巨噬细胞与肿瘤治疗的研究进展
2023-05-30张玉民综述刘金剑刘鉴峰审校
王 丹,张玉民(综述),刘金剑,刘鉴峰(审校)
(中国医学科学院放射医学研究所放射医学与分子核医学重点实验室,天津 300192)
肿瘤发生不仅取决于癌细胞的内在特性,还取决于癌细胞与肿瘤微环境(tumor microenvironment,TME)成分的相互作用。巨噬细胞是TME中最丰富的免疫细胞之一,通常具有2种极化状态,可响应不同的刺激而分化为M1型和M2型[1]。在肿瘤发展的初始阶段,巨噬细胞可以通过杀死肿瘤细胞直接促进抗肿瘤反应。由于肿瘤或Th2细胞开始主导TME,肿瘤相关巨噬细胞(tumor-associatd macrophage,TAM)开始表现出免疫抑制的M2型,通过产生大量生长因子、细胞外基质重塑分子和细胞因子调节肿瘤的生长、迁移和血管生成[2]。已有许多研究表明M2表型的巨噬细胞与肝癌、结肠癌、胰腺癌、甲状腺癌和脑肿瘤等有关[3],这表明M1-M2型巨噬细胞极化可能在癌症治疗中发挥一定的作用。
1 巨噬细胞的极化分型
巨噬细胞具有很强的可塑性,并具有功能多样性。最初认为巨噬细胞表现为M1表型参与抗肿瘤免疫,但在恶性肿瘤进展过程中显示其可以促进癌症的发生,刺激血管生成和抑制抗肿瘤免疫,这是由于巨噬细胞已经表现成M2表型所导致的[4]。巨噬细胞的表型可以通过TME中的一些因素例如调节性T细胞产生的免疫抑制细胞因子、趋化因子、肿瘤细胞产物来调控,也可以通过Th1和Th2辅助细胞的细胞因子库改变。微生物刺激如脂多糖和Th1相关细胞因子如干扰素γ(interferon-γ,IFN-γ)可以将巨噬细胞极化为M1表型[5],并在促炎、杀死微生物和肿瘤抵抗过程中发挥一定的作用。M1型巨噬细胞有以下特征:包括抗原呈递能力,高产量的白介素(interleukin,IL)-6、IL-12和IL-23,高产量的有毒中间体包括一氧化氮(nitric oxide,NO)、活性氧中间体和基质金属蛋白酶12,如图1所示[6]。研究显示,在乳腺癌小鼠模型中,唑来膦酸修饰的纳米颗粒有效抑制了破骨细胞形成,并同时诱导巨噬细胞向M1促炎表型极化[7]。Th2细胞因子如IL-4和IL-13可以使巨噬细胞极化为M2表型,M2型巨噬细胞在抗炎、组织修复重塑、寄生虫清除、促进肿瘤生长和免疫调节过程中起作用。Haydar等[8]研究显示阿奇霉素通过抑制STAT1和核转录因子(nuclear factor kappa B,NF-κB)信号通路使巨噬细胞极化为M2型,从而有效控制囊性纤维化患者的过度炎症。根据诱导信号极化的方式,M2型巨噬细胞又可分为以下4种:M2a、M2b、M2c和M2d型。其中M2a型是由IL-4或IL-13诱导的,通过分泌细胞外基质促进组织修复;M2b型是通过暴露于免疫复合物和Toll样受体(Toll-like receptors,TLRs)或IL-1受体的激动剂参与抗炎反应和免疫调节功能诱导的[9];M2c型由糖皮质激素和IL-10诱导抑制免疫反应和组织重塑。受TME影响的TAM在肿瘤的生长、侵袭和转移中起着至关重要的作用。TAM是一种替代活化的M2型巨噬细胞。一些TAM看起来类似于M2b表型(IL-10高,IL-12低),而另一些在鼠类和人类肿瘤研究中已显示其肿瘤坏死因子α(tumour necrosis factor-α,TNF-α)低,表型类似于M2c表型。但是,一些学者将TAM归类为M2d巨噬细胞,其表达高水平的血管内皮生长因子和IL-10。由腺苷引起的M2d型巨噬细胞中,白血病抑制因子和IL-6被认为在诱导血管生成、调节肿瘤进展和增强肿瘤的存活方面发挥关键作用(图2)。
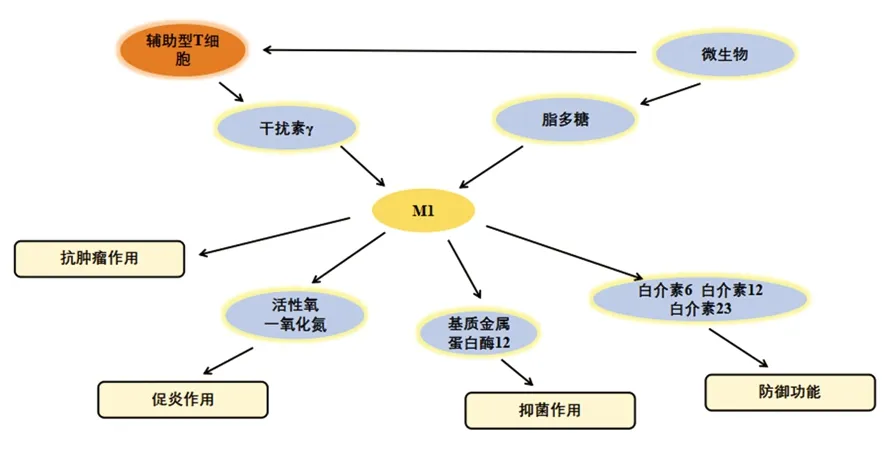
图1 M1型巨噬细胞的激活因子和功能
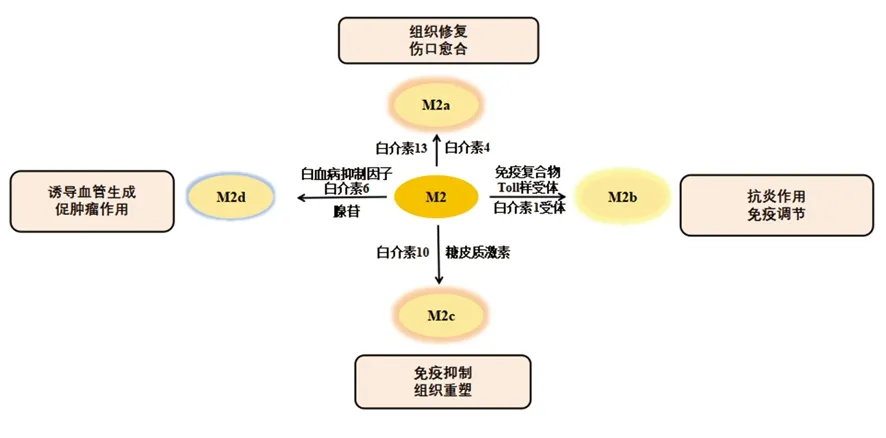
图2 M2型巨噬细胞的激活因子和功能
2 巨噬细胞的表面标志物-细胞因子和趋化因子
循环单核细胞和组织巨噬细胞存在不同的受体和独特的分泌模式见表1[6]。M1型巨噬细胞高度表达主要组织相容性复合体(major histocompatibility complex,MHC)Ⅱ、IL-1R、2、TLR4、CD80、CD86和其他刺激性分子。同时,M1型巨噬细胞分泌促炎细胞因子如TNF-α和IL-1,以及一些趋化因子如CCL2、CCL3、CCL5、CXCL8、CXCL9、CXCL10、CXCL11和CXCL16,会产生高水平的重要炎症因子,包括IL-23、IL-6、IL-12[4]。M1型巨噬细胞也与活性氧(reactive oxygen species,ROS)的合成和NO的释放有关。褐藻糖苷作为一种抗氧化剂,可抑制单核细胞/巨噬细胞和侵袭性癌细胞中的细胞内ROS和线粒体超氧化物水平。研究显示,与ROS抑制剂相比,褐藻糖苷直接诱导单核细胞向M1型巨噬细胞极化,并将M2巨噬细胞重新极化为M1表型[10]。NO也在M1型巨噬细胞中扮演重要的角色。研究显示,M1表型TAM负责诱导性一氧化氮合成酶的表达并催化NO的合成,释放的NO进入TME通过作用于M1型巨噬细胞在肿瘤抑制中发挥作用[11]。此外,高通量的NO到肿瘤基质也刺激TAM向M1表型转换[12],从而协调基质内的抗肿瘤信号。
M2型巨噬细胞也表达许多MHC-Ⅱ分子,但是这种表达不足以有效地呈递抗原;其还表达高水平的精氨酸酶1,可以促进多胺的合成并刺激组织修复、细胞生长、胶原蛋白形成等。其可细分为M2a、M2b、M2c和M2d型。M2a型巨噬细胞除了产生CCL17、CCL18、CCL22和CCL24外,还表达高水平的表面分子和受体如CD163、CD23、CD209、Fizz1、Arg1、Ym1/2、IL-4R、FcR、CXCR1、CXCR2和Dectin-1等。M2b型巨噬细胞表达高水平的表面分子CD80和CD86,并产生TNF-α、CCL1、IL-1、IL-6和IL-10。M2c型巨噬细胞表达高水平的表面受体和分子,包括CD14、CD50、MR和SR,并产生IL-10、CCL16、CCL18、CXCL13和转化生长因子β(transforming growth factor-β,TGF-β)。M2d型巨噬细胞(TAM)表达高水平的血管内皮生长因子和CD163,产生细胞因子如IL-10、IL-12、TNF-α和TGF-β,并分泌趋化因子CCL5、CXCL10、CXCL16、CCL18。

表1 巨噬细胞的表面标志物、细胞因子和趋化因子
3 M1-M2极化的分子途径
巨噬细胞的极化受转录因子和miRNA驱动的几种机制的影响如图3。
3.1干扰素调节因子(interferon regulatory factor,IRF)/转录激活因子(signal transducer and activator of transcription,STAT)信号通路 巨噬细胞向M1或M2表型极化主要是由IRF/STAT信号通路诱导的。其中IRF3、IRF5、STAT1和STAT5负责驱动M1极化,而IRF4、STAT3和STAT6则提供M2表型的激活信号。IFN-γ是一种有效的内源性巨噬细胞激活因子,可以主要激活STAT1,并通过IFN-γ/JAK/STAT1通路发出信号诱导M1样巨噬细胞极化[13]。除了STAT1信号通路,有研究显示,通过阻止CrkL-STAT5复合物的形成,可以减轻M1表型转换和巨噬细胞炎症[14]。Th2细胞因子如IL-13和IL-4,其启动子受信号转导和STAT6调节,通过诱导过氧化物酶体增殖物激活受体表达在巨噬细胞中产生M2样激活[15]。Wu等[16]研究显示玛格毒素通过抑制p-STAT1活性调节M1标志物的表达,并通过促进p-STAT6活性来增加M2标志物的表达。此外,IRF1、IRF5和IRF-8与M1表型极化相关,而IRF3和IRF4参与M2巨噬细胞的极化过程[17]。
3.2低氧诱导因子(hypoxia inducible factor,HIF)信号通路 除了IRF/STAT转录因子外,缺氧也会影响巨噬细胞的极化。缺氧能以不同的方式激活M1和M2巨噬细胞中的HIF。Th1细胞因子能够诱导HIF-1α表达,引发M1应答;而Th2细胞因子在M2巨噬细胞中会活化HIF-2α。这些差异取决于HIF-1和HIF-2分别激活或抑制NO合成的能力。最近的一项研究表明,从低氧肿瘤区域释放的外泌体而不是从常氧区域释放的外泌体能促进M2型巨噬细胞极化[18]。具有抗肿瘤活性的M1型巨噬细胞停留在肿瘤中的显示,表明可开发一种新颖的利用TAM对抗癌症的治疗方法。
3.3NF-κB信号通路 NF-κB在促炎性巨噬细胞扮演着重要角色。事实上,活化的异质二聚体NF-κB(p50-p65),促进了抗炎性巨噬细胞中炎症基因的转录;而抑制异质二聚体NF-κB(p50-p50)则阻止了这些基因的转录。Liu等[19]研究显示颗粒蛋白前体通过NF-кB和MAPK通路抑制LPS诱导的巨噬细胞M1极化。其他转录因子包括转录因子激活蛋白1、Kruppel样因子4和过氧化物酶体增殖物激活受体也可以调节巨噬细胞的活化状态。
3.4miRNA信号通路 miRNA也会干扰巨噬细胞的极化。miR127、miR155和miR223是M1极化的关键调控因子,而miR146a则促进了M2极化,降低了M1标记的表达。Wang等[20]研究显示miR-99b的靶向递送可重新编程与肿瘤相关的巨噬细胞表型,从而导致肿瘤消退。由于上述通路有时会相互作用,因此研究与M1和M2表型相关的巨噬细胞极化的信号通路在理解和创造预防肿瘤发展的治疗方法方面起着重要的作用。
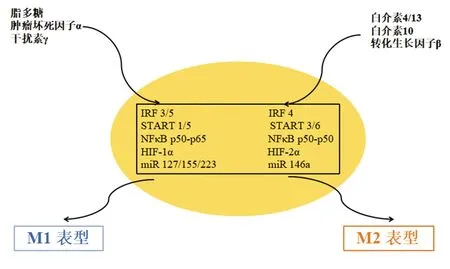
图3 巨噬细胞向M1和M2极化的分子途径
4 TAM与癌症
TAM是肿瘤微环境中主要的浸润性白细胞,并且在炎症与癌症之间发挥关键作用。在癌症初期,TAM可能发挥强大的免疫活性;但在后期,TME通过富集生长因子和抗炎介质,使TAM向促进肿瘤生长的M2型极化。
TAM影响肿瘤进展的不同方面。首先,其可以调节细胞外基质降解的酶和蛋白酶如基质金属蛋白酶、纤溶酶、骨连接蛋白和组织蛋白酶,TAM对细胞外基质的破坏促进了肿瘤细胞的扩散和转移。其次,TAM也导致肿瘤内的免疫抑制环境,TAM通过抑制抗原呈递细胞,使其无法分泌IL-12,并且产生高水平的IL-10和TGF-β,从而阻断T细胞增殖,抑制细胞毒性T淋巴细胞(cytotoxic T lymphocyte,CTL)反应并激活Treg[21]。作为对微生物信号、组织损伤、细胞因子和代谢产物的响应,组织中的单核细胞和巨噬细胞能够进行重新编程,实现M1与M2表型之间的转化。TAM促进肿瘤生长的性质及其从M2型巨噬细胞向M1表型转变的能力,使其成为一个有吸引力的抗癌治疗靶点。通过靶向TAM,许多研究在不同的肿瘤模型中均取得了良好的抗肿瘤作用。目前,放射疗法、化学疗法和免疫疗法仍然是临床上肿瘤治疗的重要方式。然而,由于治疗耐药性或放疗和化疗过程中的不良反应,一些接受治疗的患者仍然患有肿瘤转移或局部复发性疾病。因此,靶向TAM的药物与其他癌症治疗方法相结合的策略将提供一种有希望的手段。
4.1靶向TAM的化学疗法 虽然化疗药物的开发主要是为了诱导快速分裂的癌细胞死亡,但许多药物对非癌细胞群体也有药理作用。有研究表明氯膦酸盐脂质体耗尽巨噬细胞可阻止骨髓瘤的发展[22],其发生机制为含有氯膦酸盐的脂质体被巨噬细胞吞噬后,经过溶酶体处理破坏氯膦酸盐在细胞中释放,随后氯膦酸盐代谢成类似于ATP的物质对巨噬细胞产生细胞毒性进而抑制骨髓瘤的发展。
除了发挥细胞毒作用外,几种化疗药物还能调节巨噬细胞极化的表型。2个主要的转录因子(STAT3和STAT6)已大量报道参与阻断M2表型。白藜芦醇通过抑制STAT3通路实现对TAM向M2型极化的抑制,进而成功的抑制了小鼠肺癌异种移植模型中的肿瘤生长,其阻断TAM的M2样极化与STAT3活性降低有关[23]。类似地,芬维A胺抑制STAT6的磷酸化实现对TAM向M2极化的有效抑制,在结肠直肠小鼠模型中,这种效应伴随着肿瘤中M2样巨噬细胞数量的减少和血管生成的抑制[24]。此外,其他STAT6激活的抑制剂(TMC264,AS1517499)也被开发出来,但尚未进行临床研究。紫杉醇作为一种常用的抗肿瘤化疗药物,有研究显示其通过将与肿瘤相关的巨噬细胞重编程为TLR4依赖性的M1型,从而抑制了肿瘤的生长[25]。为了对抗这些耐药机制,化疗和免疫治疗的结合可能会使肿瘤消退更有效。值得注意的是,紫杉醇和吉西他滨是2种化疗药物,其可能诱导免疫原性死亡,通常与其他TAM调节治疗药物联合治疗。
4.2靶向TAM的放射疗法 放射疗法是一种主要用于恶性肿瘤的局部治疗形式。许多研究显示放射疗法会改变巨噬细胞的表型,这为癌症的治疗提供了一个新的思路。Stary等[26]研究显示直肠癌患者的短程放疗与TAM向M1样促炎表型的转变有关。但是,关于放疗对巨噬细胞表型的影响有矛盾的结果。有研究表明,肿瘤照射与TAM的免疫抑制表型有关[27]。Seifert等[28]研究显示与未放射的小鼠相比,来自放射治疗的浸润性和浸润前胰腺肿瘤的巨噬细胞比例更大,具有免疫抑制性的M2样表型。这提示放疗的某些方面会影响巨噬细胞的表型。因此,为了分析放疗对巨噬细胞重编程的影响,Genard等[29]根据放射剂量进行了分类(低剂量为<1 Gy的剂量,中剂量为1~10 Gy的剂量,高剂量为>10 Gy的剂量),显示高剂量和低剂量对巨噬细胞从M2向M1的极化无影响,而中剂量明显引起TAM重编程。同时在进行全身放疗的实验中显示:健康小鼠在接受低剂量或中剂量的全身辐照后,巨噬细胞均趋向M1表型。显然,辐照的效果似乎取决于肿瘤模型,辐照剂量和组织等多种因素。
至于不同放射剂量导致TAMs的表型不同,是由于不同剂量的放疗后,p50-p50和p65-p502个二聚体组成的NF-κB平衡状态的不同[29]。照射剂量对巨噬细胞NF-κB平衡的影响相反:低剂量或1Gy以下剂量均不改变照射后巨噬细胞核p50-p50 NF-κB的丰度。中剂量或1-10 Gy剂量诱导NF-κB平衡从非活性同源二聚体(p50-p50)切换到活性异源二聚体(p50-p65),一旦p50-p65被转录到核内,促炎因子(TNF-α、IL-10、IL-6)等就会高表达,进而使巨噬细胞表现为M1表型。高剂量或高于10 Gy的剂量不能改变NF-κB的平衡,使得TAM倾向于M2表型。
基于放疗对TAM的极化影响,山东师范大学唐波教授课题组开发了X射线诱导表型转化策略[30],将工程化修饰的、M0代的巨噬细胞在体外经X射线照射之后,改造成具有激活肿瘤免疫的M1型TAMs。体内外试验证实了X射线诱导表型转化策略能够将巨噬细胞改造成M1型,进而显著抑制小鼠肿瘤的生长。因此,巨噬细胞极化策略将有助于增强临床上放射治疗或放疗联合免疫治疗的抗肿瘤效率。
4.3靶向TAM的免疫疗法 在过去的几年中,已经开发了多种肿瘤学免疫策略来重新激活适应性免疫和先天免疫系统,以建立强大的抗肿瘤免疫应答。作为经典抗癌治疗的替代方法,肿瘤通常会对此产生抗药性。免疫检查点抑制剂——针对细胞毒性T淋巴细胞抗原4的单克隆抗体,可增强细胞毒性CD8+T细胞活性的程序性细胞死亡受体1和程序性死亡受体-配体1的临床试验已显示成功地治疗了黑色素瘤和肺癌。但是,在大多数情况下,出于未知原因只有一小部分患者对免疫疗法完全有效。
目前已经显示了几种常用于将TAM从M2型重新编程为M1表型的药物,主要包括TLR激动剂、靶向M1表型的抑制蛋白单克隆抗体以及其他化合物。TLR激动剂是一种很有前途的抗肿瘤药物,已有研究显示包裹CpG寡脱氧核苷酸的靶向铁蛋白纳米颗粒可以诱导TAM从M2表型极化为M1表型,并抑制肿瘤生长[31]。此外,研究显示R848能够通过巨噬细胞极化下调肿瘤免疫抑制微环境中髓源抑制性细胞的表达,进而增强了联合治疗中奥沙利铂的抗肿瘤作用[32]。
另一种有利于TAM细胞毒功能的方法是用单克隆抗体刺激CD40。抗CD40单抗可以促进巨噬细胞的抗肿瘤作用,特别是通过加强NO和TNF-α的分泌,因此可以诱导CD8+T细胞抑制肿瘤的生长和转移。抗CD40单抗可在TAM和肿瘤浸润性单核细胞质膜中诱导PD-L1上调,同时有研究显示PD-L1轴的阻断与抗CD40和抗CTLA-4 单抗在结肠癌和乳腺癌模型中显示出广泛的存活率[33]。
重编程TAM的第三个替代方法是使用不同的化合物。最著名的一种是FDA批准的IFN-γ。研究显示白藜芦醇类似物的治疗显着增加了脾细胞中IFN-γ分泌细胞的表达并减少了M2型巨噬细胞的浸润,诱导的IFN-γ逆转了TAM的特性,促使TAM重编程为M1型[34]。不同的纳米粒子(nanoparticles,NPs)如金属NPs、氧化物NPs、质粒DNA NPs等,可以诱导M0巨噬细胞向各种表型极化并调节巨噬细胞重编程。Chattopadhyay等[35]研究显示抗原键合修饰的氧化钴NPs通过调节NADPH氧化酶和p38MAPK将TAM重编程为促炎表型。除此之外,还显示了一种小的类黄酮化合物DMXAA,既是血管破坏剂,也是干扰素基因刺激因子的鼠激动剂。可以使M2表型的巨噬细胞重新极化,从而发挥肿瘤的免疫治疗[36]。其发挥作用的机制是利用DMXAA激活干扰素基因刺激因子成为二聚化的结构,随后进一步与环鸟腺苷酸小分子结合,构象发生改变,招募TANK结合激酶1蛋白,促使其磷酸化IRF3,磷酸化的IRF3入核,诱导I型干扰素的产生,进而调控巨噬细胞向M1型极化。因此,利用TAM重编程的策略调控免疫治疗会在一定程度上更有效地抑制肿瘤生长和转移。
5 结论与展望
越来越多的研究已经显示巨噬细胞极化在癌症的发生发展中发挥重要作用。由于化学疗法、免疫疗法和放射疗法的局限性,利用TAM重编程的能力将靶向TAM与这三种疗法结合将为进一步提升癌症治疗的精准性提供可能。TAM重编程或者靶向TAM治疗是通过哪些机制实现对化学疗法、免疫疗法和放射疗法抗肿瘤效率的提升,需要具体的研究以期为癌症的临床治疗提供更有效的治疗思路和方法。
