盐胁迫对“宝岛蕉”光合作用和植株养分分布特征的影响
2023-06-15兰柳艳黄虹心汤狄华项俊华罗晓慧
兰柳艳,王 蕾,沈 欢,黄虹心,汤狄华,项俊华,罗晓慧,武 鹏
(1 广西农业职业技术大学,南宁,530007;2 广西贺州市检验检测中心,广西贺州,542800;3 广西农业科学院生物技术研究所,南宁,530007)
盐害是农业生产中常见的阻碍植物正常生长的因子。近年来由于不合理的灌溉施肥、盐碱地种植以及海水浇灌等,使得植物常遭受盐胁迫。盐胁迫主要影响光合作用和根部养分吸收两个过程,两个过程对作物的生长发育均至为重要[1]。广西南端具有面向南海的绵长海岸线,是我国唯一沿海的自治区,从北海市到防城港市,广西海岸线长度1 500 km左右。海岸线上广泛种植香蕉,海水常倒灌进陆地,造成高浓度的盐胁迫;每年8—9月为台风高发期,从南海上形成的水汽随台风进入内陆,也会引起低浓度的盐胁迫[2]。香蕉是著名的热带水果,产量仅次于柑桔,居世界第二位[3]。广西是我国第二大香蕉产区,常年遭受海水倒灌和台风,广西沿海地区香蕉产业受到严重影响[4-5]。因此,研究盐胁迫对香蕉生长的影响、分析其耐盐机制对于广西香蕉产业具有重要意义。“宝岛蕉”是目前我国香蕉主推品种,因其较高的抗枯萎病特性及高产、稳产等优点,在我国香蕉枯萎病疫区种植面积不断扩大。因此,我们以“宝岛蕉”幼苗为试材,研究盐胁迫对其光合作用和植株养分分布特征的影响,分析其耐盐机制,助推沿海地区“宝岛蕉”高质量发展。
1 材料与方法
1.1 试验设计
2020年在广西北海市合浦县进行试验,2020年3月“宝岛蕉”香蕉组培营养杯苗移栽大田,株行距2.0 m×2.2 m,种植密度为150株/667 m2,统一常规管理,选择植株健康、生长一致的20叶苗供试。海水Na+浓度达到0.80%[2],故设置NaCl胁迫浓度(质量体积比)梯度为0%(对照)、0.01%、0.05%、0.10%、0.20%、0.40%、0.60%、0.80%,每处理重复30株,试验当天每株浇不同浓度盐水10 L后,每天滴灌滴水2 L保持土壤湿润,7 d后测定相关指标。
1.2 测定指标与方法
光合指标测定。叶片叶绿素含量提取采用混合液浸提法,在波长663、645 nm下测定吸光度,计算叶绿素总量[6]。9:00选取各处理植株从上往下数第3、4、5片叶,用暗适应夹遮光处理30 min后,10:00用配备 6400-40叶绿素荧光叶室的便携式光合测定系统(LI-6400XT,LI-COR)检测叶片净光合速率(Pn)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)、初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv=Fm-Fo)、PSII原初光能转换效率(Fv/Fm)等参数。
植株营养成分测定。采集试验区域中心长势基本一致的健康植株,挖取整株,每处理重复3株,每株分成根、球茎、假茎和叶片等4个部位,就地称量各部位鲜质量,按照四分法选取各部位具有代表性的植物组织作为分析样品[7],每部位鲜样质量大于500 g。新鲜样品洗净后用吸水纸吸干水分,105 ℃下杀青15 min,70~80 ℃下烘干至恒重,参照鲍士旦的方法[8],全氮采用凯氏定氮法,全磷采用钼锑抗比色法,全钾采用火焰光度计法,镁、钙、钠采用干灰分-原子吸收分光光度计法,氯采用硝酸银滴定法测定。
1.3 数据统计与分析
使用SPSS 22.0进行差异显著性分析(Duncan’s,p<0.05)。
2 结果与分析
2.1 对叶片光合特性的影响
由表1可以看出,盐浓度≤0.2%时,不同处理的叶绿素含量与对照差异不显著;当盐浓度≥0.4%时,随着盐浓度增大,叶绿素含量呈逐渐下降趋势,各处理间差异显著。盐浓度≤0.05%时,不同处理的气孔导度、蒸腾速率和净光合速率与对照差异不显著;当盐浓度≥0.05%时,不同处理气孔导度、蒸腾速率和净光合速率呈逐渐下降的趋势,各处理间差异显著。盐浓度≤0.05%时,不同处理的胞间CO2浓度与对照差异不显著;当0.05%<盐浓度<0.4%时,胞间CO2浓度逐渐下降;当盐浓度>0.4%时,胞间CO2浓度逐渐上升。Fv/Fm为PSII系统的原初光能转换效率[9],不同盐浓度处理的Fv/Fm与对照差异不显著,说明经受盐胁迫时,“宝岛蕉”对盐胁迫具有一定程度的耐受力,不同盐浓度胁迫均未对“宝岛蕉”光能转化系统造成显著性影响。

表1 不同盐浓度胁迫对“宝岛蕉”20叶幼苗叶片叶绿素含量和光合参数的影响
2.2 对植株各部位营养元素含量的影响
由表2可以看出,相同盐浓度胁迫下各部位全氮含量以叶片全氮含量最高,全氮含量从高到低依次为叶片>球茎>假茎>根。盐浓度0.01%处理的各部位全氮含量与对照差异不显著;盐浓度0.01%~0.20%处理时,各部位全氮含量随着盐浓度的升高缓慢下降;盐浓度0.2%~0.8%处理时,各部位全氮含量迅速下降,各处理间差异显著。与全氮含量不同,相同盐浓度胁迫下假茎的全磷、全钾含量高于其他部位,全磷含量从高到低依次为假茎>叶片>球茎>根,全钾含量从高到低依次为假茎>根>球茎>叶片;盐浓度0.01%处理的各部位全磷、全钾含量与对照差异不显著;盐浓度0.01%~0.20%处理时,各部位的全磷、全钾含量随着盐浓度升高缓慢下降;其中盐浓度0.05%处理叶片全磷、全钾含量与对照差异不显著;盐浓度0.2%~0.8%处理时,各部位全磷、全钾含量随着盐浓度的升高而快速下降。
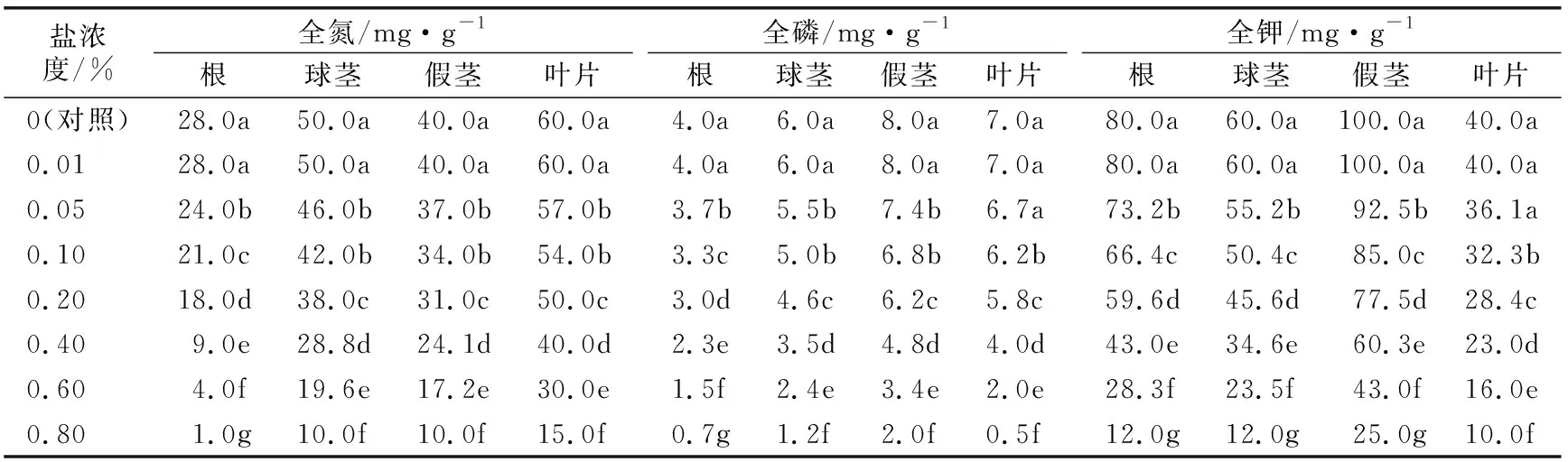
表2 不同盐浓度胁迫的“宝岛蕉”20叶幼苗植株各部位全氮、全磷和全钾含量变化
2.3 对各部位全镁、全钙含量的影响
由表3可以看出,叶片的全镁、全钙含量高于其他部位,相同盐浓度处理的全镁含量从高到低依为叶片>假茎>球茎>根;在盐浓度0.01%处理时,各部位的全镁、全钙含量与对照差异不显著;盐浓度0.01%~0.20%处理时,各部位的全镁、全钙含量随着盐浓度的升高缓慢下降;盐浓度0.2%~0.8%处理时,各部位全镁、全钙含量随着盐浓度的升高而快速下降;盐浓度0.01%处理时,叶片的全镁含量与对照差异不显著,之后开始缓慢下降,没有快速下降的趋势;盐浓度≤0.20%时,叶片的全钙含量与对照差异都不显著;盐浓度0.2%~0.8%处理时,叶片全钙含量开始缓慢下降,没有快速下降的趋势;叶片的全镁、全钙含量在各盐浓度的下降比例均小于其他部位。
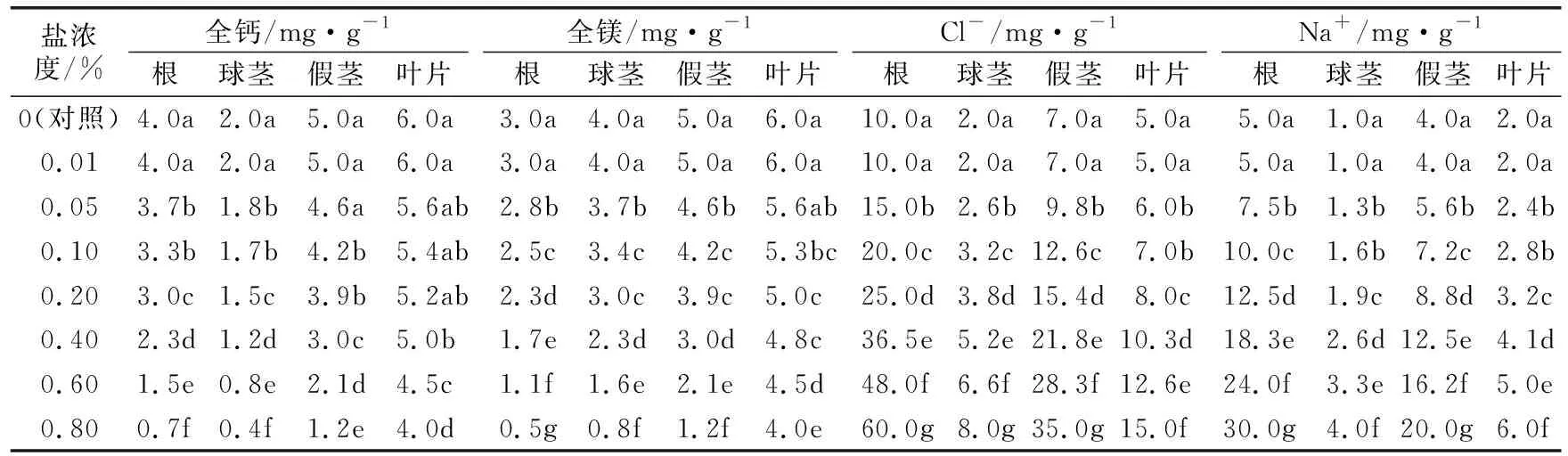
表3 不同盐浓度胁迫的“宝岛蕉”20叶幼苗植株各部位全钙、全镁、Cl-和Na+含量的变化
2.4 对各部位Cl-、Na+含量的影响
由表3可以看出,相同盐胁迫处理的植株根系的Cl-、Na+含量均高于其他部位,Cl-、Na+含量从高到低依次为根>假茎>叶片>球茎;盐浓度0.01%处理时,各部位Cl-、Na+含量与对照差异不显著;盐浓度>0.01%处理时,叶片和球茎的Cl-、Na+含量随着盐浓度的升高而缓慢升高,根和假茎的Cl-、Na+含量随着盐浓度的升高而快速升高;根和假茎的升高幅度远大于叶片和球茎,说明“宝岛蕉”的根和假茎部位具有较高聚集Na+的作用。
3 结论与讨论
叶绿素含量直接决定植物光合作用的潜力[10]。盐胁迫下植物叶片中叶绿素含量下降的主要原因是盐胁迫提高了叶绿素酶的活性,促进了叶绿素降解[11]。本研究中,一定浓度盐胁迫并没有造成叶片叶绿素含量显著变化;盐浓度>0.20%时,叶片叶绿素含量才逐渐降低,与对照差异显著。这与菠菜、银杏等[12-13]植物面临盐胁迫时,叶绿素含量变化相似。
气孔是植物与外界进行气体交换的主要通道,叶片通过气孔吸收CO2,气孔导度的高低决定了叶片吸收CO2的能力,一般认为盐胁迫引起叶片水势及气孔导度降低,限制了CO2到达光合机构,从而抑制光合作用和蒸腾作用[14]。本研究中,低浓度盐(盐浓度≤0.05%)胁迫并没有造成气孔导度的变化,当盐浓度>0.05%时,气孔导度才逐渐下降,说明“宝岛蕉”有一定的抗盐能力。当气孔导度逐渐下降时,叶片的净光合速率、蒸腾速率都随之逐渐减小。这与菜豆、小麦等植物相似,气孔因素是影响光合作用的主因[15-16]。虽然盐浓度升高,胞间CO2浓度呈先下降后升高的趋势,这可能是高浓度盐胁迫下净光合速率持续降低,对CO2利用减少,导致CO2过剩;这和盐胁迫时黄瓜的表现一致,均呈现出高浓度盐胁迫下胞间CO2过剩的情况[17]。
光合作用有光反应和暗反应两个过程。叶绿体上有两套光合作用系统,即光合作用系统一(PSI)和光合作用系统二PSII,在光照条件下,两个系统分别吸收700 nm和680 nm波长的光子作为能量,将从水分子在光解过程中得到的电子不断传递[9]。Fv/Fm代表PSII系统的原初光能转换效率。本研究中,任何浓度的盐胁迫7 d后,其Fv/Fm与对照无显著性差异,说明一定范围盐胁迫并没有造成“宝岛蕉”光反应阶段PSII系统的破坏。
由于叶片蒸腾拉力的作用,水分从土壤经过根细胞吸收,经过根和茎的导管最终流入水势最低的叶片里。在此过程中,土壤中的NO3-、NH4+、H2PO4-、K+、Mg2+、Ca2+等各种离子随着水分一起进入植物体内[18]。在盐胁迫存在的条件下,土壤中由于较高的盐分而降低了根细胞的吸水能力,土壤中的NO3-、NH4+、H2PO4-、K+、Mg2+、Ca2+等进入植物体内的量也随之减少;另一方面,盐离子与各种营养元素相互竞争进入细胞也会造成植物养分离子吸收的减少[19]。本文研究结果表明,盐胁迫导致“宝岛蕉”植株各部位全氮、全磷、全钾、全镁以及全钙营养元素的降低,这与很多作物相似,如菠菜、巴西蕉等[20-21]。虽然盐胁迫导致植物对养分离子吸收的普遍降低,但对养分离子吸收量分配比例不同。如耐盐的菠菜品种叶片选择性吸收K+、Ca2+、Mg2+,以降低Na+对叶片的影响[20]。本文研究结果表明,各浓度盐胁迫下,“宝岛蕉”叶片和球茎的K、Ca、Mg元素的下降比例远低于其他部位;说明遭受盐胁迫时,其叶片和球茎可能具有选择性吸收K+、Ca2+、Mg2+的能力,“巴西蕉”也被证明了这一现象[21]。
为维持细胞内微环境的稳定,调节Na+的分布是高等植物的重要耐盐机制之一。植物体除了在细胞水平上使Na+外排以减少细胞对Na+的吸收之外,还对Na+在不同部位的分布进行调节以降低其对植物的伤害。植物体内存在明显的排Na+机制,主要涉及1个或者多个过程[22]。即(1)根细胞不吸收Na+,即使进入根细胞也通过Na+/H+泵排出;(2)植物将吸收的Na+贮存于根茎基部,从而阻止Na+向叶片运输;(3)植物吸收的Na+在运输过程中,被茎木质部或韧皮部传递细胞吸收;(4)植物吸收的Na+向地上部特别是叶片和果实运输的选择性降低,而K+运输选择性增加。植物通过这些机制有效降低体内尤其是地上部的Na+含量,缓解盐胁迫的伤害。本文研究结果表明,“宝岛蕉”在盐胁迫的过程中,根和假茎部位的Cl-和Na+含量增加量远大于叶片和球茎的增加量。这表明“宝岛蕉”耐盐机制可能是将较多的Na+贮存于根部,从而阻止部分Na+向叶片运输,及Na+在运输过程中被茎木质部或韧皮部传递细胞吸收。这与“巴西蕉”的耐盐机制相同[22]。耐海水菠菜品种根系和叶柄以及水稻根部和叶鞘具有较高聚集Na+的作用,具有同样的耐盐机制[20,23]。
综上所述,“宝岛蕉”遭受盐胁迫时,低浓度盐并未对光合作用造成影响;高浓度时,随着盐浓度增加光合效率逐渐降低。“宝岛蕉”遭受盐胁迫时并没有使PSII系统的原初光能转换效率降低,而是导致叶绿素含量和气孔导度降低,说明光合作用降低是由于叶绿素含量下降和气孔导度较低,从而导致CO2这一原材料不足。遭受盐胁迫过程中,“宝岛蕉”抑制了根部营养元素吸收,导致植株各部位营养元素含量下降;其中各部位的N、P含量下降比例相似,而叶片和球茎K、Mg、Ca含量下降比例远低于其他部位,表明遭受盐胁迫时,“宝岛蕉”叶片和球茎可能具有选择性吸收K+、Ca2+、Mg2+的能力。根和假茎部位的Cl-和Na+含量的增加量远大于叶片和球茎的增加量,表明“宝岛蕉”耐盐的机制可能是通过根和假茎积累较多的Cl-和Na+,从而阻止向叶片运输。
