不同生境剑叶金鸡菊瘦果和种子的特性
2023-06-08徐德聪王宇沈章军陈延松
徐德聪 王宇 沈章军 陈延松

摘要: 為探讨不同生境对剑叶金鸡菊(Coreopsis lanceolata L.)的瘦果、种子特性的影响,揭示其环境适应机制,本研究采集了安徽省铜陵市郊区山坡上(生境Ⅰ)、铜陵市杨山冲尾矿库区(生境Ⅱ)和尾矿库覆土复垦区(生境Ⅲ)3种不同生境下的剑叶金鸡菊成熟瘦果,测定其长、宽、厚、千粒质量、密度、含水率等形态、品质指标以及重金属、钙和总磷的含量,利用生物体视显微镜、扫描电子显微镜观察其外观形态和内部构造。结果表明,不同生境剑叶金鸡菊瘦果和种子有相似的形态构造,可区分于其他种,但附属结构及种子表面微形态存在差异。不同生境剑叶金鸡菊瘦果的长、宽、体积和千粒质量均表现为生境Ⅰ和生境Ⅱ显著大于生境Ⅲ(P<0.05),生境Ⅰ与生境Ⅱ之间无显著差异(P>0.05),瘦果含水率表现为生境Ⅰ显著大于生境Ⅲ,生境Ⅲ显著大于生境Ⅱ。不同生境剑叶金鸡菊瘦果的厚和密度均无显著差异。主成分分析结果表明,3个生境分为不同的区域,进一步证明不同生境间瘦果形态和品质出现了差异。瘦果对重金属的积累并没有随着基质中重金属含量的增加而增加,但矿区生境瘦果中钙、总磷含量均显著高于山坡生境。相关性分析结果表明,基质中Cu、Zn含量均与瘦果长、宽、体积和千粒质量呈显著或极显著负相关,瘦果含水率与基质营养成分、重金属含量均有显著相关性,总的来说,过高的重金属含量会使瘦果变小、变轻,含水率下降。由此,可以初步判断剑叶金鸡菊主要通过瘦果大小和含水率等繁殖策略的改变及对重金属采取避性途径来响应尾矿库区重金属含量高、营养贫乏及持水能力差等基质条件,具有较强的环境适应能力。
关键词: 生境;剑叶金鸡菊;瘦果;种子;微形态;形态;品质
中图分类号: S682.1 文献标识码: A 文章编号: 1000-4440(2023)02-0530-09
Characteristics of achenes and seeds of Coreopsis lanceolata under different habitats
XU De-cong, WANG Yu, SHEN Zhang-jun, CHEN Yan-song
(School of Life Sciences, Hefei Normal University, Hefei 230601, China)
Abstract: To explore the effects of different habitats on achenes and seeds characteristics of Coreopsis lanceolata and reveal its environmental adaptation mechanism, the mature achenes of C. lanceolata were collected from three different habitats including slopes (I), Yangshanchong copper mine tailings pond (Ⅱ) and its soil covering reclamation area (Ⅲ) in Tongling. The morphological and quality indices of the achenes including the length, width, thickness, thousand-grain weight, specific gravity, and water content were determined. The contents of heavy metals, calcium and total phosphorus in the achenes were also measured. The microscopic morphology and internal structure of achenes and seeds of C. lanceolata were observed by biological stereoscopic microscope and scanning electron microscope (SEM). The results showed that the achenes and seeds collected from different habitats had similar morphological structures and could be distinguished from other species, whereas the accessory structure of achenes and the surface micromorphology of seeds were different among three habitats. The length, width, volume and thousand-grain weight of achenes collected from habitats Ⅰ and Ⅱ were both significantly higher than those of achenes collected from habitat Ⅲ (P<0.05), but there was no significant difference between habitats Ⅰ and Ⅱ. The water content of achenes in habitat Ⅰ was significantly higher than that in habitat Ⅲ, while this index in habitat Ⅲ was significantly higher than that in habitat Ⅱ. There were no significant differences in thickness and specific gravity of achenes among three habitats. The results of principal component analysis showed that the three habitats were divided into three groups, it was further proved that the morphology and quality of achenes were different among the three habitats. The accumulation of heavy metals in achenes did not show the same increasing trend as that in substrates, meanwhile the contents of calcium and total phosphorus in achenes in habitats Ⅱ and Ⅲ were significantly higher than those in habitat Ⅰ. The correlation analysis results showed that there were significantly and extremely significantly(P<0.01) negative correlations of the length, width, volume and thousand-grain weight of achenes with the contents of Cu and Zn in substrates, and the water content of achenes showed significant relationship with the contents of nutrient and heavy metals in substrates. They indicated that higher heavy metals contents in substrates could cause smaller, lighter, and lower water contents of achenes. In conclusion, C. lanceolata had a strong environmental adaptability in mine tailings, in that they could change reproductive strategies including achene size and water content, and keep their achenes free from heavy metals in response to substrate conditions including high heavy metals contents, poor nutrition, and poor water holding capacity.
Key words: habitats;Coreopsis lanceolata;achene;seed;micromorphology;morphology;quality
有色金属矿开采是全球环境污染的重要因素,会对周边生态系统造成严重破坏[1-3],例如大量含重金属尾矿砂堆积在露天区域(尾矿库),受数量和区域的限制无法直接清除这些尾矿[4],只能就地治理。采取单一[5]或综合治理方法[6]对尾矿库进行治理,其中植物修复具有投入成本低,環境友好,长期适用性和美观性的优势,获得广泛认可。然而,由于尾矿库具有极端的生境条件,如基质物理结构不良,持水保肥能力差,养分贫乏,重金属毒性强等特点,影响了植物的定居[7],植被恢复的难度较大。因此,凡是能忍受尾矿库恶劣条件的植物在退化矿区修复中都起着非常重要的作用[8]。
近年来,利用乡土物种进行植被重建被普遍认为是非常有效的手段[9-10]。成功的植被恢复需要充分了解乡土物种的各种适应性,如繁殖、生长、营养需求和功能适应性等,特别是已经自然定居在尾矿库的植物,因为尾矿库生境条件对大多数植物生长发育是有害的,会触发植物发生各种形态、生理和代谢的变化[11]。作为其生存策略的一部分,定居尾矿库的植物调整生殖性状,包括种子形态、休眠和发芽率等,其中种子的大小和质量是影响植物环境适合度和对环境胁迫适应性的最相关因子[12]。目前选择能够产生大量可存活种子的物种仍然是矿区植被恢复最适当和经济的方式[9],特别是在需要进行大规模植被恢复的地区。因此,了解尾矿库定居植物种子特性及其适应机制对矿区植被的恢复是必不可少的[13-14]。
剑叶金鸡菊(Coreopsis lanceolata L.)在铜陵杨山冲铜尾矿库及城市周边山坡均能生长发育并形成优势种群,作为菊科植物,具有极强的有性繁殖能力,分布范围较广,这为研究其对环境的繁殖适应策略提供了条件[15]。剑叶金鸡菊能适应恶劣的条件,而外表没有毒性症状,因此,猜想从这些植株上采集的瘦果和种子是否与非尾矿生境中的瘦果和种子呈现不同的特性,瘦果和种子是否帮助其逃避不利环境,确保植物的定居和更新。目前,国内外主要开展了剑叶金鸡菊栽培繁育技术及生理生态特性方面的研究[16-17]。对剑叶金鸡菊瘦果和种子的形态、品质特征以及异质环境下其特性的差异等方面还缺乏研究。本研究拟分别采集山坡上(生境Ⅰ)、铜陵市杨山冲尾矿库区(生境Ⅱ)和尾矿库覆土复垦区(生境Ⅲ)3种生境下的剑叶金鸡菊瘦果,比较分析不同生境下瘦果和种子的形态、品质特征及重金属吸收的差异,探讨其繁殖性状对异质生境的响应策略,以期为其在尾矿库生态修复中的合理利用提供实证参考。
1 材料与方法
1.1 试验材料
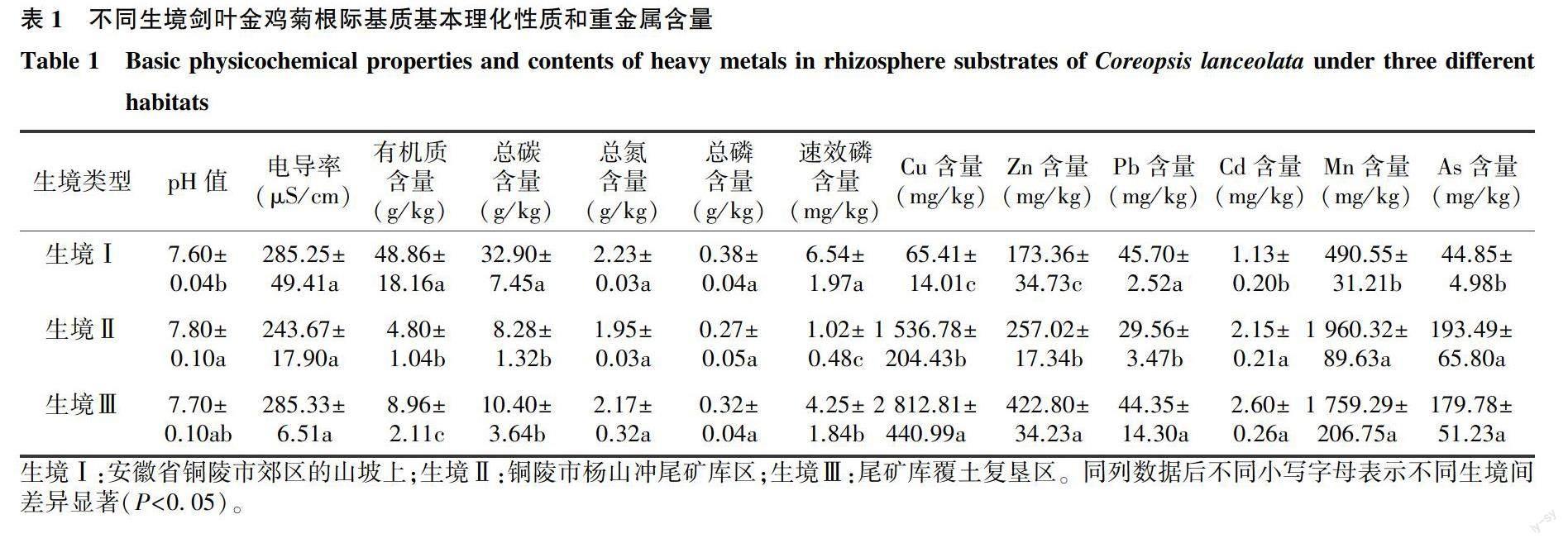
供试的剑叶金鸡菊瘦果分别采自安徽省铜陵市郊区的山坡上(生境Ⅰ)、铜陵市杨山冲尾矿库区(生境Ⅱ)和尾矿库覆土复垦区(生境Ⅲ),其中矿区概况见参考文献[15],山坡距离尾矿库约20 km。3种生境剑叶金鸡菊根际基质在有机质含量、总碳含量、速效磷含量、重金属含量上均出现显著差异(表1),测定方法和部分数据来自徐德聪等[15]的研究结果。剑叶金鸡菊在4月底至5月初开花,花期至10月中旬,于9月上旬随机选择瘦果大部分成熟的头状花序剪下,装入保鲜袋中带回实验室,晒干后置于干燥阴凉处,集中去杂,精选出瘦果。
1.2 试验方法
1.2.1 瘦果和种子外形特征及种皮显微结构观察 随机选择不同生境下干燥、饱满、成熟的剑叶金鸡菊瘦果和去果壳的种子,并在解剖镜下用解剖刀分别沿种子长轴和短轴切开,在生物体视显微镜下观察瘦果和种子的外观形态以及种子纵横切面结构并拍照。不同生境种子用导电胶粘贴于样品台上,采用ISC-20离子溅射仪喷金,通过SU-1510扫描电子显微镜观察种子表面纹饰,分别在×300和×500下进行拍照。参照文献[18]~[20]中的果实和种子外观形态和微形态学描述术语,对瘦果和种子的形状、表面特征等进行描述。
1.2.2 瘦果千粒质量、大小、密度及含水率测定 不同生境下瘦果千粒质量均用四分法随机抽样,用电子天平称其质量,每个生境设4个重复,计算瘦果平均千粒质量。每个生境随机选择15粒瘦果,用电子数显游标卡尺测量瘦果最长处、最宽处、最厚处,长、宽、厚的乘积代表种子体积,每个生境设4个重复,计算得到瘦果平均体积。瘦果含水率采用传统的烘干减质量法:每个生境分别随机选取100粒瘦果,用电子天平称质量,记录质量(m1),然后将其置于干净托盘中,在75 ℃烘箱干燥至恒质量,记录质量(m2),每个生境设3个重复,最后计算种子含水率。瘦果密度及含水率分别按以下公式计算:
瘦果密度=瘦果质量/瘦果体积(1)
瘦果含水率=(m1-m2)/m1×100%(2)
1.2.3 瘦果中重金属元素含量、钙含量、总磷含量测定 参照Xu等[21]的方法称取0.3 g清洗干净、烘干的瘦果,用硝酸-高氯酸湿法消解后,使用ICP-MS测定各重金属元素的含量,使用ICP-AES测定钙、总磷含量。各指标均重复测定3次,结果取平均值。
1.3 数据处理及统计分析
采用Excel2010和SSPS22.0进行数据处理与统计分析,数据以平均值±标准差的形式表示,对不同生境参数进行多重比较(LSD),依据瘦果形态与品质指标采用主成分分析对3种生境下的瘦果进行排序,并采用Person相关系数描述瘦果形态、品质指标与植物生长基质因子间的关系。
2 结果与分析
2.1 不同生境剑叶金鸡菊瘦果和种子的外观形态
3种生境下剑叶金鸡菊的成熟瘦果呈圆形或椭圆形,棕褐色,革质果壳,有金黄色或棕黄色宽翅,翅表面光滑,顶端有2个短鳞片,腹面上下两端有棒块状隆起(图1A、图1G、图1M)。凸起的瘦果背面粗糙且有纵棱(图1B、图1H、图1N)。瘦果腹面中部的颗粒状突起表现为生境Ⅰ比生境Ⅱ、生境Ⅲ多且排列紧密,生境Ⅲ的排列最为稀疏。种子呈卵形,浅黄色,略粗糙且微有光泽,有透明膜质种皮包裹,腹面和背面中央均有一条隆起的线纹,基部中央为种孔(图1C、图1D、图1I、图1J、图1O、图1P);种子的纵切面为新月形,质地为粉质,中间有明显的分层线,顶端为胚(图1E、图1K、图1Q),横切面为纺锤形,同样有明显的分层线(图1F、图1L、图1R),不同生境间无明显差异。
在扫描电子显微镜下,剑叶金鸡菊种子表面微形态复杂,种子较完整的种皮上可见长方形细胞规则纵向排列呈条状或网格状隆起的纹饰,但表面凸起分布不明显,隆起纹饰之间形成浅沟(图2A~图2F),其中生境Ⅲ种子表面还具明显的丝状连结物(图2C、图2F)。
2.2 不同生境剑叶金鸡菊瘦果大小、形态、品质特征比较
2.2.1 大小和形态特征 不同生境剑叶金鸡菊瘦果的平均长、宽、厚、长宽比和体积等大小和形态特征指标的变化如图3显示。除瘦果厚以外,其他指标在不同生境间均表现为生境Ⅲ与生境Ⅰ、生境Ⅱ有显著差异(P<0.05),但生境Ⅰ与生境Ⅱ之间差异均不显著(P>0.05)。其中,剑叶金鸡菊瘦果的平均长、宽和体积均表现为生境Ⅰ和生境Ⅱ显著大于生境Ⅲ;而长宽比表现为生境Ⅲ显著大于生境Ⅰ和生境Ⅱ。说明不同的基质环境可能是影响剑叶金鸡菊瘦果大小和形状的重要因素。
2.2.2 品质特征 图4显示,不同生境剑叶金鸡菊瘦果的千粒质量、含水率存在显著差异(P<0.05),但密度无显著差异(P>0.05)。剑叶金鸡菊瘦果的千粒质量表现为生境Ⅰ和生境Ⅱ显著大于生境Ⅲ,生境Ⅰ与生境Ⅱ之间无显著差异。剑叶金鸡菊瘦果的含水率表现为生境Ⅰ最大(11.76%),生境Ⅲ次之(6.98%),生境Ⅱ最小(5.01%),且三者之间均有显著差异(P<0.05)。同样说明不同的基质环境可能是影响剑叶金鸡菊瘦果品质特性的重要因素之一。
2.2.3 剑叶金鸡菊瘦果形态、品质指标的主成分分析 不同生境剑叶金鸡菊瘦果形态和品质指标的主成分分析因子载荷见表2。主成分1的特征值为5.792,主成分2的特征值为2.208,它们的累计方差贡献率达到100.000%,完全反映了所有指标的信息。主成分1的方差贡献率是72.396%,主要包含长、宽、厚、长宽比、体积和千粒质量这些信息;主成分2的方差贡献率是27.604%,主要包含了密度和含水率这2个信息。分别以主成分1和主成分2的得分作为横、纵坐标得到3个生境的瘦果排序图(图5),3个生境分为不同的区域,进一步证明生境间瘦果形态和品质出现了差异。
2.3 不同生境剑叶金鸡菊瘦果中重金属元素、Ca、总磷含量分析
表3显示,不同生境剑叶金鸡菊瘦果中重金属元素含量均表现为Zn含量最高,而Cd含量最低。除了生境Ⅰ中的Cd含量显著高于其他2个生境、生境Ⅲ中的Mn含量显著高于其他2个生境,其余重金属元素含量在不同生境间均无显著差异。说明剑叶金鸡菊瘦果对重金属的积累并没有随着基质中重金属含量的增加而增加,但瘦果中总磷含量在不同生境间均出现了显著差异,且生境Ⅱ和生境Ⅲ中瘦果钙含量均显著高于生境Ⅰ。
2.4 瘦果形态、品质指标与生长基质因子的相关性
表4显示,基质基本理化性质指标与瘦果大小(长、宽、厚、体积)相关性较弱,但基质中Cu、Zn含量均与瘦果长、宽、体积、千粒质量呈显著(P<0.05)或极显著(P<0.01)负相关。瘦果含水率与大部分基本理化指标(有机质含量、总碳含量、总磷含量和速效磷含量)均呈极显著正相关,但与Pb以外的基质中所有重金属的含量均呈负相关。可见,在本研究所有测定指标中,瘦果含水率受到较多基质生境因子的影响。
3 讨论
菊科作为被子植物中种类最多的一科,其瘦果、种子特性是形态分类学上的重要依据。通过对3种生境的剑叶金鸡菊瘦果和种子进行体视显微镜及扫描电镜观察,在《中国植物志》[20]关于剑叶金鸡菊瘦果基本形态特征只有形状、边缘有宽翅和顶端有2短鳞片等特点介绍基础上,增加了瘦果背面和腹面表面特征、果壳质地介绍内容,特别是腹面的颗粒状突起附属结构[22],是一个重要鉴别特征。《中国植物志》[20]中无剑叶金鸡菊种子特征的描述,本研究也做了重要的补充,如种子有透明种皮、腹面和背面中央均有1条隆起的线纹,顶端为种子的胚;种皮长方形细胞规则纵向排列、呈条状或网格状隆起的纹饰,隆起纹饰之间形成浅沟等特征[19],可将其作为区别于其他种的依据。
此外,本研究结果表明,基质中重金属含量高的生境Ⅲ中瘦果长、宽、体积和千粒质量都显著低于其他2个生境,这与Murawska-Wlodarczyk等[13]發现非金属生境的拟南芥种子平均比金属生境种子大18%的研究结果类似。Dresler等[23]关于蓝蓟不同居群种子大小的研究以及Babst-Kostecka等[24]关于孪果荠不同居群种子质量的研究中均有类似结论。说明金属超量积累植物和金属抗性植物的繁殖器官对基质中重金属含量的响应有相似性,瘦果和种子变小或质量变轻是对金属胁迫的一种忍耐机制。因为本研究的3种生境除了基质环境不同外,其他环境因子基本一致,尤其是生境Ⅲ的尾矿覆土往往会使重金属聚集在土层[15]。有趣的是,本研究中生境Ⅱ中重金属含量也较高,但瘦果大小(长、宽、厚、体积)与生境Ⅰ并无显著差异,这可能是因为生境Ⅲ中的金鸡菊植株是人工种植在覆土尾矿上,而生境Ⅱ中的所有金鸡菊植株是自然定居到尾矿库上的[15],自然定居植物的繁殖器官大小正常可能是对金属生境的一种适应。Pan等[25]认为当生境条件不利时,种子的保持可能优先于营养发育。Jiménez-Ambriz等[26]也提出金属污染点的植物营养生长受限,而种子会保持正常甚至大于非金属污染点。不同的结论可能与植物生长基质中金属含量是否对植物造成胁迫以及胁迫程度轻重有密切关系,而且植物自身的一种适应性保护机制也不可忽视。此外,Murawska-Wlodarczyk等[13]发现金属生境与非金属生境间的拟南芥种子长宽比没有显著差异,但本研究基质重金属含量高的生境Ⅲ中瘦果长宽比却显著大于其他2个生境,这可能是由物种自身对生境条件响应的差异所致。主成分分析结果也进一步证明不同生境间瘦果形态和品质出现了明显差异。
金屬生境、非金属生境种子中重金属积累状况因植物种类不同而不同。Dresler等[23]指出,铅锌尾矿生境和无污染土壤生境中的蓝蓟种子在大小和品质方面都存在差异,且铅锌尾矿生境中的种子Pb、Zn含量远高于无污染土壤生境中的种子。Babst-Kostecka等[24]关于孪果荠的研究结果表明,金属生境(高Zn含量)和非金属生境条件下种子中总Zn含量并无明显差异,只是金属生境种子胚乳中的Zn含量明显高于非金属生境。Babst-Kostecka等[27]发现,锌超富集植物拟南芥的种子在非金属生境中Zn含量最高,其他元素在不同生境中差异不明显。本研究3个生境基质中重金属含量有明显差异,尤其是Cu、Mn和As,但不同生境剑叶金鸡菊瘦果中均没有重金属的高度积累,且3个生境瘦果中Cu含量并没有显著差异,由此可以推断剑叶金鸡菊瘦果对重金属的抗性主要采取避性途径。一般认为,在含金属土壤中耐性植物种子会尽力避免吸收过量毒性金属[28]。植物对金属的避性机制有着多种途径,有研究结果表明,Ca的吸收与重金属离子的吸收和分布密切相关,Ca元素在植物体各种膜结构中通过调节膜透性控制物质交流转运[29]。有研究报道,金属生境中植物叶片[30-31]、种子[27]中的Ca含量均显著高于非金属生境。本研究结果也表明,矿区生境剑叶金鸡菊瘦果中Ca含量均显著高于山坡生境,且矿区生境瘦果中Cd含量显著低于山坡生境,结合Ca元素在植物体内有减弱Cd元素[31]和Zn元素[32]毒害作用的功能,可以认为Ca元素在剑叶金鸡菊瘦果内的升高是其具有重金属避性的一个可能途径,是其对极端生境的一种生理上的自我保护。此外,磷元素储存也受到Ca元素影响,而且磷对重金属元素吸收也有着重要影响[33],本研究结果表明剑叶金鸡菊瘦果磷吸收增加的同时也可能抑制了部分重金属元素的吸收。
相关性分析结果表明,基质中Cu、Zn含量与瘦果的长、宽、体积、千粒质量均呈不同程度的负相关,这进一步说明基质中重金属含量是瘦果大小及千粒质量变化的直接影响因子。此外,所测基质基本理化性质对瘦果大小和千粒质量影响较小,而对瘦果含水率有着明显影响,3种生境瘦果含水率呈现生境Ⅰ>生境Ⅲ>生境Ⅱ的变化规律,其中,基质有机质含量、总碳含量、总磷含量、速效磷含量均与瘦果含水率呈极显著正相关。钟军弟等[34]也发现假臭草种子含水率表现为高养分生境高于低养分生境。张文标等[35]的研究结果表明,光照条件是影响夏蜡梅果实、种子形态的最主要因子,但果实和种子大部分的表型性状与土壤中的速效磷含量、铵态氮含量有着一定程度的相关性,且种子厚主要受土壤电导率的影响。Balekoglu等[36]研究发现,6-8月最高气温的平均值与松果和种子的大小、质量呈显著正相关。说明不同生境间环境因子越复杂,与果实、种子表型性状存在相关性的因子会越多。本研究的生境因子相对单一且典型,这也可能是基质基本理化性质对瘦果大小和千粒质量影响较小的原因。此外,生境Ⅱ瘦果的含水率最低,可能还与铜尾矿主要是砂性质地,保肥持水能力差,尾砂分层不均匀,上部粒径稍粗,下部粒径较小且易被水力洗脱等密切相关[37]。瘦果含水率与基质中Pb以外的其他重金属含量均呈负相关,说明基质中重金属含量也是影响瘦果含水率的重要因素。不同生境剑叶金鸡菊瘦果后续种植在不同污染程度土壤上的生长、生理、繁殖特征均有待进一步研究。
4 结论
本文研究结果表明,3种不同生境下的剑叶金鸡菊瘦果外形基本一致,可区分于其他种,特别是腹面的颗粒状突起附属结构,可将其作为一个重要鉴别特征,但不同生境间突起附属结构有一定差异;不同生境间种子形态结构无明显差异,表面微形态结构有微小差异。基质中重金属含量是影响瘦果大小和千粒质量的主要因子,重金属含量过高会使瘦果长、宽、体积和千粒质量减小,而瘦果含水率受到基质营养条件和重金属含量的共同影响,且基质营养条件似乎对其影响更明显,说明瘦果含水率对环境因子响应更敏感。矿区生境中的剑叶金鸡菊瘦果对重金属的抗性采取避性途径,且瘦果中Ca含量、P含量对其吸收重金属起到一定抑制作用。由此,可以初步判断剑叶金鸡菊通过改变瘦果大小和含水率以及避免吸收过量重金属来响应尾矿库区重金属含量高、营养含量低及持水能力差等基质条件,特别是矿区剑叶金鸡菊瘦果中Ca含量的明显升高可能提高了其总体的抗逆性,使其具有较强的环境适应能力。
参考文献:
[1] IBRAHIMPOUR S, KHAVANINZADEH A R, TAGHIZADEHMEHRJARDI R, et al. Dustrelated impacts of mining operations on rangeland vegetation and soil: a case study in Yazd province, Iran[J]. Environmental Earth Sciences, 2021, 80(14): 467.
[2] JAHROMI M A, JAMSHIDI-ZANJANI A, DARBAN A K. Heavy metal pollution and human health risk assessment for exposure to surface soil of mining area: a comprehensive study[J]. Environmental Earth Sciences, 2020, 79(14): 365.
[3] MODOI O C, ROBA C, TOROK Z, et al. Environmental risks due to heavy metal pollution of water resulted from mining wastes in NW Romania[J]. Environmental Engineering and Management Journal, 2014, 13(9): 2325-2336.
[4] GONZ?LEZ-VALDEZ E, ALARC?N A, FERRERA-CERRATO R, et al. Seed germination and seedling growth of five plant species for assessing potential strategies to stabilizing or recovering metals from mine tailings[J].Water, Air, & Soil Pollution, 2016, 227(1): 24.
[5] LUO Z T, TANG C B, HAO Y H, et al. Solidification/stabilization of heavy metals and its efficiency in lead-zinc tailings using different chemical agents[J]. Environmental Technology, 2022, 43 (11): 1613-1623.
[6] NEJAD Z D, KIM J W, JUNG M C. Reclamation of arsenic contaminated soils around mining site using solidification/stabilization combined with revegetation[J]. Geosciences Journal, 2017, 21(3): 385-396.
[7] CROSS A T, IVANOV D, STEVENS J C, et al. Nitrogen limitation and calcifuge plant strategies constrain the establishment of native vegetation on magnetite mine tailings[J]. Plant and Soil, 2021, 461(1/2):181-201.
[8] FREITAS H, PRASAD M N V, PRATAS J. Plant community tolerant to trace elements growing on the degraded soils of S?o Domingos mine in the south east of Portugal: environmental implications[J]. Environmental International, 2004, 30 (1): 65-72.
[9] RAMOS S J, CALDEIRA C F, GASTAUER M, et al. Native leguminous plants for mineland revegetation in the eastern Amazon: seed characteristics and germination[J]. New Forests, 2019, 50(5):859-872.
[10]NEDUNURI K V, LOWELL C, MEADE W, et al. Management practices and phytoremediation by native grasses[J].International Journal of Phytoremediation,2010,12(2):200-214.
[11]CARVALHO M E A, GAZIOLA S A, CARVALHO L A, et al. Cadmium effects on plant reproductive organs: physiological, productive, evolutionary and ecological aspects[J]. Annals of Applied Biology, 2021,178(2):227-243.
[12]WANG Y P, WANG J F, GUO S G, et al. CRISPR/Cas9-mediated mutagenesis of C1BG1 decreased seed size and promoted seed germination in watermelon[J]. Horticulture Research, 2021,8:70.
[13]MURAWSKA-WLODARCZYK K, KORZENIAK U, CHLEBICKI A, et al. Metalliferous habitats and seed microbes affect the seed morphology and reproductive strategy of Arabidopsis halleri[J]. Plant and Soil, 2022,472(1/2):175-192.
[14]S?NCHEZ-L?PEZ A S, THIJS S, BECKERS B, et al. Community structure and diversity of endophytic bacteria in seeds of three consecutive generations of Crotalaria pumila growing on metal mine residues[J]. Plant and Soil, 2018, 422(1/2):51-66.
[15]徐德聰,孙庆业,沈章军,等. 铜尾矿库剑叶金鸡菊根际尾矿和植株的重金属元素含量及相关性分析[J]. 植物资源与环境学报, 2018, 27(1): 27-36.
[16]许 媛,孙 进,王 军,等. 剑叶金鸡菊对植物群落结构的影响[J]. 海洋湖沼通报, 2009(2): 73-78.
[17]SAITO T I, OKUBO K. Influences of invasive herb Coreopsis lanceolata on riparian endemic herbs in relation to the understory light availability[J]. Landscape and Ecological Engineering, 2013, 9(2): 271-280.
[18]刘长江,林 祁,贺建秀. 中国植物种子形态学研究方法和术语[J]. 西北植物学报, 2004, 24(1): 178-188.
[19]王利丽,杨 珂,郭雨轩,等. 果实种子类药材微形态的鉴别研究[J]. 电子显微学报,2020, 39(3):313-322.
[20]中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1979:365.
[21]XU D C, SHEN Z J, DOU C M, et al. Effects of soil properties on heavy metal bioavailability and accumulation in crop grains under different farmland use patterns[J]. Scientifc Reports, 2022,12(1): 9211-9226.
[22]杜珍珠. 石河子綠洲—荒漠区菊科植物及种子形态学研究[D]. 石河子:石河子大学,2013.
[23]DRESLER S, BEDNAREK W, HAWRYLAK-NOWAK B, et al. Morphometric and phytochemical profile of seeds of metallicolous and nonmetallicolous Echium vulgare populations[J]. Biochemical Systematics and Ecology, 2017, 70:304-310.
[24]BABST-KOSTECKA A, PRZYBY?OWICZ W J, VAN DER ENT A, et al. Endosperm prevents toxic amounts of Zn from accumulating in the seed embryo-an adaptation to metalliferous sites in metal- tolerant Biscutella laevigata[J]. Metallomics,2020, 12(1):42-53.
[25]PAN G, ZHANG H, LIU P, et al. Effects of manganese stress on phenology and biomass allocation in Xanthium strumarium from metalliferous and non-metalliferous sites[J]. Ecotoxicology and Environmental Safety, 2019, 172:308-316.
[26]JIM?NEZ-AMBRIZ G, PETIT C, BOURRI? I, et al. Life history variation in the heavy metal tolerant plant Thlaspi caerulescens growing in a network of contaminated and noncontaminated sites in southern France: role of gene flow, selection and phenotypic plasticity[J]. New Phytologist, 2007, 173:199-215.
[27]BABST-KOSTECKA A, PRZYBY?OWICZ W J, SEGET B, et al. Zinc allocation to and within Arabidopsis halleri seeds: diferent strategies of metal homeostasis in accessions under divergent selection pressure[J]. Plant-Environment Interactions. 2020, 1(3):207-220.
[28]BOTHE H, S?OMKA A. Divergent biology of facultative heavy metal plants[J]. Journal of Plant Physiology, 2017, 219: 45-61.
[29]蒋廷惠,占新华,徐阳春,等. 钙对植物抗逆能力的影响及其生态学意义[J]. 应用生态学报, 2005, 16(5):971-976.
[30]TURNAU K, OSTACHOWICZ B, WOJTCZAK G, et al. Metal uptake by xerothermic plants introduced into Zn-Pb industrial wastes[J]. Plant Soil, 2010, 337(1/2):299-311.
[31]PROBST A, LIU H Y, FANJUL M, et al. Response of Vicia faba L. to metal toxicity on mine tailing substrate: geochemical and morphological changes in leaf and root[J]. Environment and Experimentl Botany, 2009,66(2):297-308.
[32]TODESCHINI V, LINGUA G, DAGOSTINO G, et al. Effects of high zinc concentration on poplar leaves: a morphological and biochemical study[J]. Environment and Experimentl Botany, 2011,71(1): 50-56.
[33]刘登彪,郑骏宇,赵 涛,等. 不同供磷水平对四种超富集植物生长及吸收重金属的影响[J]. 环境科学学报, 2015, 35(4):1198-1204.
[34]钟军弟,陈 燕,刘锴栋,等. 不同生境下假臭草的种子特性分析[J]. 江苏农业科学, 2017, 45(9): 87-91.
[35]张文标,金则新. 夏蜡梅果实和种子形态变异及其与环境因子相关性[J]. 浙江大学学报(理学版), 2007, 34(6):689-695.
[36]BALEKOGLU S, CALISKAN S, DIRIK H. Effects of geoclimatic factors on the variability in Pinus pinea cone, seed, and seedling traits in Turkey native habitats[J]. Ecological Processes, 2020, 9:55.
[37]周元祥. 安徽铜陵典型尾矿库地球化学和环境地球化学效应[D]. 合肥:合肥工业大学, 2009.
(责任编辑:王 妮)
