利用数据库解析高原红细胞增多症与肺动脉高压基因表达差异的关联性*
2023-07-14申杨磊方龙伟多吉卓玛巴桑卓玛
申杨磊,方龙伟,央 拉,多吉卓玛,巴桑卓玛
西藏大学高原健康科学研究中心,西藏拉萨 850000
高原红细胞增多症(HAPC)是高原地区常见的慢性高原病之一,其主要特征为红细胞过度增生,表现为血红蛋白浓度显著增加,其病变多呈慢性过程,后期常伴有全身多系统、器官、组织不同程度损害[1]。
根据世界卫生组织的分类,肺动脉高压(PH)可分为动脉型PH、左心疾病所致PH、肺脏疾病和(或)低氧所致PH、慢性血栓和(或)栓塞性PH、多种未明机制所致PH 5类[2]。其中高原肺动脉高压(HAPH)属于第3类,是由于高原低氧环境,肺换气代偿性增加、血细胞比容升高、血管收缩引起的一种慢性高原病[3],与HAPC一样,也是非常典型的危害高原人群健康的疾病,而且现有研究表明,HAPC与PH间存在共同的病理特征:(1)红细胞与血红蛋白增加;(2)肺泡壁明显增厚,纤维增粗,弥散功能下降;(3)肺动脉压力变化与红细胞过度增生有关。HAPC的主要表现为红细胞与血红蛋白异常增加,对PH的文献查阅结果同样发现红细胞与血红蛋白增加,而红细胞与血红蛋白异常增加可对机体心、肺、肾等脏器功能造成损伤。此外,还有学者通过研究过度表达人类促红细胞生成素(EPO)的转基因小鼠,分析小鼠血液中红细胞增生情况与肺动脉压力的变化,发现PH的发生与红细胞过度增生有直接关系[4-5]。有学者研究发现,HAPC患者肺泡壁明显增厚和纤维增粗,使肺泡间质弥散功能下降,红细胞的过度增加使肺动脉压力明显升高[6-8]。以上相关研究均表明HAPC与PH间具有相同的发病特征。但目前检索到的HAPC与PH两种疾病间的相关性研究较少,因此利用生物信息学工具对两种疾病的相关数据进行检索,筛选出共同的差异基因和信号通路,但因目前数据库中关于HAPH相关基因的数据太少,本文就HAPC与PH的基因差异进行比较和分析,旨在为进一步探讨HAPC与PH之间发病机制的异同及关联提供理论依据。
1 材料与方法
1.1数据来源 使用GEO数据库(https://www.ncbi.nlm.nih.gov/geo/)对以人类为研究样本的HAPC与PH基因数据进行检索,获取GSE29977[9]、GSE53408[10]、GSE703[11]、GSE113439[12]4个数据集,对其研究选择的样本与研究方法进行筛选、排除后保留GSE29977与GSE53408数据集。GSE29977芯片属于GPL570平台[(HG-U133_Plus_2)Affymetrix Human Genome U133 Plus 2.0 Array]基因芯片,共包括10个样本,其中5例HAPC患者、5例健康对照者;GSE53408芯片属于GPL6244平台[(HuGene-1_0-st)Affymetrix Human Gene 1.0ST Array transcript(gene)version]基因芯片,共包括23例样本,其中12例PH患者与11例健康对照者。
1.2差异基因筛选 通过R语言对数据集进行均一化处理后,使用Limma包[13]对数据中的差异基因进行筛选,然后在筛选结果基础上使用Bonferroni法对P值进行校正,GSE29977数据集校正后P值应<0.01;GSE53408数据集校正后P值应<0.004 54;差异基因的筛选标准应满足校正后P值且满足log2|FC|>1。得到两个数据集中符合标准的差异基因,再绘制韦恩图进行重叠筛选,得到两个数据集中共同的差异基因。
1.3基因本体(GO)分析、京都基因与基因组百科全书(KEGG)分析、基因富集(GSEA)分析 DAVID数据库(https://david.ncifcrf.gov/)是具有多种生物数据和分析工具的软件,可对目的数据进行富集分析,并提供注释信息。本研究通过DAVID数据库对筛选得到的共同差异基因进行GO分析、KEGG富集分析和功能注释,筛选条件为P<0.05。GO分析结果由3个部分构成:生物学过程(BP)、分子功能(MF)及细胞成分(CC)。GSEA分析是将所有基因纳入研究再筛选出差异显著的富集通路,筛选条件为P<0.05。
1.4GeneCards分析 GeneCards数据库(https://www.genecards.org/)包括了多种生物的基因数据和大量疾病相关基因,该数据库对相关基因进行细致的分类,可以查阅到基因分布的组织、作用途径和与疾病相关度的得分。故利用GeneCards数据库对筛选到的差异基因进行检索,观察差异基因是否与相关疾病的致病基因相符、在人体组织中的分布是否与疾病相关。
2 结 果
2.1HAPC与PH基因筛选 GSE29977数据集标准化后进行富集分析,对富集出的差异基因进行筛选,得到1 627个差异基因(815个上调基因,812个下调基因),校正后符合筛选阈值(P<0.01;log2|FC|>1)的基因有396个,其中188个上调基因和208个下调基因。GSE53408数据集标准化后进行富集分析,筛选得到3 455个基因(695个上调基因,2 760个下调基因),校正后符合筛选阈值(P<0.004 54;log2|FC|>1)的基因有791个,其中695个上调基因和96个下调基因。将两个数据集转换为一致的基因ID并排除其中无名称基因,对剩余基因使用维恩图进行重叠筛选,共得到138个共同差异基因,校正后共同的差异基因有11个,分别为内皮细胞黏附分子(ESAM)、成纤维细胞生长因子18(FGF18)、主要组织相容性复合体(MHC)Ⅱ类人类白细胞抗原DQB1(HLA-DQB1)、同源异构体A7(HOXA7)、POZ/BTB和AT结合基序锌指蛋白1(PATZ1)、整合素亚基B4(ITGB4)、甲基硫核糖-1-磷酸异构酶(MRI1)、基质金属肽酶8(MMP8)、Caspase12、10号染色体开放阅读框25(C10orf25)、H因子(CFH)。
2.2GO分析及KEGG分析 对138个差异基因进行功能注释,然后进行GO分析以及KEGG分析。校正前,BP差异基因主要富集在:中胚层形态生成、初级胚层形成、原肠胚形成;CC差异基因主要富集在:黏附连接、细胞与细胞连接、蛋白酶体调控颗粒碱基亚复合物、间质矩阵、宿主细胞内部分等;MF差异基因主要富集在:激活转录因子、MHCⅡ类受体活性、UDP葡萄糖基转移酶活性、腺苷脱氨酶活性、磷酸酪氨酸残基结合等。KEGG分析提示差异基因主要富集在:内质网蛋白质加工、细胞外基质(ECM)受体相互作用、金黄色葡萄球菌感染、半胱氨酸和蛋氨酸代谢等。
先对校正后的11个差异基因进行功能注释,然后进行GO分析及KEGG分析。BP差异基因主要富集在初级胚层形成、原肠胚层形成,CC差异基因主要富集在MHCⅡ类蛋白复合物、复杂炎性体、核膜,MF差异基因主要富集在MHCⅡ类受体活性、半胱氨酸型内肽酶活性参与凋亡信号通路、胰岛素样生长因子Ⅰ结合等。KEGG分析提示差异基因主要富集在金黄色葡萄球菌感染、细胞黏附分子、肌动蛋白细胞骨架的调节。见图1~4。
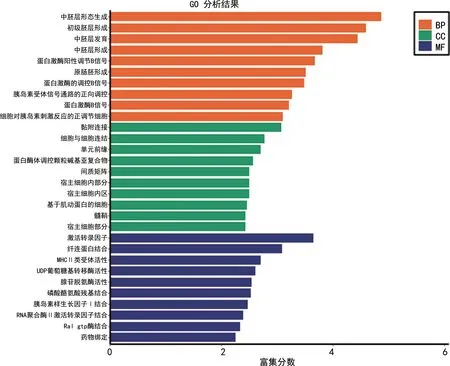
图1 校正前差异基因GO分析结果

图2 校正后差异基因GO分析结果

图3 校正前差异基因 KEGG分析结果

图4 校正后差异基因KEGG分析结果
2.3GSEA分析 由于得到的数据集差异基因较少,进一步使用GSEA分析对差异基因附近的通路进行分析,探索HAPC与PH的共同性。GSE53408数据集的GSEA分析结果显示只有两个通路具有显著差异。细胞周期信号通路具有最高标准化富集得分[NES,NES=2.236 2,P<0.000 1,错误发现的比率(FDR)<0.014],其次为卵母细胞减数分裂通路(NES=2.014 0,P<0.001 2,FDR<0.027)。GSE29977数据集的GSEA分析结果并未发现两种疾病间共同的信号通路,需要在未来进行进一步研究分析。见表1。

表1 GSEA分析筛选GSE29977数据集差异富集的信号通路
2.4GeneCards分析 通过GeneCards网站对筛选出的138个共同差异基因和校正后得到的11个差异基因数据进行分析。对未校正前筛选到的131个(有7个基因数据库未能识别)共同差异基因检索结果显示,差异基因在人体多个组织均有表达,由于PH属于心血管系统疾病(或循环系统疾病),故对心血管系统疾病的分类进行检索分析,结果显示与这131个共同差异基因相关的疾病中包含PH。对筛选到的138个差异基因与人类PH所有相关基因进行比对,结果显示相同的基因有60个,与PH高度相关的基因有血管紧张素转换酶(ACE)、微小RNA、SMAD通路蛋白(SMAD)、纤维母细胞生长因子(FGF)、血小板衍生生长因子(PDGF)。
对校正后的11个差异基因进行对比,显示有5个相同基因:FGF18、HLA-DQB1、PATZ1、MMP8、CFH。结果在疾病分类中未查阅到PH。对校正后11个差异基因进行检索,结果显示其中4个基因在肺部高表达,包括ESAM、FGF18、HLA-DQB1、PATZ1。PATZ1主要表达于肺茎支气管中;ESAM表达于肺血管内皮细胞表面;HLA-DQB1表达在肺、血液、呼吸道上皮细胞表面、支气管中部。对相关疾病的分析结果显示,高血压分类与PH相关性评分较高(相关性得分为34.1),故推测HAPC与PH间可能存在相关性。
3 讨 论
通过知网和Pubmed两个数据库对本次筛选出的相关基因进行检索,发现与本次筛选结果相同的基因有ACE、PDGF、HLA-DQB1。研究表明,ACE是肾素-血管紧张素系统产生,具有强烈的血管收缩作用。机体在低氧环境下促使ACE产生且受低氧诱导因子(HIF)调控,傅薇[14]研究表明HAPC患者骨髓单个核细胞、骨髓液、血液及骨髓的ACE水平均增高,证明ACE是HAPC的发生机制之一,同时ACE的同素体ACE2广泛分布在心、肾、肺等器官,在肺中分布于Ⅰ型肺泡上皮细胞、Ⅱ型肺泡上皮细胞、支气管上皮细胞、巨噬细胞、肺血管内皮细胞、平滑肌细胞。相关研究表明ACE可以促进血管紧张素2(AngⅡ)产生,AngⅡ可以诱发肺血管内平滑肌细胞迁移,刺激炎症因子、转化生长因子-β(TGF-β)产生,诱导肺实质细胞凋亡,AngⅡ还可进一步促进血管收缩及平滑肌细胞生长迁移,加重PH发生[15]。查阅现有PDGF相关研究结果显示,PDGF家族包括PDGFA、PDGFB、PDGFC、PDGFD等。PDGF具有趋化、收缩血管、促进分裂作用,参与磷酸酯酶激活与前列腺素代谢。殷玉娟等[16]研究显示,HAPC患者血小板活化,PDGF表达升高,血栓形成使得血液成分和血流状态改变[17],可以使HAPC患者血氧饱和度下降10%[18],加重血管细胞损伤,促使血栓形成增加,故PDGF是HAPC的发病原因之一。彭虹艳[19]研究发现,PDGF可以在有氧环境下促进主动脉血管平滑肌细胞糖酵解增强,PDGF通过PI3K/AKT/mTOR信号通路上调HIF-1α,使得丙酮酸脱氢酶(PDH)磷酸化活性下降,产生肺动脉平滑肌Warburg效应,使得肺动脉平滑肌异常增殖、肺动脉重构,证明PDGF在PH的发生过程中发挥重要作用。检索现有与人类白细胞抗原系统(HLA)相关文献,结果显示其具有高度基因多态性,其中DQA1和DQB1具有多种免疫学功能,且已有研究表明HLA除与免疫系统疾病相关外,还与呼吸系统、消化系统、心血管系统疾病存在相关性[20]。青格乐图等[21]对青海地区HAPC患者人类白细胞抗原DQA1(HLA-DQA1)和HLA-DQB1的6个等位基因与健康对照者进行对比,结果发现HLA-DQB1与肺动脉高压的发生密切相关[22]。通过对已有文献进行查阅,与本次分析筛选到的差异基因进行对比,结果显示有多个已验证的疾病相关基因,其余未验证的差异基因需在未来进行实验验证。
本研究还存在以下不足:(1)在GEO数据库中检索到的研究样本较少;(2)GeneCards在线数据库分析结果与目前已有的基因研究结果相符的数据较少;(3)GSEA分析结果未发现共同的信号通路,需进一步进行分析研究;(4)本研究仅是对GEO数据库中已有的相关研究数据进行分析研究,需要进行实验验证。
综上所述,本研究为HAPC与PH的相关研究提供了思路和研究方向,下一步可对筛选的差异基因进一步分析,并进行功能验证进而探索不同的PH类型与HAPC的相关性。
