树鼩形觉剥夺性弱视模型的视网膜形态变化△
2023-07-14赵秋语
赵秋语 陈 黎 胡 敏
弱视是儿童视力下降最常见的原因,常表现为单眼或双眼最佳矫正视力下降。弱视在全球范围内的患病率为1%~2%[1],部分地区弱视发病率已接近5%[2]。弱视是由儿童时期的视网膜输入异常引起的,其潜在的神经缺陷主要存在于中枢神经系统,慢性异常视网膜输入会导致中枢结构和功能的发育异常[3-4]。但在弱视形成过程中,视网膜和视神经的结构是否发生改变这一观点是存在争议的[5-8]。目前弱视的定义仍在强调眼部的临床检查无可见器质性病变,但已有研究报告了黄斑和视网膜神经纤维层厚度存在差异,具体机制尚不清楚[9-12]。本实验选择与人类视觉系统高度相似的树鼩作为实验动物建立形觉剥夺性弱视模型,从形态学方面观察弱视形成及恢复过程中视网膜的变化。
1 材料与方法
1.1 实验动物及分组
本实验共选取30只出生后18 d尚未睁眼的树鼩幼崽,体重约50 g,性别不限,由中国医学科学院医学生物学研究所树鼩种质资源中心提供[SCXK(滇)K2018-0002]。入选树鼩需进行初步体检以排除眼部疾患及全身异常。
分组:将30只树鼩随机分为6组,每组5只:弱视模型N1组(左眼眼睑缝合2个月);弱视模型N2组(左眼眼睑缝合1个月);弱视恢复模型N3组(左眼眼睑缝合1个月后打开1个月);弱视恢复模型N4组(左眼眼睑缝合1个月后打开,换缝合右眼眼睑1个月);C1对照组(双眼正常视物2个月);C2对照组(双眼正常视物1个月)。树鼩由专业人员饲养,饲养条件符合规范。
1.2 方法
1.2.1 模型建立
树鼩称重后,使用30 g·L-1戊巴比妥(5 mL·kg-1)进行腹腔注射麻醉。手术区域消毒好后,用眼科显微剪剪去上下睑缘1 mm皮肤组织,止血后从内眦部开始行上下眼睑皮肤的缝合。操作结束后树鼩交由专业人员饲养,各组树鼩饲养条件相同。术后每天检查树鼩是否出现缝线脱落、缝合后眼部或全身感染等情况并及时处理,实验过程中淘汰不符合实验条件的树鼩。树鼩手术与实验操作均符合标准及规范,且通过了云南大学伦理道德委员会的审查。
1.2.2 HE染色
造模完成后,在对应时间(弱视模型N1、N3、N4组和C1对照组:出生后2个月零18 d;弱视模型N2组和C2对照组:出生后1个月零18 d)对各组内3只树鼩使用戊巴比妥过量麻醉后行颈椎脱臼处死。处死后尽快摘除眼球,在角巩膜缘后方进行穿刺,随后浸入组织固定液固定24 h,之后将标本取出,去除角膜、晶状体及玻璃体后将视网膜修剪为适合大小(保留视盘),然后组织脱水、石蜡包埋、切片后行HE染色,观察视网膜各层结构及感光细胞、双极细胞和神经节细胞数量变化。
1.2.3 透射电镜检查
造模完成后,各组内其余2只树鼩在对应时间使用戊巴比妥过量麻醉后行颈椎脱臼处死,取出眼球,用刀片切取视盘旁三分之一处约2 mm×2 mm大小的视网膜组织块,迅速放入体积分数3.5%戊二醛固定液中,随后进行电镜标本的制备、摄片,观察视网膜各层细胞的超微结构变化。
1.3 统计学分析
2 结果
2.1 组织学观察结果
本实验选取样本视盘旁三分之一处作为观察范围,固定光学显微镜放大倍数(×400)及图片截取宽度后摄片,进一步观察细胞结构和数量的改变(图1)。
2.1.1 视网膜形态学改变
光学显微镜下可见,C1、C2对照组树鼩视网膜由下至上可分为9层:视网膜色素上皮层、感光细胞层(视锥视杆细胞层)、外界膜、外核层、外丛状层、内核层、内丛状层、神经节细胞层、神经纤维层。
与C2对照组相比,弱视模型N2组树鼩视网膜中神经节细胞层厚度变薄,神经纤维层质地疏松,双极细胞层细胞层数明显减少,视锥视杆细胞内外节层与色素上皮层分离间隙增大(图2B黄色箭头处),细胞体积缩小、形态不规则且细胞间间隙增大;缝合时间更长的弱视模型N1组树鼩视网膜中神经节细胞层厚度进一步降低,神经纤维层萎缩(图2A黄色箭头),双极细胞层厚度进一步降低,细胞间隙扩大,视锥、视杆细胞形态较弱视模型N2组有一定程度的恢复;弱视恢复模型N3、N4组树鼩视网膜在各层均表现出一定程度的恢复,弱视恢复模型N3组树鼩视网膜的视锥视杆细胞核层及内外节层细胞间隙增大,排列紊乱(图2C黄色箭头),双极细胞核层厚度明显增加,细胞间隙增大;弱视恢复模型N4组树鼩视网膜视锥视杆细胞层细胞形态出现一定程度的恢复,但二者与C1对照组相比仍有差距(图2)。
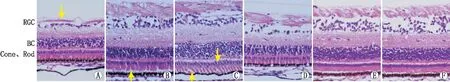
A:弱视模型N1组,黄色箭头示神经节细胞层;B:弱视模型N2组,黄色箭头示视锥视杆细胞内外节层与色素上皮层间隙;C:弱视恢复模型N3组,黄色箭头示视锥视杆细胞层及内外节层;D:弱视恢复模型N4组;E:C1对照组,F:C2对照组。RGC:神经节细胞层;BC:双极细胞层;Cone、Rod:视锥视杆细胞层。图2 各分组视网膜切片HE染色图(×400)
2.1.2 细胞数量的变化
感光细胞:除弱视模型N1与弱视恢复模型N4组、C1对照组与C2对照组间感光细胞数量差异无统计学意义(均为P>0.05)外,其余各组间感光细胞数量差异均有统计学意义(均为P<0.001)(表1)。
双极细胞:双极细胞数量在C1、C2对照组中最多,其余各组的双极细胞数量均低于C1、C2对照组(均为P<0.01);除弱视模型N1与弱视恢复模型N4组、C1对照组与C2对照组间双极细胞数量差异无统计学意义(均为P>0.05)外,其余各组间双极细胞数量差异均有统计学意义(均为P<0.001)(表1)。
神经节细胞:神经节细胞数量在C1、C2对照组中最多,两组间差异无统计学意义(P>0.05),弱视恢复模型N3组与C2对照组比较,差异无统计学意义(P>0.05),其余各组间神经节细胞数量差异均有统计学意义(均为P<0.001)(表1)。

表1 各组树鼩视网膜不同类别细胞数量
2.2 电镜观察视网膜各层细胞超微结构改变
(1)C2对照组:视锥视杆细胞、双极细胞、神经节细胞胞核圆润饱满,细胞膜完整、染色质丰富、分布均匀;视锥视杆细胞外节膜盘叠状结构明显且均匀,内节线粒体嵴规则、无皱缩;神经纤维分布均匀,内质网、核糖体未见明显异常(图3)。
(2)弱视模型N2组:视锥视杆细胞核体积缩小、异染色质增多、核不规则(图4A红色箭头处),双极细胞、神经节细胞可见线粒体肿胀、嵴溶解消失,内质网池形成(图4B、C红色箭头处),神经纤维水肿(图4D红色箭头处)、排列紊乱,视网膜细胞数较弱视模型N1组上升。
(3)弱视恢复模型N3组:视锥视杆细胞、双极细胞数量明显增多,细胞膜较完整,染色质稍有边集(图5A、C),视锥视杆细胞外节膜盘间隙增大,局部出现空泡变(图5B红色圆圈内),神经节细胞胞核固缩、体积缩小、异染色质增多、核不规则,细胞膜皱缩,细胞质浓缩(图5D),神经纤维规则分布(图5E)。
(4)弱视恢复模型N4组:视锥视杆细胞、双极细胞、神经节细胞数量正常(图6A、C、D红色箭头处),细胞形状稍不规则,染色质边集,部分可见线粒体肿胀,视锥视杆细胞外节膜盘间隙显著增大,部分可见空泡化(图6B红色圆圈内),神经纤维形态稍不规则(图6E红色圆圈内)。
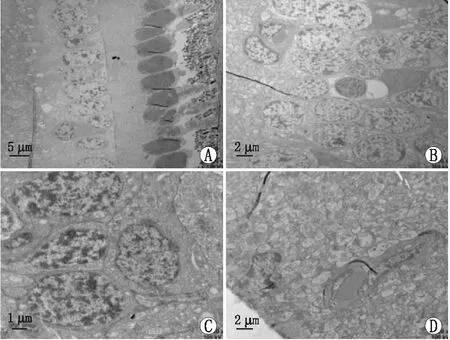
A:视锥视杆细胞层;B:双极细胞层;C:神经节细胞层;D:神经纤维层。图3 C2对照组视网膜细胞透射电镜图

A:视锥视杆细胞层,红色箭头示核体积缩小、异染色质增多、核不规则;B:双极细胞层,红色箭头示内质网池形成;C:神经节细胞层,红色箭头示内质网池形成;D:神经纤维层,红色箭头示神经纤维水肿。图4 弱视模型N2组视网膜细胞透视电镜图

A:视锥视杆细胞胞核层;B:视锥视杆细胞内外节层,红色圆圈示膜盘间隙增大,出现空泡化;C:双极细胞层;D:神经节细胞层;E:神经纤维层。图5 弱视恢复模型N3组视网膜细胞透视电镜图

A:视锥视杆细胞胞核层,红色箭头示视锥视杆细胞;B:视锥视杆细胞内外节层,红色圆圈示膜盘空泡化;C:双极细胞层,红色箭头示双极细胞;D:神经节细胞层,红色箭头示神经节细胞;E:神经纤维层,红色圆圈示神经纤维形态不规则。图6 弱视恢复模型N4组视网膜细胞透视电镜图
3 讨论
3.1 建立形觉剥夺性弱视模型的物种选择
过去60年间对于各种动物弱视模型的研究为我们探索弱视的发病机制和治疗方法提供了多种途径。形觉剥夺性弱视是唯一一种可以在所有物种中建模的弱视类型,目前应用于形觉剥夺性弱视研究的动物主要有猫、大鼠、豚鼠、松鼠、鸡、树鼩、恒河猴等,研究显示,单眼剥夺所导致的视力下降在不同物种之间存在显著差异[13-16]。
考虑到人类弱视的复杂性和多样性,以及视觉通路组织上存在的物种差异,某些动物模型其实并不能完美地模拟人类弱视的临床表现。不同的物种之间光栅灵敏度存在着巨大的差异,而这种差异可能反映了各个物种在视网膜解剖学上的根本差异(例如缺少中央凹以及视网膜中央锥体密度的差异)。
在不同物种之间,单眼剥夺后视力下降的程度也有很大差异。在进行单眼剥夺操作后,猕猴的弱视眼是彻底失明的[17],猫的弱视眼也是失明的,但一段时间后会有一定程度的恢复[18],而啮齿动物模型的弱视眼光栅灵敏度仅降低了一个八度。这些差异是源于物种间视觉通路组织解剖或生理上的差异,还是源于物种间神经回路可塑性的差异,目前还没有达成一致意见。
树鼩视觉发达,具有一些灵长类动物的视觉特征(已初步具有双眼视觉或立体视觉),并且已经完成了全基因组的测序[19],在神经和免疫系统方面与人类高度同源,与灵长类相比,实验成本更低且更易获取。尽管树鼩的视网膜厚度与人类相比较薄,但其视网膜结构及主要组成细胞与人类极为相似[20]。本课题组前期已建立形觉剥夺性弱视的树鼩模型[21],视觉诱发电位检测也已证实此模型建立成功[22]。因此,本实验选取树鼩作为实验动物建立形觉剥夺性弱视模型以探究弱视形成及恢复过程中视网膜的变化特征。
3.2 单眼形觉剥夺树鼩视网膜组织学及超微结构
在既往研究中,关于弱视形成过程中视网膜和视神经的结构是否出现改变这一观点是存在争论的。一些研究报道了黄斑和视网膜神经纤维层厚度存在显著差异[23-26],然而其他一些研究表示没有发现明显的视网膜结构差异[27-28]。早期的Meta分析表明弱视视网膜黄斑厚度增加[29], 然而近几年的报道回顾了弱视中OCT结果后,认为在弱视中黄斑和视网膜神经纤维层变化的证据仍然是存在争议的[5]。
本研究主要观察感光细胞、双极细胞、神经节细胞的改变,发现单眼形觉剥夺会对树鼩视网膜各层细胞产生不同程度的损害。在光学显微镜下观察剥夺眼视网膜HE染色发现:神经节细胞核层细胞减少,厚度变薄,双极细胞核层细胞层数明显减少,厚度变薄,视锥视杆细胞内外节层以及细胞核层细胞体积缩小、形态不规则且细胞间间隙增大,细胞数量减少。在缝合时间更长的组中,除上述变化外,神经节细胞和双极细胞的数量进一步减少,神经纤维层出现明显萎缩,推测随着剥夺时间的延长,视网膜损伤加重,弱视严重程度也随之加深。通过透射电镜观察到剥夺眼视网膜出现了细胞核体积缩小、核不规则、异染色质增多、线粒体肿胀、内质网池形成等损伤表现。
关于视网膜功能是否存在损害这一问题,目前尚无定论。一些研究小组报道了视网膜电图的振幅降低和延迟[30-32],但也有其他研究称屈光参差性斜视受试者的视网膜反应没有差异[33],出现这样的结果可能是由于其他疾病被误诊为弱视或是疾病与弱视同时存在。由于视神经发育不全是单眼弱视的基础,所以当这些原因同时存在的时候,通常很难确定视网膜异常是弱视直接导致的,还是基于其他原因出现的损伤。视网膜损伤可能是弱视的原因,也可能是进一步的神经损伤引起的代偿机制。总之,弱视与视网膜结构和功能之间的确切关系仍不清楚。虽然某些视网膜变化可能与弱视有关,但尚不清楚这些变化可能会导致什么样的病理结果,究竟是视网膜的损伤导致了弱视,还是视觉通路上的神经损伤导致了视网膜的代偿,这个问题还有待探究。
3.3 弱视的治疗
本研究分别模拟了两种不同的弱视治疗方式:去除剥夺后自然恢复和去除剥夺后遮盖优势眼。在光学显微镜下观察两组弱视恢复树鼩模型的视网膜HE染色发现:缝合左眼1个月后打开1个月,视锥视杆细胞胞核层及内外节层细胞间隙增大,排列紊乱,数量增多,双极细胞核层细胞数目减少,细胞间隙增大。缝合左眼1个月后打开换缝右眼1月,视锥视杆细胞、双极细胞、神经节细胞数量均有明显减少。两个弱视恢复模型的透射电镜结果显示:两组树鼩视网膜中均存在不同程度的细胞损伤,包括染色质边集、外节膜盘间隙增大、线粒体肿胀等,而当去除损害因素后视网膜细胞出现不同程度的恢复,说明其具有可塑性。
通过对比两个弱视恢复模型的组织学及透射电镜结果后发现,去除剥夺组中视网膜各层细胞的数量和形态较去除剥夺前均有一定程度的恢复,其中以视锥视杆细胞的数量增多最为明显。HE染色结果显示,视锥视杆细胞胞核层及内外节层呈现出多层结构,细胞间隙增大,内外节层排列紊乱,电镜结果也显示同样的多层结构。但既往研究表明,树鼩的视网膜感光细胞为单排排列结构[34-35],这是树鼩感光细胞的特征。在对此组进行多次重复实验之后,基本可以排除在切片过程中有各层细胞错位这一可能,我们更倾向于认为是在去除剥夺后某些信号通路被激活,从而引起了视网膜感光细胞的快速增生。但究竟有哪些信号通路参与了这一过程,这样快速增生的细胞是否存在功能缺陷,或者细胞间间隙增大及排列紊乱是否会导致细胞间信号传递的延迟,后续还需进一步探究。
同时我们也观察到,在弱视恢复模型N4组中视网膜各层细胞出现了新的损害。既往研究发现,弱视的影响并不仅局限于弱视眼,非弱视眼在一定程度上受到弱视眼的抑制[36],其在空间、位置和运动敏感性方面也存在损害[37]。树鼩的婴儿期是从出生到出生后30 d[38],而我们开始反转缝合的时间是在树鼩出生后48 d左右,仍处于树鼩视觉发育的关键期内。所以我们推测此时的树鼩仍对形觉剥夺敏感,可能是反转缝合后对侧眼对原弱视眼的抑制引起了新的损害,也可能是最初的单眼剥夺引起的损害尚未恢复,亦或是两种原因同时存在。究竟是什么原因所致,还需要后期进一步研究加以证实。
由于目前弱视发病机制尚未明确定论,许多弱视治疗效果不理想。目前国内外针对弱视的治疗多为遮盖优势眼、屈光矫正及视觉训练。动物实验表明,压抑优势眼对弱视的治疗效果与关键期相关。在猕猴中,从出生到大约10周的关键期内,单眼剥夺显著干扰了条纹皮质中典型的眼部优势柱的发育,具体表现为早期视觉皮质中双眼反应细胞数量减少,对空间频率的敏感性改变[17]。为了评估从这种剥夺中恢复的可能性,研究者对一部分动物进行了反向缝合。当在6周左右进行反向缝合时,眼优势柱结构是有可能恢复正常的。然而在1岁时进行反向缝合时,几乎是不可能恢复的,这再次说明早期发育后可塑性降低。多项研究表明,相较于单纯的遮盖治疗,视觉训练可以显著改善青少年和弱视成人的视敏度和视觉空间能[39-41]。现阶段,应用于临床弱视治疗的药物仍以左旋多巴和胞苷二磷酸胆碱为主[42-43],但随着对弱视致病机制认知的深入,针对视神经通路可塑性的研究也在逐步开展。
4 结论
本研究结果表明,在树鼩形觉剥夺性弱视模型中,树鼩视网膜各层细胞均有明显改变,并且具有一定的可塑性。虽然目前弱视的定义仍认为眼部无器质性病变,本研究结果明确显示弱视眼视网膜部位存在损伤,只是可能这种损伤处于细胞水平,目前临床上的常规检查无法探测到。本研究可能为形觉剥夺性弱视的发病机制及治疗提供新的思路。
