.10.1)将拟南芥和水稻SBE 蛋白序列与芋所有的蛋白序列进行比对,E-value 设为1E-20。将上述获得的CeSBE 蛋白序列进行合并去冗余,去冗余后的蛋白序列作为候选的CeSBE家族蛋白序列。使用NCBI 保守结构域数据库(CDD)(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)和Pfam 數据库(https://pfam.xfam.org/)进一步验证候选CeSBE 蛋白序列的保守结构域。使用mafft 软件(版本:v7.427)对来自芋(C. esculenta)、拟南芥(Arabidopsis thaliana)、水稻(O. sativa)、马铃薯(S. tuberosum)、玉米(Z. mays)、小麦(T. aestivum)、番茄(Solanumlycopersicum)和葡萄(Vitis vinifera)的SBE 蛋白进行多序列比对,然后利用MEGA 7 软件的邻接法构建系统发育树(Bootstrap 值为1000)。
1.2.2 CeSBE 蛋白特性和基因结构分析 使用ExPASy-ProtParam 软件( http://web.expasy.org/protparam/)预测CeSBE 蛋白的理化性质;使用MEME 5.4.1 软件(https://meme-suite.org/meme/tools/meme)预测CeSBE 蛋白的保守motif,然后使用InterProScan 数据库(http://www.ebi.ac.uk/interpro/)对预测的motif 进行功能注释;使用NCBI 数据库(https://www.ncbi.nlm.nih.gov/)和Pfam 数据库(http://pfam.xfam.org/)预测CeSBE蛋白的保守结构域; 使用GSDS 2.0 软件(http://gsds.gao-lab.org/)预测CeSBE 基因的结构特征; 使用PlantCARE 数据库( http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对CeSBE 基因上游2 kb 序列预测启动子区域的顺式作用元件。
1.2.3 CeSBE 基因的转录组分析 对CeSBE 基因在不同组织、球茎不同发育阶段的转录表达情况进行分析。课题组前期对芋发育90 d 的叶片、叶柄、球茎和根开展了转录组测序,从转录组数据中提取CeSBE 基因的FPKM 值,分析CeSBE 基因在不同组织的表达差异;同时,在前期研究中,课题组还对球茎发育30、60、90、120、150、240 d的球茎组织开展了转录组研究[8],从球茎发育的转录组数据中提取CeSBE 基因的FPKM 值,分析CeSBE 基因在球茎不同发育阶段的表达差异。利用CeSBE 基因的FPKM 值,在百迈克云平台( https://international.biocloud.net/zh/software/tools/detail/small/305)绘制CeSBE 基因在不同组织和球茎不同发育阶段的聚类热图。
1.2.4 CeSBE 基因的荧光定量RT-PCR 分析 使用植物多糖多酚总RNA 提取试剂盒[天根生化科技(北京)有限公司,DP441]提取所有样品的总RNA,使用 TIANScriptⅡcDNA 第一链合成试剂盒[天根生化科技(北京)有限公司,KR107]将提取的总RNA 反转录成cDNA 第一链。以不同组织和球茎不同发育阶段的cDNA 为模板,使用AnalytikJena qTOWERE2.2 荧光定量PCR 仪进行荧光定量RT-PCR 试验。根据芋CeSBE 基因序列信息,利用Primer 5.0 软件设计荧光定量RT-PCR引物(表1),以芋Actin 基因为内参基因[21]。使用2×ChamQUniversal SYBR qPCR Master Mix 试剂盒(南京诺唯赞生物科技有限公司,Q711-02)进行荧光定量RT-PCR 试验。反应体系为:SYBRGreen Master Mix 10 μL,上下游引物(10 μmol/L)各0.4 μL,模板cDNA 1.0 μL,双蒸水补足20.0 μL。
荧光定量RT-PCR 反应程序如下:95 ℃预变性3 min,每循环95 ℃ 15 s,57 ℃ 15 s,72 ℃ 20 s,45 个循环,每个样品3 次重复。采用2-ΔΔCT 法计算基因的相对表达量[22]。
2 结果与分析
2.1 CeSBE 蛋白的鉴定和系统发育分析
在芋基因组(Niu2)中共鉴定到3 个SBE 基因,分别命名为CeSBE1(Taro_034516)、CeSBE2(Taro_009230)和CeSBE3(Taro_053106)。
CeSBE1、CeSBE2 和CeSBE3 蛋白氨基酸数量分别为828、845、598,分子质量分别为92 956.71
Da、95 625.13 Da、69 169.16 Da,等电点分别为5.22、5.41 和7.36。CeSBE 蛋白在氨基酸数量、分子质量和等电点的差异可能反映其在各种生物过程中的功能差异。
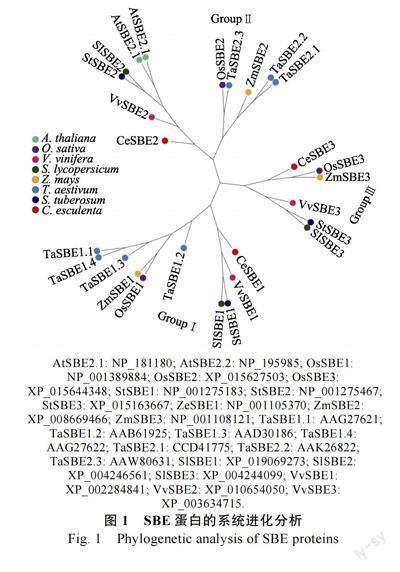
为了研究芋CeSBE 蛋白的进化关系,将27个SBE 蛋白进行系统进化分析,包括3 个芋CeSBE、2 个拟南芥AtSBE、3 個水稻OsSBE、3个马铃薯StSBE、3 个玉米ZmSBE、7 个小麦TaSBE、3 个番茄SlSBE 和3 个葡萄VvSBE。系统进化分析显示(图1),27 个SBE 蛋白被分为3 个群组(Group I、Group II 和Group III)。GroupI 包含10 个SBE 蛋白,包括芋CeSBE1、水稻OsSBE1、马铃薯StSBE1、玉米ZmSBE1、小麦TaSBE1.1/1.2/1.3/1.4 、番茄SlSBE1 和葡萄VvSBE1;Group II 包含11 个SBE 蛋白,包括芋CeSBE2、水稻OsSBE2、拟南芥AtSBE2.1/2.2、马铃薯StSBE2 、玉米ZmSBE2 、小麦TaSBE2.1/2.2/2.3 、番茄SlSBE2 和1 个葡萄VvSBE2;Group III 包含6 个SBE 蛋白,包括芋CeSBE3、水稻OsSBE3、马铃薯StSBE3、玉米ZmSBE3、番茄SlSBE3 和葡萄VvSBE3。
在芋3 个CeSBE 中,CeSBE1、CeSBE2 和CeSBE3 分别归属于Group I、Group II 和Group III(图1)。在Group I 中,CeSBE1 与番茄SlSBE、马铃薯StSBE 和葡萄VvSBE 形成的分支聚在一起; 在Group II 中, CeSBE2 处于拟南芥AtSBE2.1/2.2、马铃薯StSBE2、番茄SlSBE2 和葡萄VvSBE2 形成分支的根部;在Group III 中,CeSBE3 与水稻OsSBE3 和ZmSBE3 形成的分支聚在一起。
2.2 芋CeSBE 基因结构和保守motif 分析
外显子-内含子结构特征分析进一步支持了CeSBE 基因的进化特征(图2A)。CeSBE1、CeSBE2和CeSBE3 外显子数量分别为16、22、10,说明3 个CeSBE 基因在进化和功能上发生了差异。为了进一步探究CeSBE 蛋白结构多样性和预测其功能,使用MEME 5.4.1 软件在3 个CeSBE 蛋白中鉴定了7 个保守的motif,并在InterPro 数据库中进行了注释,所有motif 均注释为1,4-α-葡聚糖分支酶( 1,4-alpha-glucan-branching enzyme,IPR037439)(圖2B、表2)。CeSBE1 和CeSBE2均包含所有的7 个motif,而CeSBE3 只有3 个motif(motif1、motif2、motif3)(图2B)。使用NCBI 数据库和Pfam 数据库对芋CeSBE 的保守结构域进行预测,结果显示CeSBE1 包含alphaamylase_C 和alpha-amylase 结构域,CeSBE2 包含alpha-amylase_C、alpha-amylase 和CBM_48 结构域,CeSBE3 包含alpha-amylase 和CBM_48 结构域(表3)。3 个CeSBE 在外显子数量、保守motif 和保守结构域的差异,可能反映了3 个CeSBE 之间不同的进化历史和功能差异。
2.3 芋CeSBE 基因启动子区域顺式作用元件分析
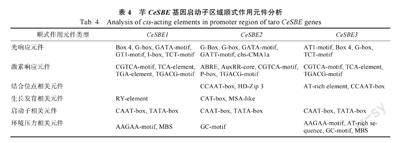
提取3 个芋CeSBE 基因CDS 上游2 kb 序列,使用PlantCARE 数据库预测该区域的顺式作用元件,共预测到55 个顺式作用元件,其中29 个具有功能注释。将具有功能注释的顺式作用元件分为6 类,包括响应光、激素的元件以及与环境压力、结合位点、生长发育和启动子相关的元件(表4)。响应光的顺式作用元件有10 个,其中G-box是3 个CeSBE 基因共有的,其他如Box 4、GT1-motif、GATA-motif 等在其中的1 个或2 个基因出现。响应激素的顺式作用元件有7 个,其中AuxRR-core 和TGA-element 为生长素响应元件,TGACG-motif 和CGTCA-motif 为茉莉酸甲酯(MeJA)响应元件,ABRE 为脱落酸(ABA)响应元件,P-box 为赤霉素(GA)响应元件,TCA-element 为水杨酸响应元件。其他类型顺式作用元件如与分生组织表达有关的顺式调节元件CAT-box,参与干旱诱导的MYB 响应元件MBS,转录起始位点上游30 bp 附近的核心启动子元件 TATA-box 等。说明芋CeSBE 基因可以响应各种环境变化并协调芋的生长发育,但3 个CeSBE基因在不同类型的响应元件中存在差异。
2.4 芋CeSBE 基因表达模式分析
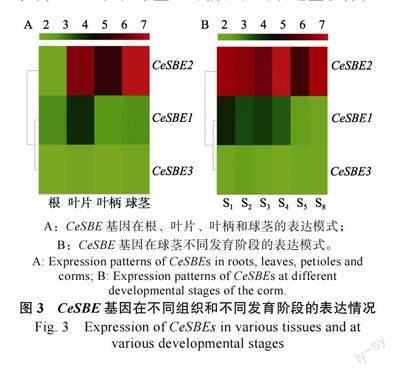
为了研究CeSBE 基因在芋生长发育中的功能,根据课题组前期对芋球茎淀粉的积累规律,对植株发育90 d 的不同组织(叶片、叶柄、球茎和根)和球茎不同发育阶段的样品进行了转录组测序,从转录组数据中提取3 个CeSBE 基因的FPKM 值,对CeSBE 基因在不同组织和球茎不同发育阶段的表达模式进行了分析(图3)。在不同组织中,CeSBE2 在球茎的表达量最高,FPKM值为203.27,其次为叶片和叶柄,FPKM 值分别为108.72 和46.48,根的表达量最低,FPKM 值仅为3.95;CeSBE1 在叶片的表达量较高,FPKM值为17.86,在球茎、叶柄和根的表达量较低,FPKM 值均小于10;CeSBE3 在所有的组织的表达量均较低,FPKM 值均小于5(图3A)。在球茎不同发育阶段中,CeSBE2 在所有发育阶段均有较高的表达量,其中表达量最高的为S4,FPKM值为128.71;CeSBE1 在球茎发育前期(S1~S3)的表达量相对略高,FPKM 值大于9,发育后期(S4~S8)表达量较低,FPKM 值小于8;CeSBE3的表达量在所有发育阶段均较低(FPKM 值小于4)(图3B)。CeSBE2 在球茎中表达量最高,且在球茎的所有发育阶段均表现持续的高表达,说明其可能在芋球茎发育和淀粉积累中起关键作用。
2.5 芋CeSBE 基因的qRT-PCR 分析
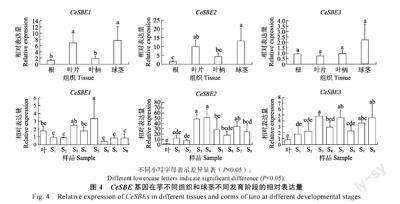
转录组测序结果显示,3 个CesBE 基因在芋不同组织和球茎不同发育阶段表现出不同表达模式,尤其是CeSBE2 在球茎中的表达量最高,且在球茎不同发育阶段持续高表达(图3)。因此,本研究使用荧光定量RT-PCR 进一步验证转录组数据的准确性(图4)。3 个CeSBE 基因在不同组织中显示出不同水平的组织特异性表达。
CeSBE2 在球茎和叶片显著高表达(P<0.05),CeSBE1 与CeSBE2 类似,在球茎和叶片显著表达(P<0.05),但整体表达量较低,CeSBE3 在所有的组织中表达量均较低。在球茎不同发育阶段中,以叶片的表达量为对照,CeSBE2 在球茎发育的所有阶段均有较高的表达量,表达量呈现先升高后降低的趋势;CeSBE3 与CeSBE2 类似,在所有发育阶段的表达量均高于叶片,但整体表达量均较低;CeSBE1 在球茎的S3~S5 阶段的表达量略高于其他发育阶段,但整体表达量均较低。荧光定量RT-PCR 与转录组测序分析保持一致的结果。
2.6 芋球茎发育过程中CeSBE 基因的表达与淀粉含量变化分析
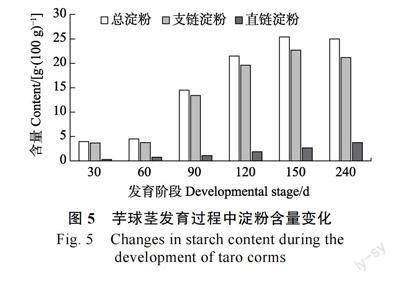
在前期研究中,本课题组对芋球茎发育过程中总淀粉、支链淀粉和直链淀粉的含量进行了测定[8]。总淀粉含量在球茎发育前期较低, 从3.95 g/100 g(30 d)增加到4.51 g/100 g(60 d),在球茎发育中期, 淀粉含量迅速增加, 从14.5 g/100 g(90 d)增加到25.4 g/100 g(150 d);直链淀粉含量在球茎发育过程中缓慢增加,从0.28 g/100 g(30 d)增加到3.76 g/100 g(240 d);支链淀粉含量变化与总淀粉含量变化趋势一致,在球茎发育前期较低,从3.67 g/100 g(30 d)增加到3.77 g/100 g(60 d),在球茎发育中期,支链淀粉含量也迅速增加,从13.4 g/100 g(90 d)增加到22.7 g/100 g(150 d)(图5)。支链淀粉在球茎各发育阶段占总淀粉含量的83.59%~92.91%,说明芋球茎总淀粉含量的增加主要由支链淀粉含量增加引起。在球茎发育过程中,CeSBE2 的表达量远高于CeSBE1 和CeSBE3,因此CeSBE2 可能是支链淀粉合成的关键基因。在球茎发育前期,CeSBE2 表达量较低,此时球茎支链淀粉和总淀粉含量维持在较低水平;在球茎发育中期,CeSBE2 表达量迅速升高且在120 d 达到峰值,此时支链淀粉和总淀粉含量迅速增加;在球茎发育后期,CeSBE2 表达量略有下降,支链淀粉和总淀粉含量缓慢增加。CeSBE2 表达量变化与球茎支链淀粉和总淀粉含量增加趋势一致。
3 讨论
尽管芋也是高淀粉作物,但与其他高淀粉类作物如水稻(O. sativa)、马铃薯(S. tuberosum)和木薯(M. esculenta)等的研究相比,芋的研究相对较少,特别是在支链淀粉生物合成和球茎发育方面[9, 17, 23]。SBE 是支链淀粉生物合成的关键酶,影响淀粉的产量和结构,同时还在植物生长发育中发挥作用[1, 11]。在本研究中,从芋基因组中鉴定了3 个CeSBE 基因,这与水稻(O. sativa)、玉米(Z. mays)、高粱(Sorghum bicolor)、大麥(Hordeum vulgare)和马铃薯(S. tuberosum)等作物的SBE 基因数量一致[1, 12]。但同时也发现,与其他一些作物的SBE 基因数量存在差异,如拟南芥(A. thaliana)有2 个SBE 基因,木薯( M. esculenta ) 有6 个SBE 基因, 小麦(T. aestivum)有7 个SBE 基因,香蕉(Musa acuminata)在不同的基因组中SBE 基因也存在差异,A 基因组有7 个SBE 基因,B 基因组有6 个SBE 基因[1, 12, 24]。根据系统发育分析,3 个CeSBE蛋白分别在不同的群组,这与大多数谷类作物和园艺作物的SBE 分类一致[11]。系统发育分析得到了基因结构和保守结构域分析的进一步支持。外显子-内含子结构特征分析显示3 个CeSBE 基因的外显子数量存在差异,分别为16、22、10,与在香蕉(M. acuminata)上报道的一致[12]。在保守结构域上,CeSBE1 和CeSBE2 蛋白均包含alpha-amylase_C 和alpha-amylase 两个结构域,CeSBE3 只有alpha-amylase 结构域,这与之前在其他作物上报道的一致,均为SBE1 和SBE2 的结构域特征高度保守,而SBE3 缺乏部分保守结构域[11];在保守motif 上,CeSBE1 和CeSBE2 均包含7 个motif,而CeSBE3 只有3 个motif,与在木薯(M. esculenta)上报道的一致[24]。
淀粉代谢受到植物内部时钟和外部昼夜变化的严格调节,SBE 基因的转录表达也呈现昼夜波动[25-26]。在本研究中共有10 个顺式作用元件为光响应元件,与在苹果(Malus domestica)、菠菜(Spinacia oleracea)、马铃薯(S. tuberosum)和拟南芥(A. thaliana)等作物上报道的一致[11]。除了光响应元件外,还有7 个激素响应元件,包括生长素、MeJA、ABA、GA 和水杨酸等,与在木薯上报道的一致[24]。CeSBE 基因上游包含的顺式作用元件说明该基因受环境和激素的调控。
已经有许多作物的研究表明,SBE 基因在贮藏器官的发育过程中发挥关键作用。例如,甘薯(Ipomoea batatas)和板栗(C. mollissima)SBE基因在贮藏器官发育过程中表达量持续增加,直至贮藏器官发育成熟[13, 27]。在豌豆(Pisum sativum)中,SBEII 的缺失导致种子出现皱褶表型,淀粉生物合成减少50%[28]。在本研究中,只有CesBE2 在球茎不同发育阶段均有较高的表达水平,说明CeSBE2 可能在球茎发育过程中发挥关键作用,支链淀粉与总淀粉在球茎不同发育阶段的比例也印证了CeSBE2 的作用,与在苹果(M.domestica)和香蕉(M. acuminata)上报道的一致[12, 29]。鉴于SBE 基因在支链淀粉生物合成中的关键作用,许多作物通过抑制SBE 基因的表达来实现高直链淀粉新种质的创制,目前已经在水稻(O. sativa)、玉米(Z. mays)、木薯(M. esculenta)及小麦(T. aestivum)等作物上获得成功,创制了高直链淀粉的种质[9, 16-17, 30-31]。本研究结果为芋品种改良和高直链淀粉新种质的创制提供了很好的基础和思路。
参考文献
[1] TETLOW I J, EMES M J. A review of starch-branching enzymes and their role in amylopectin biosynthesis[J]. IUBMB Life, 2014, 66(8): 546-558.
[2] BAYSAL C, HE W, DRAPAL M, VILLORBINA G,MEDINA V, CAPELL T, KHUSH G S, ZHU C, FRASER P D, CHRISTOU P. Inactivation of rice starch branching enzyme IIb triggers broad and unexpected changes in metabolism by transcriptional reprogramming[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(42): 26503-26512.
[3] CAI Y, LI S, JIAO G, SHENG Z, WU Y, SHAO G, XIE L, PENG C, XU J, TANG S, WEI X, HU P. OsPK2 encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, compound granule formation and grain filling[J].Plant Biotechnology Journal, 2018, 16(11): 1878-1891.
[4] WANG Z, MA S, SUN B, WANG F, HUANG J, WANG X,BAO Q. Effects of thermal properties and behavior of wheat starch and gluten on their interaction: a review[J]. International Journal of Biological Macromolecules, 2021, 177:474-484.
[5] ZHONG Y, LI Y, QU J, ZHANG X, SEYTAHMETOVNA S A, BLENNOW A, GUO D. Structural features of five types of maize starch granule subgroups sorted by flow cytometry[J]. Food Chemistry, 2021, 356: 129657.
[6] TIESSEN A, HENDRIKS J H, STITT M, BRANSCHEID A,GIBON Y, FARRÉ E M, GEIGENBERGER P. Starch synthesis in potato tubers is regulated by post-translational redox modification of ADP-glucose pyrophosphorylase: a novel regulatory mechanism linking starch synthesis to the sucrose supply[J]. Plant Cell, 2002, 14(9): 2191-2213.
[7] DONG M Y, FAN X W, LI Y Z. Cassava AGPase genes and their encoded proteins are different from those of other plants[J]. Planta, 2019, 250(5): 1621-1635.
[8] DONG W Q, HE F L, JIANG H P, LIU L L, QIU Z Y. Comparative transcriptome sequencing of taro corm development with a focus on the starch and sucrose metabolism pathway[J]. Frontiers in Genetics, 2021, 12: 771081.
[9] WANG J, HU P, LIN L, CHEN Z, LIU Q, WEI C. Gradually decreasing starch branching enzyme expression is responsible for the formation of heterogeneous starch granules[J]. Plant Physiology, 2018, 176(1): 582-595.
[10] ZHU J, YU W, ZHANG C, ZHU Y, XU J, LI E, GILBERT R G, LIU Q. New insights into amylose and amylopectin biosynthesis in rice endosperm[J]. Carbohydrate Polymers,2020, 230: 115656-115662.
[11] YU J, WANG K, BECKLES D M. Starch branching enzymes as putative determinants of postharvest quality in horticultural crops[J]. BMC Plant Biology, 2021, 21(1): 479-494.
[12] MIAO H, SUN P, LIU Q, LIU J, JIA C, ZHAO D, XU B,JIN Z. Molecular identification of the key starch branching enzyme-encoding gene SBE2.3 and its interacting transcription factors in banana fruits[J]. Horticulture Research, 2020,7(1): 1354-1368.
[13] CHEN L, LU D, WANG T, LI Z, ZHAO Y, JIANG Y,ZHANG Q, CAO Q, FANG K, XING Y, QIN L. Identification and expression analysis of starch branching enzymes involved in starch synthesis during the development of chestnut (Castanea mollissima Blume) cotyledons[J]. PLoS One, 2017, 12(5): e0177792.
[14] YANDEAU-NELSON M D, LAURENS L, SHI Z, XIA H,SMITH A M, GUILTINAN M J. Starch-branching enzyme IIa is required for proper diurnal cycling of starch in leaves of maize[J]. Plant Physiology, 2011, 156(2): 479-490.
[15] PAN T, LIN L, WANG J, LIU Q, WEI C. Long branch-chains of amylopectin with B-type crystallinity in rice seed with inhibition of starch branching enzyme I and IIb resist in situ degradation and inhibit plant growth during seedling development: degradation of rice starch with inhibition of SBEI/IIb during seedling development[J]. BMC Plant Biology, 2018, 18(1): 9-19.
[16] UTSUMI Y, UTSUMI C, TANAKA M, TAKAHASHI S,OKAMOTO Y, ONO M, NAKAMURA Y, SEKI M. Suppressed expression of starch branching enzyme 1 and 2 increases resistant starch and amylose content and modifies amylopectin structure in cassava[J]. Plant Molecular Biology,2022, 108(4/5): 413-427.
[17] LUO S, MA Q, ZHONG Y, JING J, WEI Z, ZHOU W, LU X, TIAN Y, ZHANG P. Editing of the starch branching enzyme gene SBE2 generates high-amylose storage roots in cassava[J]. Plant Molecular Biology, 2022, 108(4/5): 429-442.
[18] LU T J, LIN J H, CHEN J C, CHANG Y H. Characteristics of taro (Colocasia esculenta) starches planted in different seasons and their relations to the molecular structure of starch[J]. Journal of Agricultural and Food Chemistry, 2008,56(6): 2208-2215.
[19] NAGAR C K, DASH S K, RAYAGURU K, PAL U S,NEDUNCHEZHIYAN M. Isolation, characterization, modification and uses of taro starch: a review[J]. International Journal of Biological Macromolecules, 2021, 192: 574-589.
[20] 顧绘, 陈赛男, 李良俊, 程立宝. 芋淀粉分支酶SBE 基因的克隆与表达分析[J]. 园艺学报, 2016, 43(10): 2049-2058.
[21] 王立, 殷剑美, 韩晓勇, 张培通, 郭文琦, 李春宏. 芋淀粉合成酶AGPase 基因的克隆及表达分析[J]. 园艺学报,2016, 43(6): 1117-1125.
[22] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method[J]. Methods, 2001, 25(4): 402-408.
[23] TAKEUCHI A, OHNUMA M, TERAMURA H, ASANO K, NODA T, KUSANO H, TAMURA K, SHIMADA H. Creation of a potato mutant lacking the starch branching enzyme gene StSBE3 that was generated by genome editing using the CRISPR/dMac3-Cas9 system[J]. Plant Biotechnology (Tokyo),2021, 38(3): 345-353.
[24] PEI J, WANG H, XIA Z, LIU C, CHEN X, MA P, LU C,WANG W. Phylogeny and expression pattern of starch branching enzyme family genes in cassava (Manihot esculenta Crantz) under diverse environments[J]. Molecular and Cellular Biochemistry, 2015, 406(1/2): 273-284.
[25] BAGUMA Y, SUN C, AHLANDSBERG S, MUTISYA J, PALMQVIST S, RUBAIHAYO P R, MAGAMBO M J,EGWANG T G, LARSSON H, JANSSON C. Expression patterns of the gene encoding starch branching enzyme II in the storage roots of cassava (Manihot esculenta Crantz)[J].Plant Science, 2003, 164(5): 833-839.
[26] GRAF A, SMITH A M. Starch and the clock: the dark side of plant productivity[J]. Trends in Plant Science, 2011, 16(3):169-175.
[27] HAMADA T, KIM S H, SHIMADA T. Starch-branching enzyme I gene (IbSBEI) from sweet potato (Ipomoea batatas); encoding starch branching enzyme I (SBEI) in apple (Malus× domestica, Rosaceae) and its phylogenetic relationship to Sbe genes from other angiosperms[J]. Molecular Phylogenetics and Evolution, 2007, 43(3): 852-863.
[28] BHATTACHARYYA M K, SMITH A M, ELLIS T H,HEDLEY C, MARTIN C. The wrinkled-seed character of pea described by Mendel is caused by a transposon-like insertion in a gene encoding starch-branching enzyme[J]. Cell, 1990, 60(1): 115-122.
[29] HAN Y, GASIC K, SUN F, XU M, KORBAN S S. A gene molecular cloning and expression analysis[J]. Biotechnology Letters, 2006, 28(16): 1255-1261.
[30] ZHAO Y, LI N, LI B, LI Z, XIE G, ZHANG J. Reduced expression of starch branching enzyme IIa and IIb in maize endosperm by RNAi constructs greatly increases the amylose content in kernel with nearly normal morphology[J]. Planta,2015, 241(2): 449-461.
[31] LI J, JIAO G, SUN Y, CHEN J, ZHONG Y, YAN L, JIANG D, MA Y, XIA L. Modification of starch composition, structure and properties through editing of TaSBEIIa in both winter and spring wheat varieties by CRISPR/Cas9[J]. Plant Biotechnology Journal, 2021, 19(5): 937-951.
