紫金山南麓枫香种群结构与动态特征
2023-05-29邹朋峻关庆伟袁在翔谷雨晴牛莹莹金雪梅
邹朋峻,关庆伟*,袁在翔,谷雨晴,吴 茜,牛莹莹,陈 霞,金雪梅
(1. 南京林业大学生物与环境学院,南方现代林业协同创新中心,江苏 南京 210037;2.中山陵园管理局,江苏 南京 210014)
种群数量动态是种群个体长期适应生境的结果,也是生态学中关于种群研究的热点问题[1-3]。它不仅反映物种与生境之间的契合度[4],也反映了该物种在群落中的地位与作用[5-7]。静态生命表是研究植物种群数量动态的基本方法[8],能够反映植物种群在特定时间段内的死亡率和存活率,进而估算该种群在一定生境条件下的内禀增长率,并结合时间序列模型推测种群数量未来的变化趋势[9-12]。研究地带性优势种群的数量动态特征对于了解典型森林群落的稳定性和演替动态具有重要意义[13-16]。
枫香(Liquidambarformosana)是我国北亚热带及部分暖温带森林的上层先锋树种[17]。作为秋季彩林景观的重要组成部分,紫金山南麓枫香林的年龄约59 a,受人为干扰程度较低,林分郁闭度较高,优势种群径阶结构完整,研究其种群结构与数量动态具有重要理论价值和实践意义[18]。然而,以往关于枫香种群的研究主要为干扰条件下(如采伐)的种群结构、分布格局及天然更新特征。例如,陈水永[19]和王丹[20]的研究表明,在人为干扰严重的生境中,枫香苗木个体数量最多,且主要聚集分布在下坡位。李冬林等[17]在连云港市枫树湾的研究发现,在生物入侵和人为活动的双重干扰下枫香种群径级结构不完整,龄级结构呈偏正态分布,苗木总体较少,种群趋于衰退。王传华等[21]研究表明,枫香幼苗萌芽更新是对生境的适应,伐桩萌芽更新是对砍伐干扰的响应,其中萌芽更新在林木生长过程中存在显著的自疏现象。当前,对于森林群落长期自然演替下的枫香种群结构与数量动态特征研究依旧较为缺乏。仅少量研究表明,在中亚热带常绿阔叶林自然保护区中,枫香种群径级结构完整,幼苗个体较少,天然更新较为困难,种群呈现出前期薄弱、中期稳定、后期衰退的特征[22]。而该种群特征在北亚热带至暖温带过渡带的地带性植被针阔混交林中是否同样适用,需要做进一步实证研究。
为此,本研究以南京紫金山南麓样地中的枫香种群为研究对象,利用植物种群静态生命表、结构动态指数、时间序列模型等方法分析和预测该种群的结构特征及未来变化趋势,以期阐明自然演替条件下枫香种群的数量动态特征,为该地区森林的可持续经营提供重要的参考依据。
1 材料与方法
1.1 研究地概况及试验设计
南京紫金山位于南京市城东(118°48′24″~118°53′04″E,32°01′57″~32°06′15″N),总面积2 970 hm2,海拔最高处448.9 m,相对高度420 m,属于北亚热带向暖温带过渡的季风气候区,四季分明,年降水量在1 000~1 050 mm,年均气温15.4 ℃,年均日照2 213 h,全年无霜期322 d[23]。土壤为酸性山地黄棕壤和灰棕壤。森林资源清查数据表明,紫金山针叶林约516.71 hm2,阔叶林约1 752.50 hm2,针阔混交林约14.01 hm2,其他林分约118.04 hm2[24]。
在紫金山南麓针阔混交林(118°51′56″E,32°03′32″N)中受人为干扰较小处,用全站仪和罗盘仪设定固定样地1.36 hm2。在相邻格子法划分的100个10 m×10 m调查样方中,记录每株胸径≥5 cm林木的植物种名、胸径、树高、冠幅、坐标距离位置。在每个调查样方的4个边角及中心设置2 m×2 m调查小样方,在调查小样方内记录更新幼树、灌木、草本和藤本植物的种名、坐标距离位置、基径、株高、株数(或丛数)、盖度。
1.2 试验指标测定
1.2.1 乔木树种重要值
针阔混交林中每个树种的优势度以重要值大小来衡量,相对密度、相对频度、相对显著度及重要值的计算方法详见文献[2]。
1.2.2 种群个体径阶与龄级划分
由于缺乏江苏省枫香树的解析木资料,并且利用生长锥获取林木年轮较为困难,因而无法精确测定紫金山南麓枫香种群个体的真实年龄。但是,林木胸径与年龄存在一定的正相关关系,故采用径级来对应种群个体的龄级。枫香种群的径阶划分主要参考熊文愈等[25]对紫金山森林乔木大小等级的划分方法,并在此基础上稍做改动:在原方法上Ⅰ、Ⅱ级苗木与Ⅲ级立木合并为胸径为0~7.5 cm的幼苗幼树,定义为第Ⅰ径级,之后每径阶间隔5 cm;第Ⅰ径级对应龄级Ⅰ,第Ⅱ径级对应龄级Ⅱ,依此类推直至龄级Ⅸ。按照径阶统计种群各龄级株数。
1.2.3 种群结构动态指数

1.2.4 静态生命表与生存分析函数
参见文献 [27]对种群各龄级个体数编制静态生命表,包括种群每一龄级的存活率、死亡率、消失率等指标。引用生存率S(t)、累计死亡率F(t)、死亡密度f(t)、危险率λ(t)等4个生存分析函数,具体算法参见文献[28]。
此外,以龄级为横坐标,以标准化存活个体数的自然对数lnlx为纵坐标,绘制的种群个体存活曲线,可归为3种类型:Deevey-Ⅰ凸线型,该类型的种群在接近生理寿命前只有少数个体死亡;Deevey-Ⅱ直线型,该类型的种群在各龄级死亡率相等;Deevey-Ⅲ凹线型,该类型的种群在苗木期死亡率很高[1, 29]。此外,Deevey-Ⅱ直线型又被细分为3个亚型,其中B1亚型指种群在各龄级的存活个体数(lx)相差较大;B2亚型指种群各龄级的死亡率(qx)基本相同;B3亚型指种群在苗木期的死亡率较高,而后死亡率则明显下降。采用函数(Deevey-Ⅱ与Deevey-Ⅲ)模型对存活曲线进行拟合检验[27, 30]。
1.2.5 时间序列预测模型
采用线性平稳时间序列预测种群年龄结构动态,其计算公式详见文献[31]。本研究对枫香种群经过Ⅱ、Ⅲ、Ⅴ、Ⅶ龄级时间后的各龄级个体数进行预测。
1.3 数据分析
用Excel 2016进行数据处理, SPSS 20.0进行数据分析,Origin 9.0进行图形处理。
2 结果与分析
2.1 针阔混交林固定样地内植被特征
研究发现,紫金山南麓固定样地内胸径≥5 cm的树种有36种,总计1 616株,林分密度为1 188 株/hm2,平均胸径为21.3 cm,平均树高为12.5 m。在这36个乔木树种中,针叶树为马尾松(Pinusmassoniana)、水杉(Metasequoiaglyptostroboides)、柳杉(Cryptomeriajaponica)、侧柏(Platycladusorientalis)和杉木(Cunninghamialanceolata),共计267株。针阔混交林固定样地内针叶树与阔叶树的比例为1.0∶5.1。重要值前10位树种依次为枫香、马尾松、樟(Cinnamomumcamphora)、山胡椒(Linderaglauca)、椤木石楠(Photiniabodinieri)、水杉、柳杉、黄连木(Pistaciachinensis)、糙叶树(Aphanantheaspera)和石楠(Photiniaserratifolia),共计1 373株,占总株数的84.93%(表1)。固定样地内胸径≥5 cm枫香有528株,其相对密度(32.67%)、相对显著度(18.73%)和相对频度(34.44%)的总和最高,因而可被认为是针阔混交林主要群落(枫香+马尾松)建群种之一。另外,固定样地内植物多样性较高(Shannon-Wiener指数:乔木层3.473,灌木层3.840,草本层3.688),其中灌木有43种,草本有44种,藤本有8种。

表1 紫金山针阔混交林固定样地重要值前27个树种概况
2.2 种群结构和动态变化指数


图1 紫金山固定样地内枫香种群年龄结构图Fig.1 The age structure of Liquidambar formosana population in the permanent plot under the coniferous and broad-leaved mixed forest in Zijin Mountain, Nanjing City
2.3 种群静态生命表、存活曲线、死亡率曲线和消失率曲线
经分析,固定样地内枫香种群标准化存活个体数(lx)和生命期望值(ex)整体上随龄级增加而降低(表2)。

表2 紫金山固定样地内枫香种群静态生命表
通过比较生存曲线Deevey-Ⅱ(R2=0.663)与Deevey-Ⅲ(R2=0.406)拟合模型,结合静态生命表中种群死亡率qx前期较低后期较高(表2)可知,枫香种群的存活曲线趋于Deevey-I型(图2)。由图2可知,枫香种群死亡率、消失率曲线的变化趋势相似,总体上随龄级增加而增加,局部存在一定波动。

图2 紫金山固定样地内枫香种群存活率、死亡率和消失率曲线Fig.2 The curves of living population, mortality and disappearance rates of Liquidambar formosana population in the permanent plot under the coniferous and broad-leaved mixed forest in Zijin Mountain, Nanjing City
2.4 种群生存变化规律
分析表明,枫香种群生存率随龄级增加而逐渐降低,累积死亡率随龄级单调递增,二者互补(表3),并且在第Ⅵ龄级之后,种群生存率与累积死亡率逐渐趋于平缓。种群死亡密度仅在Ⅱ、Ⅲ龄级间数值较大,在其他龄级间相对较小;危险率则随龄级单调递增,表明固定样地内枫香种群抵抗力稳定性逐渐增强,而恢复力稳定性逐渐下降。
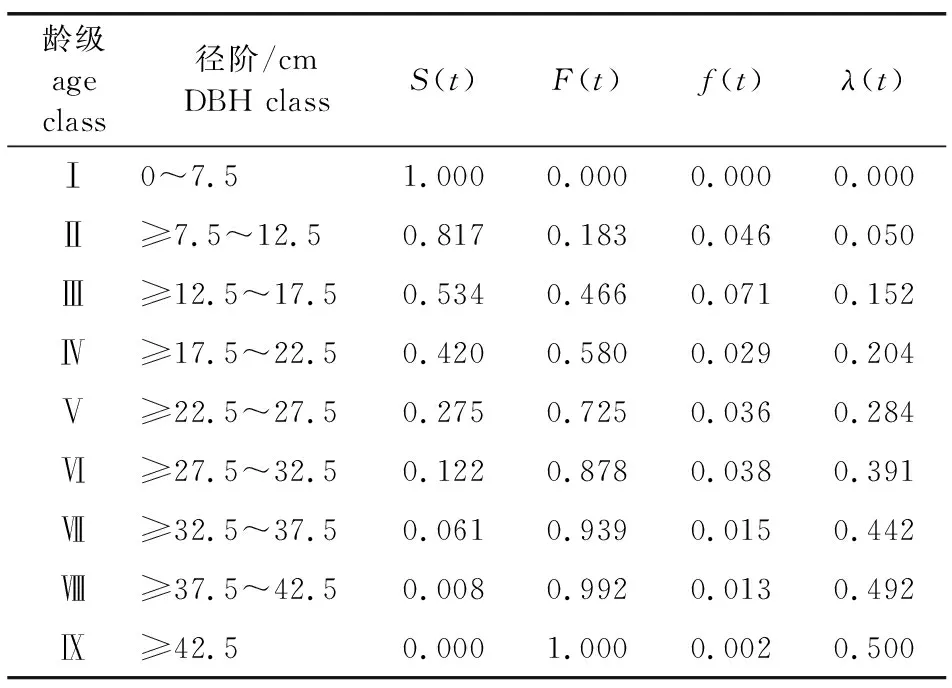
表3 紫金山枫香种群生存函数估算值
2.5 种群个体数量的时间序列预测
基于固定样地内枫香种群各龄级的个体数,利用一阶移动平均法,对经过Ⅱ、Ⅲ、Ⅴ、Ⅶ龄级时间后种群各龄级的个体数量进行预测,结果见表4。在经过Ⅱ龄级时间后,除龄级Ⅱ外,枫香种群的各龄级个体数均有不同程度的增加;经过Ⅲ、Ⅴ龄级时间后,枫香种群内中龄树(Ⅱ—Ⅵ)依旧是种群主体;并且,种群老龄树(Ⅶ—Ⅸ)个体数量在经过Ⅴ、Ⅶ龄级时间后得到了大幅提升。

表4 紫金山固定样地内枫香种群数量动态变化的时间序列预测
3 讨 论
在当前演替阶段,针阔混交林中枫香种群个体数量较多,径阶结构完整且为“金字塔”型,具有较多的苗木数量,具备较好的天然更新能力,这与干扰条件下的枫香种群呈衰退型结构特征[17, 19-20]显著不同。这是因为枫香为强阳性树种,其苗木稍耐阴,喜温湿润气候和深厚的土壤条件,不耐水涝,极易在北亚热带至部分暖温带的森林群落中成为优势树种;并且,枫香种子小且具翅,能够借助风力传播,易在山脚处散布;此外,紫金山南麓针阔混交林为砂质酸性壤土,土壤深厚,海拔较低,坡度平缓,适合枫香种群的天然更新。有研究表明,在受干扰较小的马尾松-枫香针阔混交林中,枫香种群具有较为丰富的实生幼苗库,并且能够通过萌芽更新主动适应环境压力[17]。
本次研究发现,紫金山南麓枫香种群存活曲线为Deevey-I凸线型,与安徽省黄山市查湾自然保护区枫香种群存活曲线一致[22],均表现出种群死亡率和消失率在幼龄期较低,在老龄期较高,这表明自然状态下的枫香种群个体在胸径达到Ⅶ龄级(≥32.5~37.5 cm)后可能逐渐进入生理寿命,对外在环境干扰(如种间竞争和病虫害等)的抵抗力较差,符合枫香可能是乔木树种中偏向r-对策者的推测[21]。时间序列模型的预测结果表明,枫香种群能够在未来较长的演替时间内仍呈增长趋势,这与紫金山北坡红柴枝(Meliosmaoldhamii)种群[32]、南坡栓皮栎(Quercusvariabilis)种群[2]等地带性优势种群相类似,这是因为紫金山森林质量持续改善,郁闭度较高,阳性先锋树种马尾松已无林下更新[18];虽然在紫金山南麓针阔混交林中马尾松的重要值较大,但随着森林群落进展性演替,枫香及其他地带性乡土树种会逐渐占据马尾松种群生态位,实现种群个体数量的正增长。并且,紫金山南麓枫香林空间结构(混交度、角尺度、大小比数和开敞度)较为合理,群落处于较高的演替阶段[18],因而具有较强抵抗力。此外,有研究表明,位于阳坡下坡位的枫香种群分形维数较高,生态间隙维较大,表现出较强的生态空间扩展潜力[19]。
