葡萄采后摘叶处理对诱导休眠的影响
2023-04-01李瑭白瑞雯邹利人闫可申海林齐晓光温景辉
李瑭,白瑞雯,邹利人,闫可,申海林,齐晓光,温景辉
(1. 吉林农业大学园艺学院,吉林长春 130118;2. 吉林省农业科学院果树研究所,吉林公主岭 136100)
休眠是指植物体或其器官在发育的某个时期生长和代谢暂时停顿的现象[1-3],是落叶果树生长发育中必不可少的组成部分。果树进入休眠后,树体外观停止生长,但内部营养物质含量和酶活性仍在发生变化[4-5],这些变化在增强植株抗逆性的同时,也为果树下一个生长发育阶段积累养分和能量[6]。影响休眠的因素包括树种与品种、外界因素(光照、温度、水分等)、内部因素(糖、酶、激素等)、休眠相关基因(PHY、CBF、CYC、DAM、SVP等)、表观遗传调控机制等[7]。葡萄具有产量高、适应性广、结果年限长、经济效益高等特点,是助力乡村振兴战略实施的重要树种。诱导葡萄提前休眠,可抵御北方早期断崖式降温产生的不良影响,有助于葡萄安全越冬,在生产上具有重要意义。
‘着色香’葡萄为东北地区主栽品种,色泽鲜艳、穗型美观、味甜、口感好、抗逆性强[8]。‘日150’葡萄为欧美杂交种,二倍体,果穗中等大,果皮紫红色、味甜、口感好、中熟、抗逆性强[9],为东北地区极具发展潜力的鲜食葡萄品种。前人研究表明,果实转色期及成熟期摘叶能提高葡萄果实成熟度及葡萄酒品质[10];还能提高山葡萄果实中糖、单宁、花色苷和总酚含量,同时降低果实酸含量[11],而关于人工摘叶诱导葡萄休眠的研究报道较少。本研究通过人工摘叶诱导葡萄提前进入休眠,研究不同摘叶量对树体内养分含量、抗氧化酶活性、以及对休眠相关基因表达量的影响,旨在为寒地葡萄安全越冬和优质生产提供新的技术支撑。
1 材料与方法
1.1 试验材料
试验于2020年在吉林省农业科学院果树研究所核心技术试验示范园(吉林省公主岭市)进行,果园土质为壤土,土层深厚。以生长健壮、长势一致的露地栽培5年生‘着色香’和‘日150’为试材。株距1.0 m,行距为4.0 m,厂形架式,常规水肥管理。冬季修剪后,低温来临前(10月中旬)将枝蔓下架,行间适量取土覆盖枝蔓后,再覆盖宽1.5 m、厚20 cm的玻璃丝棉防寒被,保证植株安全越冬。
1.2 试验处理
果实采收后(9月17日),从枝条基部向上分别摘去叶片总量的20%、40%、60%、80%、100%,以不摘叶为对照(CK)。每处理5株。于11月8日(处理后52 d),各处理随机采集一年生健壮枝20条,截取枝条中段20 cm的韧皮部,液氮研磨后放入﹣80 ℃冰箱保存备用。
1.3 养分含量及酶活性测定
可溶性糖与淀粉含量使用蒽酮法测定,氨基酸含量用茚三酮溶液显色法测定,游离脯氨酸含量用茚三酮比色法测定,可溶性蛋白含量用考马斯亮蓝法测定[12]。超氧化物歧化酶(SOD)活性用氯化硝基四氮唑蓝(NBT)光化还原法测定,过氧化物酶(POD)活性用愈创木酚法测定。使用上海仪电分析仪器有限公司生产N2型可见分光光度计进行比色测定。
1.4 相关基因半定量表达分析
(1)在NCBI数据库中查找5个氮代谢关键酶基因VvGDH(谷氨酸脱氢酶基因,JF796045)、VvNiR(亚硝酸还原酶基因,JF796046)、VvNR(硝酸还原酶基因,JF796047)、VvGS(谷氨酰胺合成酶基因,JF796048)和VvAS(天冬酰胺合成酶基因,JF796049);两个赤霉素合成相关基因VvKO(贝壳杉烯氧化酶基因,JQ086553)、VvGA3ox-4(GA3β-羟化酶基因,JQ086556)以及一个脱落酸合成相关基因VvNCED1(9-顺式环氧类胡萝卜素双加氧酶基因,EF625685)的DNA序列,采用Primer 5.0设计引物。
(2)采集枝条样品,用天根多糖多酚植物RNA试剂盒(DP441)提取总RNA,并用Power ScriptⅡTM反转录试剂盒反转录成总cDNA。以cDNA为模板,分别设计5个氮代谢关键酶基因、两个GA合成基因和一个ABA合成基因引物(表1),以葡萄UBI基因为看家基因,进行PCR扩增,用2%琼脂糖凝胶电泳检测扩增产物,观察条带荧光亮度,比较不同处理下的基因表达量。

表1 引物序列及目的片段大小Table 1 Primer sequence and target fragment size
1.5 数据分析
采用MS Office 2013 和SPSS 20.0软件对试验数据进行统计分析。
2 结果与分析
2.1 摘叶对枝条可溶性糖和淀粉含量的影响
由表2可知,摘叶处理后枝条中可溶性糖和淀粉含量均升高。其中,摘叶40%处理下两个品种的枝条中可溶性糖含量均显著高于CK,‘着色香’增加23.40%,‘日150’增加20.34%。说明摘叶处理能促进枝条内可溶性糖的积累,摘叶40%处理效果最为显著。‘着色香’摘叶40%处理后淀粉含量增加最多,较CK增加61.30%;‘日150’摘叶20%和摘叶40%处理下淀粉含量均显著高于CK,摘叶20%处理增加11.08 mg·g-1,摘叶40%处理增加17.17 mg·g-1。
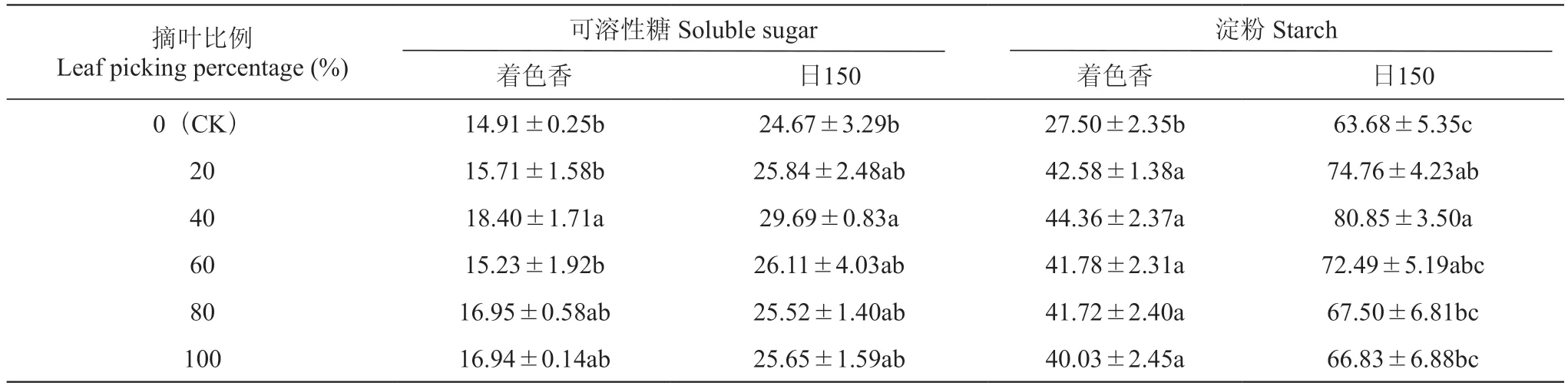
表2 摘叶处理对葡萄枝条可溶性糖和淀粉含量的影响Table 2 Effects of leaf picking treatment on content of soluble sugar and starch in grape branches mg·g-1
2.2 摘叶对枝条氨基酸、可溶性蛋白含量的影响
由表3可知,摘叶处理后枝条中氨基酸含量均升高。对于‘着色香’,摘叶40%和全摘叶处理后枝条氨基酸含量显著高于CK,分别理增加70.54%和56.58%;对于‘日150’,摘叶20%、40%、60%处理后枝条氨基酸含量均显著高于CK,分别增加25.91%、46.36%和30.91%。摘叶处理后枝条内可溶性蛋白含量均降低,‘着色香’摘叶40%处理后枝条可溶性蛋白含量显著低于CK22.47%。‘日150’摘叶20%、40%、60%处理后枝条可溶性蛋白含量均显著低于CK,分别降低18.24%、31.23%和19.66%。‘着色香’摘叶40%处理后枝条脯氨酸含量最高,为29.98 mg·g-1,显著高于CK;‘日150’摘叶20%、40%、60%处理后枝条脯氨酸含量与CK差异显著,分别增加19.25%、35.46%和18.68%。
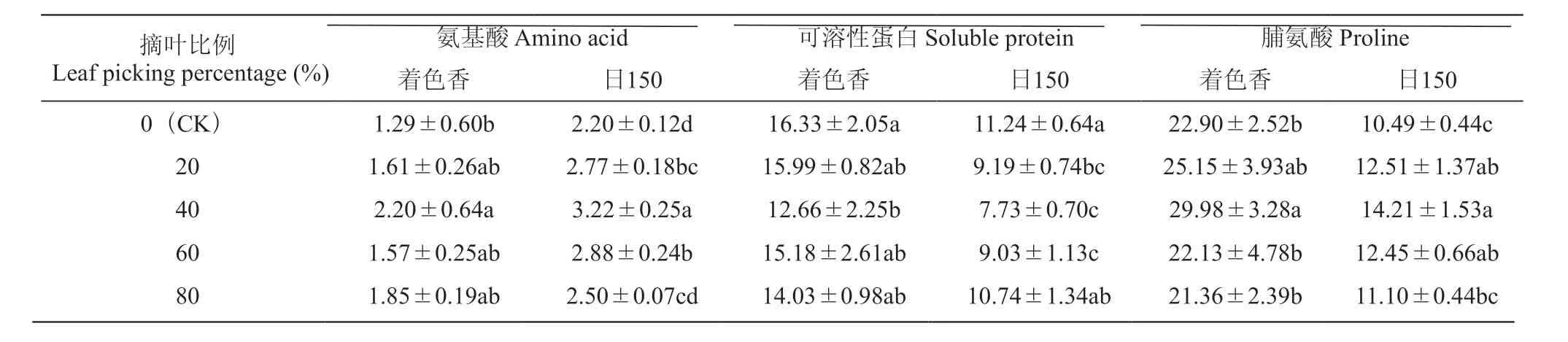
表3 摘叶处理对葡萄枝条氨基酸、可溶性蛋白和脯氨酸含量的影响Table 3 Effects of leaf picking treatment on content of aminoacid, soluble protein and proline in grape branches mg·g-1
2.3 摘叶对枝条POD和SOD活性的影响
由表4可知,摘叶后枝条内POD活性均升高。‘着色香’摘叶20%、40%、60%处理的枝条POD活性均显著高于CK;摘叶40%处理枝条POD活性最高,比CK增加146.654%;摘叶20%和摘叶60%处理分别增加134.99%和56.53%。‘日150’摘叶40%处理枝条POD活性显著高于CK 75.00%。摘叶后枝条内SOD活性均有所下降,‘着色香’摘叶20%、40%、60%处理后枝条SOD活性与CK差异显著,分别降低14.43%、20.42%、15.54%;‘日150’摘叶40%处理枝条SOD活性显著降低,较CK降低17.54%。
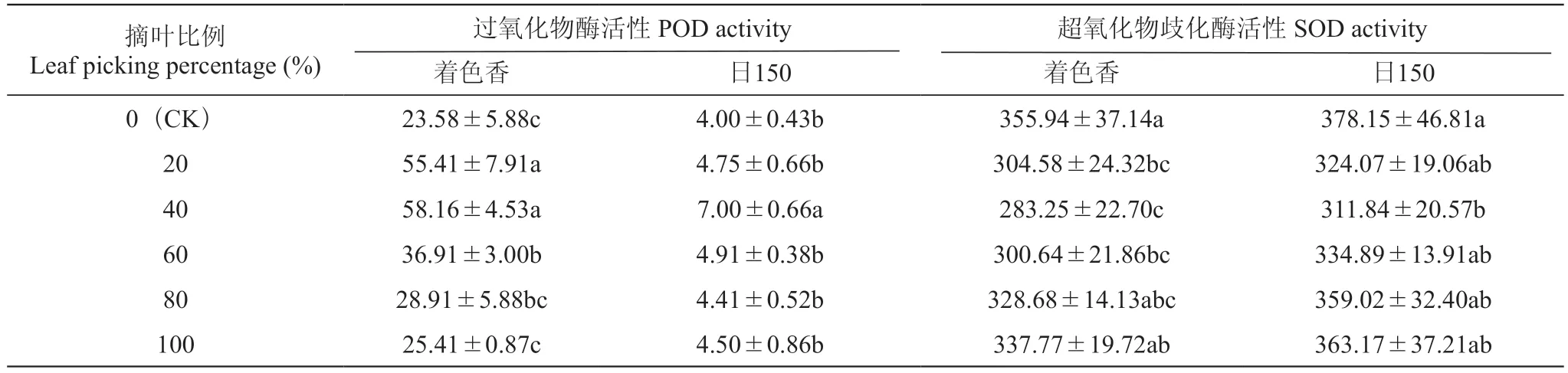
表4 摘叶处理对葡萄枝条POD和SOD活性的影响Table 4 Effects of leaf picking treatment on POD and SOD activity in grapevine branchesU·g-1
2.4 五个关键氮素基因在枝条上的半定量表达分析
由图1所示,‘着色香’谷氨酰胺合成酶基因(VvGS)在摘叶20%处理下的表达量下调,天冬酰胺合成酶基因(VvAS)在摘叶20%、60%和全摘叶处理下的表达量下调,硝酸还原酶基因(VvNR)在摘叶40%和摘叶60%处理下的表达量下调,亚硝酸还原酶基因(VvNiR)在摘叶40%处理下的表达量下调,谷氨酸脱氢酶基因(VvGDH)在各处理下的表达量差异不明显。

图1 五个氮代谢关键基因在葡萄枝条上的半定量表达结果Figure1 Semi-quantitative expression results of five key genes of nitrogen metabolism on grape branches
‘日150’谷氨酰胺合成酶基因(VvGS)在各处理下的表达量变化不明显,天冬酰胺合成酶基因(VvAS)、硝酸还原酶基因(VvNR)、谷氨酸脱氢酶基因(VvGDH)在摘叶40%处理下的表达量均下调,亚硝酸还原酶基因(VvNiR)在摘叶40%和摘叶60%处理下的表达量下调。总体来看,5个氮代谢关键基因在各处理的表达量存在明显差异,摘叶40%和摘叶60%处理下,多数基因的表达量下调明显,说明一定数量的摘叶处理能降低枝条内氮素的同化和固定速率,诱导葡萄进入休眠。
2.5 GA合成酶相关基因、ABA合成酶相关基因在枝条上的半定量表达分析
由图2可知,‘着色香’贝壳杉烯氧化酶基因(VvKO)在各处理下基因表达量变化不明显,GA3β-羟化酶基因(VvGA3ox4)在各处理的表达量均下调;9-顺式环氧类胡萝卜素双加氧酶(VvNCED1)在摘叶20%、60%、80%和全摘叶处理下表达量均上调。
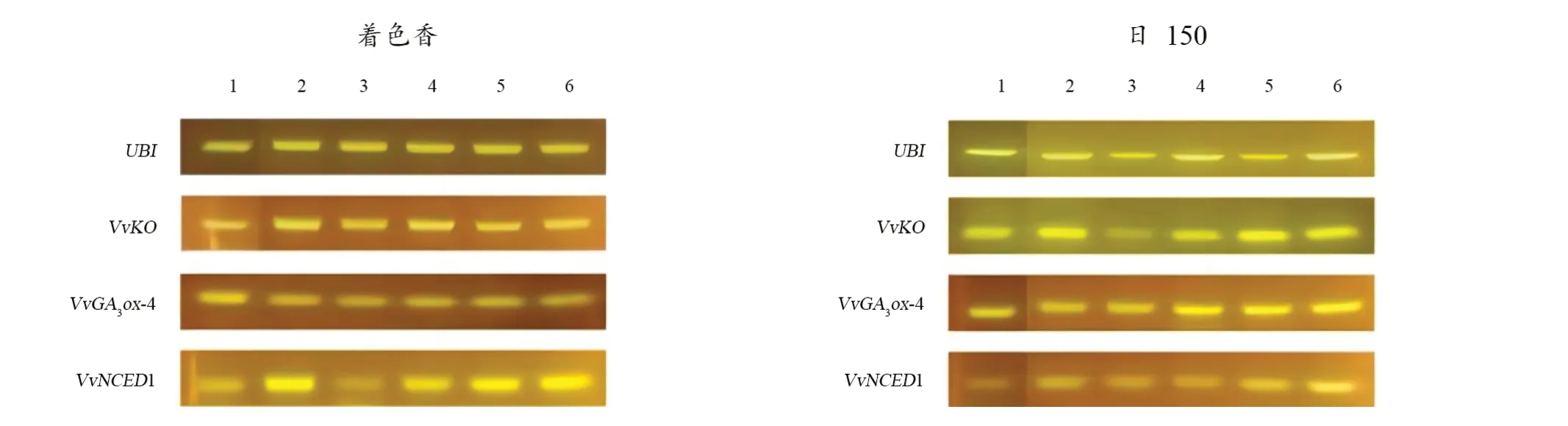
图2 GA合成酶、ABA合成酶相关基因在葡萄枝条上的半定量表达结果Figure 2 Semi-quantitative expression results of genes related to GA synthesis and ABA synthesis on grape
‘日150’贝壳杉烯氧化酶基因(VvKO)在摘叶40%处理下表达量下调,GA3β-羟化酶基因(VvGA3ox4)在摘叶20%和摘叶40%处理下表达量下调;9-顺式环氧类胡萝卜素双加氧酶(VvNCED1)在各处理下的表达量均上调。总体上,摘叶40%处理下,GA合成酶相关基因表达量降低明显,而ABA合成酶相关基因表达量在各处理下均有所上调,说明40%摘叶处理可有效抑制植株的营养生长,减缓生理代谢速率,诱导葡萄进入休眠。
3 讨论
3.1 养分物质含量与休眠
休眠期糖类物质代谢与休眠进程密切相关,影响休眠的因素会引起糖类物质的变化[4]。糖类物质不仅能为植物提供能量,维持植株的正常生理代谢,还能增强植株的渗透作用,降低植株细胞的冰点,并增强植株对不良环境的抵抗力。在枣、酥梨和油桃上的研究结果表明,在自然休眠过程中,枝芽内可溶性糖含量逐渐增加[13-15]。本研究中,摘叶40%处理下葡萄枝条的可溶性糖和淀粉含量均显著升高,有利于植株进入休眠。
落叶果树休眠期间可溶性蛋白和游离氨基酸含量一般呈负相关,这种变化是由可溶性蛋白与游离氨基酸之间互相转化形成的[16]。有研究表明,枝条内的脯氨酸含量随着环境温度的下降而升高[17]。李芙等[18]对不同葡萄品种的研究表明,随着可溶性蛋白含量的显著降低,葡萄花芽的休眠进一步加深。本研究中,摘叶40%处理下葡萄枝条中可溶性蛋白含量显著降低,氨基酸和脯氨酸含量显著增加,有利于葡萄植株提前进入休眠。
3.2 抗逆物质活性与休眠
研究表明,植物的氧化还原可平衡调节芽端分生组织的活性,直接或间接影响芽休眠[19-22]。H2O2作为传递休眠的信号物质,与其密切关联的抗氧化酶POD和SOD,在果树休眠中起重要作用[23-24]。SOD可催化O2-形成H2O2,POD则将H2O2分解成O2和H2O[25-26]。高东升等[27]以油桃和蜜桃为试材,在自然落叶前20 d用脱叶剂处理,促进了自然休眠进程的发生,并提早解除自然休眠,期间SOD活性呈下降趋势,POD活性一直升高。本试验结果表明,摘叶40%处理下葡萄枝条的POD活性显著升高,SOD活性显著下降,这有利于H2O2维持在一定水平,在促进休眠的同时,避免细胞膜脂发生过氧化反应,与前人研究结果相一致。
3.3 与休眠相关基因的表达
氮是植物体内需求量最大的矿质营养元素,影响植物正常生长发育,是植物必需的营养元素之一[28]。铵态氮和硝态氮是植物生长发育过程中的主要氮源,其中被植物体吸收的硝态氮需要在硝酸还原酶(NR)、亚硝酸还原酶(NiR)等的作用下,转化成NH3,再借助谷氨酰胺合成酶(GS)进行氨同化,合成蛋白质[29]。这其中,天冬酰胺则作为有机氮运输和储存的主要形式[30]。本试验摘叶40%处理后,多数氮代谢相关基因表达量下调,说明40%摘叶处理使植株内氮的同化和固定速率降低,营养生长减弱,生理代谢速率降低。
GA和ABA同属植物激素,通过信号传导来调控植物的各项生长发育[31],并且,GA和ABA含量与植物的休眠进程密切相关[32-33]。有研究表明,ABA能促进芽的休眠,并且间接地抑制GA20ox基因的表达[34]。葛会波等[35]研究证明,GA含量随着休眠加深而逐渐降低,且GA除通过自身信号通路来调控休眠外,还与ABA信号通路相互影响。本试验中,摘叶40%处理下GA合成相关基因表达量下调,ABA合成相关基因表达量上调,说明葡萄采收后摘叶40%处理能抑制植株的营养生长、减缓生理代谢速率,促进葡萄进入休眠。
4 结论与展望
在公主岭市,‘着色香’和‘日150’葡萄果实采收后摘叶40%处理,能有效促进植株内营养物质的积累,改变抗氧化酶活性,有效地抑制膜脂过氧化,有助于葡萄植株提前进入休眠。
关于摘叶诱导葡萄休眠的研究报道相对较少,研究也不够深入。在未来的研究中,应进一步利用高通量测序以及高效液相色谱等技术,开展转录组和代谢组学联合分析,结合内源激素含量变化,挖掘调控葡萄休眠相关的重要基因,解析葡萄休眠的代谢网络途径,为葡萄休眠机理研究及诱导休眠技术的应用奠定理论基础。
