木霉菌作用机制及其在药用植物上的应用研究进展和展望
2023-05-23马明磊王秋霞马莹莹金桥张舒娜耿业业
马明磊 王秋霞 马莹莹 金桥 张舒娜 耿业业
摘要:本文首先介绍了木霉菌的概况、分类和特性,并从木霉菌的重寄生作用、竞争作用、协同拮抗作用、诱导植物产生抗性、溶菌作用和抗生作用这6个方面描述了木霉菌自身和通过植物体的作用机制。木霉菌是一种无害、无毒、广谱抗性高的真菌,因而得到广泛的开发与应用,取得较好的应用效果。但目前木霉菌的利用还不平衡,多用于蔬菜、果树等园艺植物和小麦、玉米等农作物的病害生物防控方面,比如番茄灰霉病、香蕉枯萎病、小麦根腐病、玉米叶斑病等。但是木霉菌在药用植物上的应用较少。药用植物是经济价值比较高的植物,2020年《中国药典》对药用植物农药残留含量有了更加严格的规定。所以木霉菌作为一种有效的生物防治剂,在药用植物上还有很大的发展空间。最后介绍了木霉菌在药用植物土壤改良、提高抗性、病害防控等方面的应用进展,为木霉菌在药用植物上的深入开发和利用提供了参考。
关键词:木霉菌;分类特点;作用机制;药用植物;应用
中图分类号:S182 文献标志码:A
文章编号:1002-1302(2023)08-0008-09
基金项目:国家自然科学基金(编号:81903755);吉林省科技厅重点研发项目(编号:20220202114NC、20210202096NC);吉林省科技发展计划(编号:20210401106YY);长春市重点研发计划(编号:21ZGY17);延边州科技发展计划(编号:2021NS10);农艺与种业领域案例库建设(编号:ALK[2019]028)。
作者简介:马明磊(1997—),男,山东淄博人,硕士研究生,主要从事资源利用与植物保护研究。E-mail:925344372@qq.com。
通信作者:耿业业,博士,副教授,主要从事野生动植物保护与利用方面研究。E-mail:2305565869@qq.com。
随着农业生产的不断发展,农药虽然能够减少病害,但是会造成严重的环境污染,甚至危及人类生命健康,因此生物绿色防治变得愈发被重视。木霉菌制剂是一种生物性农药,目前使用木霉菌代替化学防治是研究较多的方法之一。近年来,木霉的研究和利用迅速发展。木霉菌及其代谢物在促进植物生长、提高养分利用率、增加产量、增强抗病性等方面有着广阔的应用价值,尤其是在当前国家倡导绿色农业的大背景情况下,大规模地发展木霉已成为建立现代高效生物控制体系的一个重要组成部分。木霉菌在园艺植物和农作物上都得到了广泛应用,但在药用植物方面的应用相对较缺乏,需要进行一系列探究,为今后木霉菌在生物防治中的应用做出参考。
1 木霉菌概况
1.1 木霉菌分类
木霉菌(Trichoderma)属于半知菌亚门丝孢纲丛梗孢目丛梗孢科木霉属。它的分布较为广泛,普遍地存在于各种不同的环境中,如根系、叶片、土壤。1932年,Weinding首次发现木霉菌能够抑制植物病原菌的生长[1],并发现该菌能在植物发生病害时的生物防治中发挥重要作用。因此从那时开始展开了对木霉菌生防研究的工作。常见种类有绿色木霉(Trichoderma.viride)、哈茨木霉(T.harzianum)、瑞氏木霉(T.reesei)、康氏木霉(T.koningii)、拟康氏木霉(T.pseudokoningii)和刺孢木霉(T.asperellum)等[2]。随着生物防治菌剂的发展,研制出的木霉菌制剂已达上百种,涉及到多种植物领域,其中木霉菌在药用植物上的研究与应用也日益增多。
1.2 木霉菌的种类和特性
木霉菌制剂作为一种生物性农药逐渐被应用。其纤维素的降解能力很强,生长速率主要表现在木质素和纤维素丰富的基质上,并且易于繁殖。木霉菌能在植物的根际繁衍,并利用土壤的空间和养分来控制有害的真菌;同时,它还能通过分泌抗生物质来抑制有害真菌的繁殖,从而达到控制病害的目的。但是,木霉菌对田间防治效果、土壤微生物的安全性和有效期尚存在不确定性[3]。近年来,木霉菌作为生物菌剂被广泛应用。结果表明,木霉菌丝具有较高的抗逆性,有利于土壤环境的竞争和群体的形成;能产生抗生物质,对霉菌有一定的效果;由于其特殊的超寄生特性,能在病原菌上引起病原菌死亡,例如能在立枯丝核菌上引起立枯丝核菌体收缩致死。
1.3 木霉菌的生长条件
适宜木霉菌生存的温度为25~35 ℃。木霉菌对生长环境要求很高,一般相对湿度要达到92%以上,93%~95%的湿度才能使其孢子形成,只有生活在相对潮湿的土壤中生命力才较强[3]。另外,最适宜木霉菌生长的pH值为3~7,因此相对酸性的土壤更适合其分生孢子的生长。
2 木霉菌的作用机制
2.1 重寄生作用
在木霉菌的抗性机理中,重寄生物是其重要的拮抗机制。尤佳琪等对7种不同的木霉菌进行了遗传分析,结果表明,重寄生可能是木霉菌的原生生活形式[4]。这是一个相对复杂的过程,其中包括对病原菌的识别、接触、穿透和寄生等一系列步骤。在确定了寄主后,木霉菌和宿主之间形成了一种新的寄生关系,产生菌丝,生长到寄主的菌丝并将其穿透,从而获得所需要营养,使病原菌中的菌丝细胞解体,致病原菌丝被破坏,从而使寄主植物有效地抵御病害。赵玳琳等通过棘孢木霉GYSW-6m1对LC0220的重寄生现象观察结果表明,棘孢木霉GYSW-6m1对草莓炭疽病病菌LC0220具有很好的重寄生效果[5]。
2.2 竞争作用
竞争效应指的是木霉菌与病原体在养分、生存空间等方面的矛盾与争斗。由于木霉菌的适应性很强,它可以与病原菌争夺养分,使病原菌获得的营养物质减少,无法正常生长发育。在生存空间方面,由于木霉菌快速生长,获得了更多的生存空间,因此,降低了病原体的生存空间,使得其不能正常地存活,有效地控制了病害的发生。滕艳萍等在木霉防治黄芪根腐病的研究中发现,木霉菌菌丝缠绕并侵入半裸镰孢,使病原菌的细胞变得稀疏,甚至有部分菌丝被分解或断裂,使其生长受到损害[6]。张丽荣等用生长速度法测得木霉菌的生长速度分别是小麦和豌豆的根腐病病菌以及小麦全蚀病病菌的1.5~2.4、6.3~12.8倍,能很好地抑制病原菌的生长[7]。邵红涛等发现,木霉菌菌株對葡萄糖的利用速度比病原菌高,木霉菌菌株S7和S2对可溶性磷的利用速度比病原菌高,而木霉菌菌株则比病原菌更好,特别是对营养的利用速率方面[8]。刘伟杰等通过利用生物控制技术对玉米病害进行了研究,结果显示,在木霉快速生长的情况下,病原菌数量会减少,对玉米植株的危害会大大减弱。木霉菌和病原菌不仅会争夺碳氮营养,也会争夺微量元素,对病原菌有很好的抑制作用,有利于玉米病害的防治[9]。
2.3 协同拮抗作用
木霉菌的协同机制是一种拮抗机制,其中至少有2种或多种机制同时或按一定顺序起作用。燕嗣皇等通过对木霉生防菌与根际微生物的交互作用的研究,证明其对大部分根际真菌均有抑制或超寄生的效果,或二者同时存在,其主要原因是导致菌种数量的下降和区系结构的改变[10]。牛芳胜研究发现,与杀菌剂联合应用的哈茨木霉菌对番茄灰霉菌的抑制作用明显,其抑菌效果比二者单独应用的效果好[11]。张婧迪等的研究结果表明,深绿木霉菌CCTCC-SWB0 199发酵代谢物加上芸薹素内酯按照不同比例混合对番茄灰霉病的防治效果好于两者单独使用[12]。谷祖敏等研究发现,木霉菌对植物病原菌的生防作用通常是多种机制作用的结果,不同菌株的生防机制则有所不同[13]。
2.4 诱导植物抗性
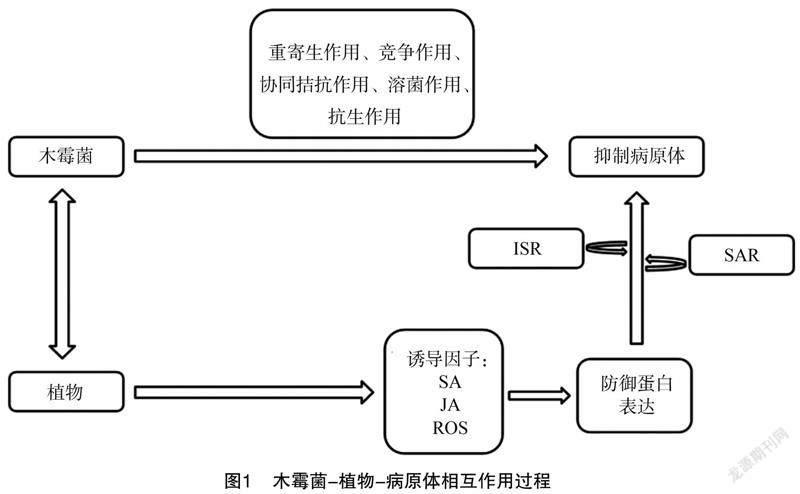
木霉菌的生物防卫机制不仅能反映微生物间的相互影响,而且常常忽略了寄主植物的参与。木霉菌植物的根部定植,释放出一些化合物,导致植物产生防御反应,寄主植物受到刺激后会加强它们对抗病原微生物的抵抗力和有效性,可有效防止侵害。López-Quintero等提出木霉菌属与植物根系形成内源性联系,可能对寄主提供一系列有益帮助,对其机制的研究有待加强[14];同年,Nawrocka等在植物与木霉菌相互作用的过程(图1)中提出,木霉菌丝释放的许多诱导因子可能诱导植物体内传递各种类型的信号,例如水杨酸(SA)、茉莉酸(JA)或活性氧(ROS),触发防御蛋白的表达,使植物有机体产生的酶能直接抑制病原微生物,从而加强植物有机体的生化功能和结构屏障,根据木霉菌菌株、植物种类以及生物和非生物条件,真菌激活的防御反应能够在2种系统性抵抗之间转换:诱导系统性抵抗(ISR)和系統获得性抵抗(SAR)[15]。木霉菌可以通过以下3种方式诱导植物产生抗性:(1)增强由MAMPs分子产生的免疫应答(MTI);(2)将效应因子触发感病的敏感度降低(ETS);(3)加强效应因子诱导的免疫应答(ETI)[16]。一般而言,木霉菌诱导植物的MTI会超过病原体相关分子模式激发的免疫反应(PTI)效应,主要是由于木霉菌可产生大量的MAMPs分子(植物效应分子),例如几丁质酶Chit42、SnodProt蛋白(SnodProt1、Sm1和EPI)、脂肽、棒曲霉素类蛋白、丝氨酸蛋白酶、22 ku木聚糖酶、无毒基因蛋白、几丁质脱乙酰基酶等[17]。此外,植物丝裂原活化蛋白激酶(MAPK)基因(TIPK)可被木霉菌特异性诱导[18]。Djonovic'[DD)]等鉴定出了绿色木霉分泌的效应分子,并证明该效应分子与植物抗性的诱导具有一定的相关性[19]。目前,在木霉菌中已发现10余种能够诱导植物产生抗性的作用分子,包括纤维素酶、蔗糖酶、MRSP1、Sm1、QID74疏水蛋白、几丁质降解酶、木聚糖酶、内切多聚半乳糖醛酸酶、抗菌肽等[20]。张婷的研究结果表明,经木霉菌包衣处理的玉米种子过氧化物酶(POD)和苯丙氨酸解氨酶(PAL)活力明显增强,而且对玉米弯孢菌叶斑的抗性明显增强[21]。此外,王淑霞等利用哈茨木霉对黄瓜根系中的多酚氧化酶、过氧化物酶等进行了检测,结果表明,哈茨木霉对黄瓜植株的防御酶活力有较大影响[22]。辛鑫等采用生防菌绿色木霉、香蕉枯萎病病原菌接种香蕉幼苗,对其超氧化物歧化酶(SOD)、POD、多酚氧化酶(PPO)、PAL、过氧化氢酶(CAT)等的活性进行了比较,结果表明,在接种木霉菌后,香蕉幼苗中 SOD、 POD、 PPO、 PAL、 CAT的活性明显增加,对香蕉的抗病作用有明显的促进作用[23]。
2.5 溶菌作用
木霉菌的溶菌作用是指木霉菌会产生一些抑制病原菌生长的蛋白酶类物质,如蛋白酶、几丁质酶、葡聚糖酶等,这些物质会消化真菌的细胞壁,使细胞壁内的多糖、几丁质、β-葡聚糖被破坏[24]。倪方方等认为,哈茨木霉的次生代谢物能溶解病原菌体的胞壁,造成细胞壁穿孔和变形[25];黄静霞等通过混合木霉菌和黄孢原毛平革菌后,降解病原菌的纤维素酶[26]。黄位年等的研究表明,木霉菌能产生大量的果糖酶、几丁质酶,并能溶解病原菌的细胞壁,从而降低病原体的入侵,对石斛的病害防治有重要影响[27]。
2.6 抗生作用
在代谢时期中,木霉菌会制造出一系列具有抗药性的化学成分,其中包含一些抗生素和酶。木霉菌也能制造出含有挥发性和非挥发性2种不同类型的抗生素。其中包括木霉菌素(triohodexrmin)、抗菌肽(peptide antibiotic)、绿木霉菌素(viridin)、胶霉毒素(gliotoxin)等[28]。木霉菌能释放出挥发性的、能抵抗病原菌的乙醛[29]。哈茨木霉抑制立枯丝核杆菌的机制是通过产生一种带有椰子气味的挥发性抗生素,后来被确定为六戊烷基吡喃和戊烯基吡喃[30]。朱天辉等对哈茨木霉和立枯丝核菌进行了比较,结果表明,FO60菌株的代谢物能显著地抑制立枯丝核菌的菌落生长,同时,非挥发性代谢物对菌丝的热稳定性也有一定的影响[31]。李琳研究发现,棘孢木霉菌的菌丝会在病原菌菌丝上螺旋缠绕,并通过酶的作用破坏病原菌细胞壁后逐渐侵入原生质体内部,从而达到防治效果[32]。徐嘉俐等的研究表明,木霉菌所产的易挥发性和难挥发性代谢物对大丽轮枝菌(Verticillium dahliae)的生长有明显的抑制作用,抑制率达76.3%[33]。
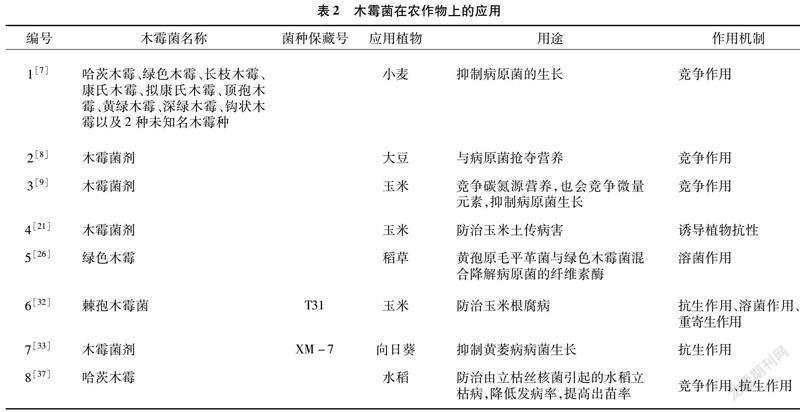
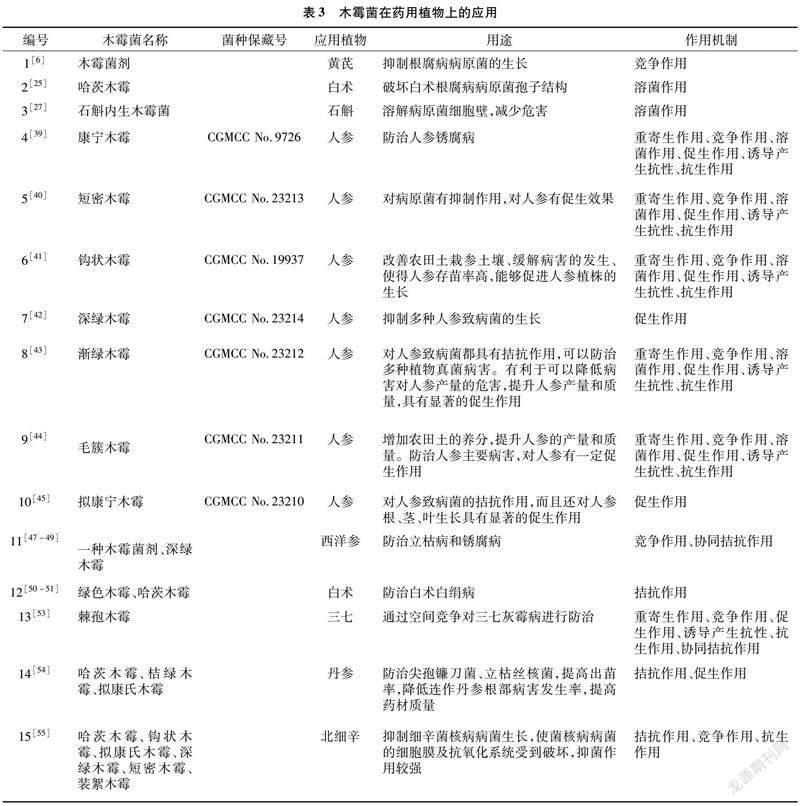
以上的作用机制在我国药用植物方面的研究结果相对较少(表1、表2、表3),研究者仍需要加大研究力度,拓宽研究方法。
3 开发与应用
3.1 木霉菌在生物防治中的应用
对于木霉菌的开发,一般是从自然界中存在的木霉菌中筛选出优良而高效的菌株,然后进一步利用生物技术改善生物体,使其具有更多种功能和更好的特性来预防病害。木霉菌菌丝生长速度很快,具有一定抗逆性,有利于土壤环境的竞争,也利于群体的形成;木霉菌也能产生有抗性的生物质来控制腐霉菌;木霉菌特有的溶菌现象可使其对病原菌
的菌丝起到一定的作用,使植物的病原体死亡,例如,木霉能在镰刀菌上引起镰刀菌萎缩而死亡[34-35]。它的优点在于无污染、无残留毒性,对土壤中的有益微生物无损害,对土壤和生态系统的微生态循环不会产生影响。
木霉属的一些真菌是存在于多种生境中的微生物群落中的重要组成部分。它具有较强的代谢能力和积极的竞争性,可以诱导多种拮抗机制的结合,产生具有抗真菌活性的次级代谢产物;真菌寄生,由于对养分或空间的竞争,从植物病原体中产生细胞壁降解酶;以及通过诱导子分子的产生和分泌诱导植物抗性。木霉菌生物防治的一般机制可分为直接作用和间接作用。其直接的影响有竞争养分或空间、产生挥发性和非挥发性的抗生素和分解酶、致病酶的失活和寄生。其间接作用主要表现在对寄主植物的生物化学和生理上的改变,例如对胁迫耐受性、无机养分的溶解性和隔离作用,以及诱发植物致病的耐药性。一些木霉菌能有效地定植在植物根系表面,引起植物代谢的巨大变化,它有利于植物生长,增加养分利用率,并增加抗病性。木霉菌产生的激发子分子激活植物防御系统的相关基因表达,促进植物根系生长和养分利用,抗性诱导是一种间接的生物控制机制,植物通过激活潜在的抗性机制来应对病原体的攻击。当植物暴露在生物或非生物诱导剂下,这一过程就会发生,以一种相对普遍的方式激活其防御机制,这种激活可以持续不同的时间,不仅在诱导部位,也能在其他远程位置,植物可能会产生植保素、细胞中的额外木质素和酚类化合物[36]。
近年来,木霉菌的应用日益广泛,也取得了较好的效果,例如国外利用木霉菌治理由立枯丝核菌引起的水稻纹枯病,使其发病率得到大幅度下降,出苗率显著提高[37];国内也利用哈茨木霉菌对茉莉白绢病进行了有效的防治[38]。李梅等利用康宁木霉菌和含有康宁木霉菌的复合微生物菌剂防治人参锈腐病,取得了显著成效[39]。王秋霞等利用短密木霉菌能够改良农田土壤,使农田土壤生物丰度更高,有效地控制人參、西洋参叶片干枯的问题,具有明显的促生作用[40]。王秋霞等利用钩状木霉菌MM3、深绿木霉菌改善农田土栽人参土壤、缓解病害的发生、提高人参的存苗率、促进生长,同时渐绿木霉菌、毛簇木霉菌、拟康宁木霉菌等对多种人参致病菌都具有拮抗作用,有利于提高人参的质量和产量,起到促生作用[41-45]。木霉菌除了在园艺植物和农作物等方面有着普遍的应用,在药用植物上也有着广阔的应用前景,同时,可以利用木霉菌在其他方面已有的研究结论,将其合理地运用在药用植物上。由于药用植物与其他栽培作物具有许多相同点,所以,可选择借鉴与已试验成功的栽培植物具有相同点的药用植物进行针对性研究,如棉花立枯病的防治方法可以在多种药用植物苗期病害上进行生物防治试验,甜椒、茉莉白绢病的研究方法可以在对黄连等药用植物白绢病进行生防研究时加以借鉴[46]。关于木霉菌在药用植物上的生物防治的报道不多,利用哈茨木霉菌防治白术根腐病,取得了很好的效果。在木霉菌防治西洋参立枯病研究方面也获得了成功[47-49]。除了以上药用植物病害,白术[50-51]、黄连的白绢病,白芨、地环、人参的根腐病[52],三七的灰霉病[53],丹参[54]、三七的根腐病以及丹参、人参、西洋参、北细辛[55]的菌核病等研究也很少,但前景十分广阔[46]。木霉菌在园艺植物、农作物、药用植物上的应用见表1、表2、表3。
目前在国外,有关木霉菌的研究和应用相对较早,发展很快,已经有了许多注册的木霉菌,如T22、T39等产品。国外对木霉菌的生防机制也有了较深的研究,不但掌握了木霉菌的生产工艺、先进设备,而且还对其生物防治的代谢物种类、化学结构、作用机制等进行了较全面的探讨。近年来,国内外学者运用蛋白质组学、基因报告技术等新手段,研究了植物、病原菌、木霉3个方面的相互影响,并对其进行基因表达的差异性分析,以更好地理解其生防机制,为今后发展木霉菌新的生物控制技术提供理论依据。但在我国,有关方面的研究并不多,在药用植物上也未进行过深入研究。
3.2 土壤改良与环境修复
由于土壤常年在使用状态下,得不到休整,随着化肥和农药使用的增多,加上连作,土壤质量受到了极大破坏。通过灌根、拌种等多种途径,利用木霉菌作为生防因子,可以有效地促进其根系的生长和增殖。土壤中细菌、放线菌和固氮菌的种群数量也有所增加。木霉菌能通过调节土壤理化性质,改善土壤的生态状况,减小土的比重,减小容重,改善土壤的结构,提高pH值,降低土壤酸化的危害,促进土壤中有益的微生物群落的形成和维持;增强土壤酶活性,促进植物对氮、磷、钾和微量元素的吸收,提高化肥的使用效率;提高土壤的渗透性,能提高含氧量,改善土壤的新陈代谢和循环,从而促进植物和微生物的生长[56-57]。木霉菌对环境有较好的适应能力,可以通过多种简单或复合的碳源和氮源进行生长,具有制备微生物肥料和土壤重金属修复相关功能,对Cu和Zn等重金属均有较强的抗性,能生成吸收或累积重金属的纤维素酶及其他水解酶,是极具开发潜力的用于强化植物修复重金属污染土壤的真菌[58]。利用麦麸发酵物,可为木霉菌在植物根际的定植提供充足的养分,从而为木霉菌的生长和繁殖创造有利的条件,并能产生促使其在培养土中生长繁殖、根系生成的生防因子,能将土壤中主要的酚酸盐物质分解,降低连作障碍,降低土壤病害的发生概率[59]。
随着我国农业的发展,对农药和化肥的使用也日益频繁,其中最为显著的是各种化学农药的过度使用。由于其毒性大,在环境中的存在备受关注。它们在环境中相互作用、共存,极大地影响和威胁着生态环境和人类健康。此外,这些污染物会影响土壤质量和肥力,修复此类环境常采用物理化学技术,但其效率较低,且需要较高的运营成本。生物修复是一种高效、广泛、经济、环保的清洁方式。利用微生物作为一种有效的生物技术策略来净化环境,对环境的风险最低[60]。因此在生物防治方面愈来愈得到人们重视。已知有许多微生物属参与生物修复,其中就有木霉菌。木霉菌是一种重要的植物病害生物防治剂,除了生物防治特性外,它们还有一个非常独特的土壤生物修复现象。在这种情况下,土壤覆盖物的生物修复和土壤微生物群的恢复尤为重要。在土壤中引入微生物是当前改善农业和林业土壤产量最有希望的方法之一。与使用单一菌株相比,使用不同菌株的木霉菌进行共培养可以更快更高效地分解固体废物。然而,这些物种也有能力依靠其他真菌生存,并具有在植物根和根际定居的能力,因此,利用木霉菌来进行环境修复是一项很有前途的技术。事实上,微生物提供了一个有用的平台,可以用于各种环境污染物的强化生物修复模型。微生物惊人的代谢能力应该得到利用,以便在降解途径的进化、生物修复和生物转化过程的新策略的开发方面取得新的突破[60]。
4 结语与展望
药用植物是经济价值比较高的植物,2020年《中国药典》对药用植物农药残留含量检测有了更加严格的规定,从原来的7种提升至33种。因此,要更加科学地施用化肥农药,不断改善种植环境,使药用植物的品质提高。开发微生物防治菌剂反映了科技进步,新技术新方法的运用必将促进生物防治,并将其作为一种重要的防治手段。木霉菌作为一种安全、有效的生防菌剂,在药用植物病害防控、农药替代、生物菌肥开发方面具有重要的应用价值[61]。生物技术的飞速发展给生物防治带来了新的机遇,也使得木霉菌剂在医药领域的应用有了更广泛的发展。结合现代生物技术,借鉴木霉菌剂在园艺作物和农作物上的应用经验,从多个方面开展药用植物木霉菌剂的开发和應用研究。
4.1 复合菌剂的开发
根据木霉菌资源的丰富性,对药用植物特有的生防菌剂进行筛选,将其混合开发出复合菌剂,从而更加有针对性地防治相应的病害或改良土壤。单一菌剂是只由一种微生物制成的,功能单一,对环境的适应能力差;复合菌剂是用2种或更多种功能的微生物组合而成,它的促生作用更强,更加稳定,是微生物菌剂未来的发展趋势。对单个木霉真菌与植物病原的拮抗机制已有一定认识,但研究各种菌对植物病原菌的拮抗机制仍是一个很大的挑战[62]。目前对构建微生物复合菌剂的研究不够深入,也需要进一步探索[63]。
4.2 次生代谢产物的应用
利用生物技术和发酵技术,提取和分析木霉菌次生代谢产物,筛选关键成分,研制生物源农药,利用其次级代谢物进行微生物制剂的开发,将成为今后发展的主要途径。木霉属真菌的次生代谢物主要有聚酮类、肽类、萜类和其他类物质[64]。可通过扩大植物幼苗根系的吸收范围和能力,增强幼苗对矿质元素的吸收,减少水分散失,使幼苗更好地保持和利用土壤中的水分。例如绿色木霉可通过自身或周围的微生物和植物产生的各种酶和次生代谢物,对有机物进行降解,使土壤中的有机质被激活,这样可以改进土壤的构造,降低干旱胁迫对玉米幼苗的伤害[65]。
4.3 与基因工程技术相结合
采用基因工程技术对微生物菌剂进行改造,开发出适应性更强,对病原菌抗性更强的目标菌株,实现微生物菌剂工业化和产业化。由于土壤环境受污染情况较严重,污染物的组成是多种多样的。但是,目前微生物制剂的品种单一,功能低效,难以适应目前的需求,缺乏复合、效率较高、目标性相对明确的微生物菌剂。因此,加强微生物的筛选和驯化,特别是针对不同微生物协同共生研究,借助基因工程技术手段,对生防菌株进行遗传改良,提高生防菌株的生防效果、拮抗性能,扩大其防治对象,培育和生产出安全、高效的复合微生物菌剂,从而推动微生物菌剂在各产业上的应用[66-67]。
总之,由于人类对生态环境和卫生问题的关注,生物防治技术的研究和应用越来越被人们所关注,而木霉菌是一种重要的植物病害生物控制菌,它的研究不仅丰富了这方面的知识,而且也使人类对其疾病的生物控制有了更多的认识。各种木霉产品的商业化,必然会激发科研单位和企业之间的合作,推动其研究和应用的进一步深化和扩大。
参考文献:
[1]Weindling R. Studies on a lethal principle effective in the parasitic action of Trichodermu lignorum on rhizoctoniasolani and other soil fung[J]. Phytopathology,1932,22:837-845.
[2]商 娜,任爱芝,赵培宝. 木霉菌在园林园艺植物上的应用及研究进展[J]. 北方园艺,2021(3):149-154.
[3]张凌萱,孙 影. 生防真菌之木霉菌的应用[J]. 广东蚕业,2020,54(6):113-114.
[4]尤佳琪,吴明德,李国庆. 木霉在植物病害生物防治中的应用及作用机制[J]. 中国生物防治学报,2019,35(6):966-976.
[5]赵玳琳,何海永,吴石平,等. 棘孢木霉[WTBX][STBX]GYSW-6m1[WTBZ][STBZ]对草莓炭疽病的生防机制及其防病促生作用研究[J]. 中国生物防治学报,2020,36(4):587-595.
[6]滕艳萍,梁宗锁,陈 蓉. 木霉防治黄芪根腐病初步研究[J]. 西北农业学报,2006,15(2):69-71.
[7]张丽荣,康萍芝,沈瑞清. 木霉菌对土传病害病原真菌的拮抗作用[J]. 内蒙古农业科技,2007,35(5):48-50.
[8]邵红涛,许艳丽. 具有生防能力的木霉菌(Trichoderma spp.)与两株大豆根腐病病原菌(Fusarium oxysporum、Rhizoctonia solani)对碳、磷、铁的竞争研究[J]. 黑龙江大学自然科学学报,2007,24(1):126-129.
[9]刘伟杰,李 民,刘艳红,等. 玉米病害生物防治中木霉菌的应用研究[J]. 世界热带农业信息,2021(10):19-20.
[10]燕嗣皇,吴石平,陆德清,等.木霉生防菌对根际微生物的影响与互作[J]. 西南农业学报,2005,18(1):40-46.
[11]牛芳胜. 哈茨木霉菌与5种杀菌剂联合对番茄灰霉病菌的协同作用及增效机制研究[D]. 保定:河北农业大学,2013:13-19.
[12]张婧迪,陈 捷,刘志诚,等. 深绿木霉菌发酵代谢物与芸薹素内酯复配效果评价[J]. 上海交通大学学报(农业科学版),2017,35(5):1-7.
[13]谷祖敏,毕 卉,张 兵,等. 不同木霉菌株对黄瓜枯萎病菌的拮抗作用[J]. 西北农业学报,2018,27(3):426-431.
[14]López-Quintero C A,Atanasova L,Esperanza Franco-Molano A,et al. DNA barcoding survey of Trichoderma diversity in soil and litter of the Colombian lowland Amazonian rainforest reveals Trichoderma strigosellum sp.[J]. Antonie Van Leeuwenhoek,2013,104(5):657-674.
[15]Nawrocka J,Malolepsza U. Diversity in plant systemic resistance induced by Trichoderma[J]. Biological Control,2013,67(2):149-156.
[16]陈 捷,朱洁伟,张 婷,等. 木霉菌生物防治作用机理与应用研究进展[J]. 中国生物防治学报,2011,27(2):145-151.
[17]Shoresh M,Harman G E,Mastouri F. Induced systemic resistance and plant responses to fungal biocontrol agents[J]. Annual Review of Phytopathology,2010,48:21-43.
[18]梁巧兰,魏列新,徐秉良,等. 深绿木霉T2蛋白质TraT2A诱导处理对接种灰霉菌百合光响应和荧光特性的影响[J]. 园艺学报,2020,47(4):769-778.
[19]Djonovic'[DD)] S,Pozo M J,Dangott L J,et al. Sm1,a proteinaceous elicitor secreted by the biocontrol fungus Trichoderma virens induces plant defense responses and systemic resistance[J]. Molecular Plant-Microbe Interactions:MPMI,2006,19(8):838-853.
[20]陈 捷. 木霉菌诱导植物抗病性研究新进展[J]. 中国生物防治学报,2015,31(5):733-741.
[21]张 婷. 木霉菌生物种衣剂的创制与应用[D]. 上海:上海交通大学,2012:69-75.
[22]王淑霞,张丽萍,黄亚丽,等. 哈茨木霉Tr-92诱导黄瓜对灰霉病系统抗性的研究[J]. 中国生物防治学报,2013,29(2):242-247.
[23]辛 鑫,刘 磊,潘江禹,等. 绿色木霉H6对香蕉枯萎病的诱导抗性作用[J]. 广东农业科学,2013,40(7):83-85.
[24]赛牙热木·哈力甫,鄧 勋,宋小双,等. 木霉菌生物防治及促进植物生长机制研究进展[J]. 吉林农业大学学报,2020,42(3):237-247.
[25]倪方方,徐红梅,宋腾蛟,等. 生防菌对白术根腐病菌的拮抗作用及盆栽防治效果[J]. 浙江中医药大学学报,2017,41(3):179-185,204.
[26]黄静霞. 黄孢原毛平革菌与绿色木霉菌混合降解稻草过程的研究[D]. 长沙:湖南大学,2015:5-6.
[27]黄位年,曾华兰,华丽霞,等. 内生木霉菌与石斛相互作用关系的研究进展[J]. 江苏农业科学,2020,48(13):57-61.
[28]阮盈盈,刘 峰. 木霉菌生物防治作用机制与应用研究进展[J]. 浙江农业科学,2020,61(11):2290-2294.
[29]刘亚苓. 两种拮抗菌对细辛叶枯病的防治效果及机制研究[D]. 北京:中国农业科学院,2020:2-3.
[30]Mukherjee P K,Horwitz B A,Kenerley C M. Secondary metabolism in Trichoderma:a genomic perspective[J]. Microbiology,2012,158(Pt 1):35-45.
[31]朱天辉,邱德勋. Ttrichoderma harzianum对Rhizoctonia solani的抗生现象[J]. 四川农业大学学报,1994,12(1):11-15.
[32]李 琳. 棘孢木霉菌的分离及其生防作用的评价与应用[D]. 长春:吉林大学,2013:13-19.
[33]徐嘉俐,王智慧,康 静,等. 向日葵黄萎病菌拮抗木霉菌的筛选[J]. 内蒙古农业大学学报(自然科学版),2016,37(3):21-25.
[34]王强强,窦 恺,陈 捷,等. 拮抗性木霉菌株抗逆性筛选评价标准与方法[J]. 中国生物防治学报,2019,35(1):99-111.
[35]柳婷婷. 两种木霉对国槐根茎腐烂病菌作用效果比较研究[D]. 聊城:聊城大学,2019:42-46.
[36]Silva R N,Monteiro V N,Steindorf A S,et al. Trichoderma/pathogen/plant interaction in pre-harvest food security[J]. Fungal Biology,2019,123(8):565-583.
[37]Devi K S,Devi P H S,Sinha B,et al. Invitro evaluation of native rice specific isolates of Trichoderma against rice sheath blight caused by Rhizoctonia solani[J]. International Journal of Current Microbiology and Applied Sciences,2020,9(7):1658-1664.
[38]杨万业. 茉莉花白绢病预防措施和处理方法[J]. 南方农业,2021,15(32):51-52,55.
[39]李 梅,陈书华,蒋细良,等. 一种木霉菌及含有该菌的菌剂和它们在防治人参锈腐病中的应用:CN104498369A[P]. 2015-04-08.
[40]王秋霞,马莹莹,金 桥,等. 一种短密木霉、人参病原菌抑菌剂及其应用:CN113913304A[P]. 2022-01-11.
[41]王秋霞,马莹莹,关一鸣,等. 一种钩状木霉菌、含有该菌的微生物菌剂及其应用:CN112063538A[P]. 2020-12-11.
[42]王秋霞,马莹莹,金 桥,等. 一种深绿木霉及抑制人参致病菌中的应用:CN113913302A[P]. 2022-01-11.
[43]王秋霞,马莹莹,金 桥,等. 一种渐绿木霉、人参属植物土壤改良剂及其应用:CN113881578A[P]. 2022-01-04.
[44]王秋霞,马莹莹,金 桥,等. 一种毛簇木霉、菌剂及其应用:CN113913301A[P]. 2022-01-11.
[45]王秋霞,马莹莹,金 桥,等. 一种拟康宁木霉、浸根液及应用:CN113913303B[P]. 2022-11-04.
[46]丁万隆,程惠珍,张国珍. 木霉在药用植物病害防治上的应用[J]. 中草药,1997,28(8):505-507.
[47]张爱华,雷锋杰,强 薇,等. 2株拮抗菌剂复配对西洋参立枯病和锈腐病的防治及促生长作用[J]. 湖南农业大学学报(自然科学版),2014,40(6):611-614.
[48]李纪顺,陈 凯,王贻莲,等. 防治西洋参立枯病木霉菌株的筛选鉴定及其小区防治效果[J]. 山东科学,2019,32(5):62-70.
[49]王 刚,钟均超,刘俊玲,等.生物农药——木霉对西洋参立枯病的防治研究[J]. 人参研究,2003,15(1):41-42.
[50]张晓梦,田永强,潘晓梅,等. 2株木霉抑菌效果及其促植物生长机制[J]. 南方农业学报,2020,51(11):2713-2721.
[51]韩金龙,朱金英,裴艳婷,等. 白术连作障碍机制与缓解措施分析[J]. 农业科技通讯,2020(8):212-215.
[52]张锋涛,崔 佳,张荣意,等. 生防木霉菌发酵条件优化、不同剂型研制及促生防病效果评价[C]// 彭友良,王 琦.中国植物病理学会2018年学术年会论文集. 北京:中国植物病理学会. 2018:481.
[53]蒋 妮,白丹宇,宋利沙,等. 棘孢木霉F2菌株对三七灰霉病的生物防治作用[J]. 江苏农业科学,2018,46(20):94-97.
[54]王 雪,陳美兰,杨 光,等. 丛枝菌根真菌与哈茨木霉菌合用对连作丹参生长及质量的影响[J]. 中国中药杂志,2014,39(9):1574-1578.
[55]王子晴,卢宝慧,田义新,等. 拮抗北细辛菌核病木霉菌的分离、鉴定及生防效果[J]. 微生物学通报,2021,48(12):4624-4635.
[56]陈建爱,杜方岭. 黄绿木霉T1010对樱桃番茄横向土壤环境性状改良效果研究[J]. 农学学报,2011,1(8):36-41.
[57]周红姿,祁 凯,赵晓燕,等. 杏鲍菇菌糠木霉发酵物对大棚土壤的改良效果[J]. 福建农业学报,2017,32(12):1359-1364.
[58]田 晔,滕 应. 木霉属真菌在重金属污染土壤生物修复中的应用潜力分析[J]. 科学技术与工程,2013,13(36):10877-10882.
[59]张 璐,杨瑞秀,王 莹,等. 甜瓜连作土壤中酚酸类物质测定及降解研究[J]. 北方园艺,2017(9):18-23.
[60]Kour D,Kaur T,Devi R,et al. Beneficial microbiomes for bioremediation of diverse contaminated environments for environmental sustainability:present status and future challenges[J]. Environmental Science and Pollution Research,2021,28(20):24917-24939.
[61]谈 韫,樊 航,张紫瑶,等. 绿色木霉菌株发酵液及分生孢子悬浮液对南方根结线虫二龄幼虫活性的影响[J]. 江苏农业科学,2022,50(12):114-120.
[62]王家有,赵 曦,冷春旭,等. 籽用南瓜辣椒疫霉菌防控技术[J]. 安徽农学通报,2022,28(5):126-128.
[63]赵思崎,王敬敬,杨宗政,等. 微生物复合菌剂的制备[J]. 微生物学通报,2020,47(5):1492-1502.
[64]杨曦亮,任梦瑶,刘 倩,等. 近十年海洋来源木霉属真菌次生代谢产物研究进展[J]. 广西科学,2021,28(5):440-450.
[65]邓 薇,张祖衔,曹宇航,等. 绿色木霉缓解干旱胁迫对玉米幼苗根系生长的影响[J]. 山东农业科学,2022,54(2):40-45.
[66]黄 勇,董运常,罗伟聪,等. 景观水体生态修复治理技术的研究与分析[J]. 环境工程,2016,34(7):52-55,164.
[67]王 雪,田 佳,徐琳琳,等. 表达hrpZPsg12基因的重组枯草芽孢杆菌工程菌株构建及其生防活性评价[J]. 西北农林科技大学学报(自然科学版),2015,43(11):139-144.
