不同钾水平对毛竹幼苗生长及生理指标的影响
2023-03-15何榜眼唐伟华黄大勇刘世男
何榜眼,唐伟华,黄大勇,刘世男
(1.广西大学 林学院,广西 南宁 530004;2.广西壮族自治区林业科学研究院,广西 南宁 530004)
钾是植物生长发育必需的营养元素之一,对植物酶的激活、光合作用、养分分配和渗透调节等具有重要影响[1-2]。第二次全国土壤普查结果显示,我国约有1/4~1/3的土壤处于一般缺钾或严重缺钾状态,在长江以南地区则更为普遍[3]。土壤钾素缺乏已成为限制作物生长发育的主要因素之一。研究表明,适量的钾能有效促进植物生长,缺钾或过量钾会对植物的生长发育产生抑制作用[4-5]。张永清等[6]指出,缺钾时小麦Triticum aestivum的株高、叶面积、根尖数、总根长、总表面积均降低。Fontana等[7]研究表明,钾素不足会引起棉花Gossypium hirsutum的株高、叶面积和根表面积减小,分枝和根的伸长受阻,导致全株的生物量降低;安琼等[8]对玉米Zea Mays、周文灵等[9]对甘蔗Saccharum officinarum的研究均表明,缺钾会导致植株株高和生物量下降,使叶片丙二醛(MDA)含量上升,并诱导超氧化物歧化酶(SOD)活性提升。陈利娜等[10]指出钾素过低或过高对黑木相思Acacia melanoxylon根系生长促进作用不大,钾素过量还会限制根系的生长。花生Arachis hypogaea在供钾素过量时会导致叶面积指数减小以及产量下降[11]。可见,适当的钾水平对植株的生长发育和产量形成尤为重要。
毛竹Phyllostachys edulis为禾本科刚竹属,是国内重要的经济竹种,其竹材可用于造纸、建筑、家具等多种用途,竹笋还可当食用蔬菜。毛竹生长迅速,需要从土壤中吸取大量的钾才能正常地生长。然而我国毛竹多分布在南方地区,其产地普遍存在缺钾的现象,加之经营方式多以粗放型为主,导致毛竹林地钾素愈加缺乏,严重制约着毛竹的产量和品质的提高[12]。目前,关于毛竹养分管理方面的研究已有不少报道,如磷、氮添加[13-14]、氮磷钾配比[15]、复合肥和有机无机复混肥施用[16-17],以及生物肥开发[12,18]等方面,而有关供钾量对毛竹生长和生理方面的效应尚未明确。基于此,本研究以毛竹实生苗为对象,研究不同钾水平对毛竹幼苗相关生长和生理指标的影响,以期为毛竹苗期钾素营养的调控提供参考。
1 材料与方法
1.1 材料培养与试验设计
试验所用毛竹种子由广西桂林市林业科学研究所提供。选取均匀饱满的种子置于75%的酒精中灭菌5 min,然后用纯水将种子清洗6~8次,在25℃下纯水浸泡36 h,之后摆放至培养皿中进行萌发。待种子萌发后将其移至育苗托盘,培养至幼苗子叶展开,挑选苗高为2.42±0.22 cm,地径为0.40±0.03 mm的毛竹幼苗移至长宽高分别为127 mm×87 mm×114 mm的96孔水培盒中用不同钾水平的营养液培养。营养液的钾源为K2SO4,其他营养元素浓度保持一致(营养液配方见表1)。培养过程始终在恒温箱内进行,恒温箱内光照和黑暗的时长分别为14 h和10 h,光照强度为10 000 lx,温度为25℃。试验采用完全随机设计,共设置6个钾水平,分别为0 (CK)、0.005 (K1)、0.01(K2)、0.1 (K3)、1 (K4)和2 mmol/L (K5)。每个钾水平设5个水培盒,每盒培养24株毛竹幼苗。培养期间72 h更换一次营养液,处理15 d后对其生长指标进行测定,然后剪取生长位置和成熟度一致的毛竹幼苗鲜叶于冰箱中-20℃下保存,用于叶片生理指标的测定,另外再取一部分植株于105℃杀青30 min,接着在75℃下烘至恒质量,用于氮、磷、钾含量的测定。
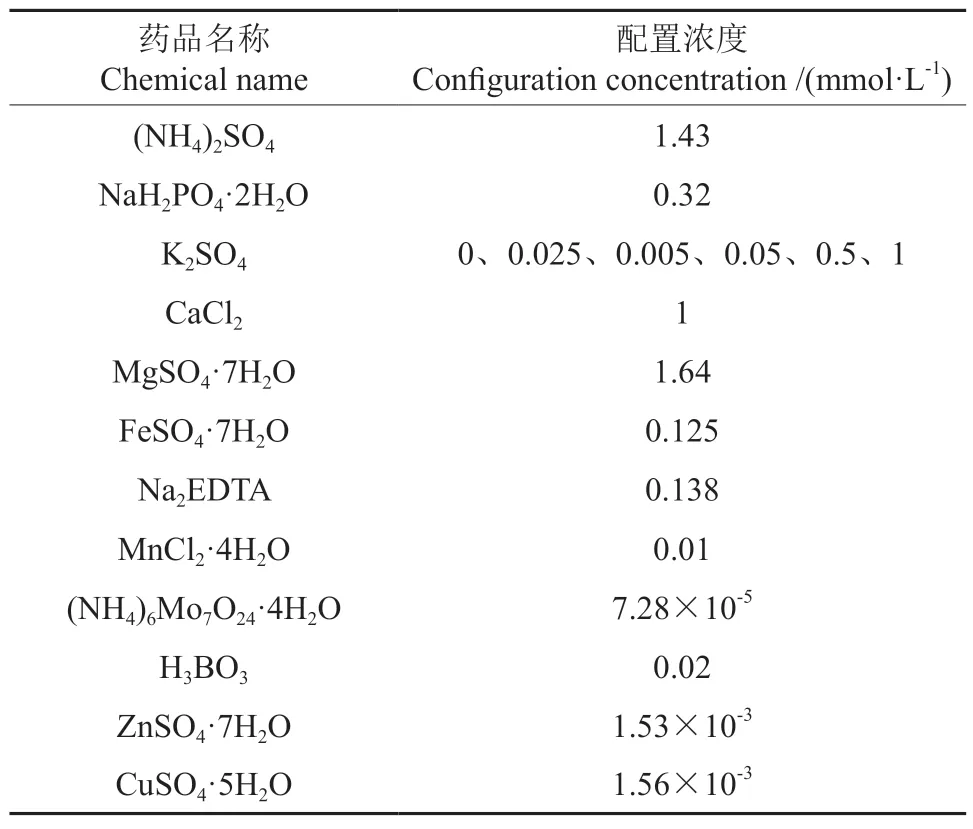
表1 营养液成分及配置浓度Table 1 Nutrient solution composition and configuration concentration
1.2 测定指标及方法
1.2.1 生长指标测定
每个处理随机选取毛竹幼苗30株,使用钢尺测量其苗高、节间距和主根长,用游标卡尺测量地径,以YMJ—B叶面积测量仪测定叶长、叶宽和叶面积,使用Epson全自动根系扫描仪进行根系图像扫描,用Win-RHIZO 2012软件分析根系图像,获得根系总根长、总根表面积、平均根径和总根体积。通过观察对根尖进行计数。
1.2.2 生理指标测定
超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法测定;过氧化物酶(POD)采用愈创木酚法测定;多酚氧化酶(PPO)活性采用邻苯二酚法测定;丙二醛(MDA)含量采用硫代巴比妥酸法测定;可溶性蛋白含量采用考马斯亮蓝G—250法测定;可溶性糖含量采用蒽酮比色法测定[19]。养分含量的测定参照土壤农业化学分析方法[20],将烘干的样品研磨过0.25 mm孔径筛,取0.1 g经H2SO4-H2O2消煮后,氮含量采用靛酚蓝比色法测定,磷含量采用钼锑抗比色法测定,钾含量采用火焰光度计测定。
1.3 数据统计与分析
采用Excel 2019软件进行数据整理和制图,采用SPSS 24.0软件对试验数据进行单因素方差分析(ANOVA),以Duncan’s新复极差法分析不同处理的差异显著性,以Pearson相关系数分析不同指标间的相关性。
2 结果与分析
2.1 不同钾水平对毛竹幼苗地上部分生长指标的影响
由表2可以看出,随着钾水平的下降,毛竹幼苗的苗高、地径、叶长、叶宽、叶面积和平均节间距均呈先上升后下降的趋势。其中,在钾水平低于或高于K4时,幼苗的苗高、地径和平均节间距均显著降低;在钾水平低于K3时,幼苗的叶长、叶宽和叶面积均显著降低,而K3、K4和K5之间幼苗叶长、叶宽和叶面积的差异则均不显著。就高径比而言,K3、K4和K5水平均显著大于CK、K1和K2水平,K3、K4和K5之间的差异则不显著。而不同钾水平对幼苗叶长宽比无显著影响。总体上看,K4水平对毛竹幼苗地上部分生长的促进效果最佳。

表2 各钾水平毛竹幼苗地上部分生长状况对比†Table 2 Comparison of growth status of the aboveground part of Phyllostachys edulis seedlings treated with different potassium levels
2.2 不同钾水平对毛竹幼苗根系生长指标的影响
如表3所示,随着钾水平的下降,毛竹幼苗总根长、主根长和根尖数均呈先上升后下降的趋势。其中,在钾水平低于K4时,幼苗的总根长、主根长和根尖数均显著降低(P<0.05),在钾水平高于K4时,幼苗主根长和根尖数显著降低(P<0.05)。不同钾水平中,K4水平幼苗的总根表面积和总根体积均显著大于CK、K1、K2和K3水平(P<0.05),K4和K5水平之间的差异则不显著。而不同钾水平对幼苗的平均根径无显著影响。总体上看,K4水平对毛竹幼苗根系生长的促进效果最佳。

表3 各钾水平毛竹幼苗根系生长状况对比Table 3 Comparison of growth status of the root of Phyllostachys edulis seedlings treated with different potassium levels
2.3 不同钾水平对毛竹幼苗叶片保护酶活性和MDA含量的影响
如图1所示,随着钾水平的降低,毛竹幼苗叶片MDA含量呈上升趋势,其中,CK、K1、K2和K3、K5水平的差异均为显著(P<0.05),而K4和K5之间的差异则不显著。SOD活性随着钾水平的降低表现为先下降后上升的趋势,其中,活性最高和最低的分别为CK和K4水平,二者与其他钾水平的差异性均达到显著水平(P<0.05);而POD活性随着钾水平的降低表现为先上升后下降的趋势,其中,CK、K1、K2和K3水平较K4、K5水平的POD活性均显著提高(P<0.05),K4和K5之间的差异则不显著;PPO活性随钾水平下降则呈“降—升—降”的趋势,其中,PPO活性最高和最低的分别为K1和K4水平,二者与其他钾水平的差异性均达到显著水平(P<0.05)。

图1 各钾水平毛竹幼苗叶片保护酶活性和MDA含量对比Fig.1 Comparison of the protective enzyme activities and MDA content in leaves of Phyllostachys edulis seedlings treated with different potassium levels
2.4 不同钾水平对毛竹幼苗叶片可溶性蛋白和可溶性糖含量的影响
由图2可看出,随着钾水平的降低,毛竹幼苗叶片可溶性蛋白含量总体呈上升趋势,可溶性糖含量则呈先上升后下降的趋势。其中,CK、K1和K2水平幼苗叶片可溶性蛋白含量较K3、K4和K5水平均显著上升(P<0.05),而K3、K4和K5之间的差异则不显著;CK至K3水平幼苗叶片可溶性糖含量较K4和K5水平均显著提升(P<0.05),而K4和K5之间的差异则不显著。

图2 各钾水平毛竹幼苗叶片可溶性蛋白和可溶性糖含量对比Fig.2 Comparison of the soluble protein and soluble sugar contents in leaves of Phyllostachys edulis seedlings treated with different potassium levels
2.5 不同钾水平对毛竹幼苗各部位氮含量的影响
如图3所示,不同钾水平下毛竹幼苗根、茎、叶和全株氮含量均存在显著差异(P<0.05)。其中,CK、K1、K2和K3水平毛竹幼苗根、茎、叶以及全株的氮含量均显著高于K4和K5水平(P<0.05),而K4和K5水平之间根、茎、叶以及全株的氮含量差异则不显著。在各器官中,不同钾水平下毛竹幼苗叶的氮含量均明显高于根和茎。

图3 不同钾水平毛竹幼苗各部位N含量对比Fig.3 Comparison of the N content in different parts of Phyllostachys edulis seedlings treated with different potassium levels
2.6 不同钾水平对毛竹幼苗各部位磷含量的影响
由图4可以看出,随着钾水平降低,毛竹幼苗根和全株磷含量的变化均呈先降后升的趋势。其中,K5水平根部磷含量最高且与其他钾水平差异均显著(P<0.05),K1水平磷含量最低,与CK、K3、K4和K5水平均差异显著(P<0.05),与K2水平的差异则不显著;就全株而言,K5水平磷含量显著高于其他钾水平(P<0.05),CK和K4水平磷含量显著高于K1、K2和K3水平(P<0.05),而K1、K2和K3水平间磷含量的差异不显著。从茎的磷含量来看,K4水平显著高于其他钾水平(P<0.05),CK、K1和K5水平显著高于K3水平,而K2与K3之间的差异则不显著。从叶的磷含量来看,K4水平显著低于CK、K1、K2和K5水 平(P<0.05),CK、K1、K2、K3和K5之间的差异则不显著。在各器官中,不同钾水平下毛竹幼苗根的磷含量均明显高于茎和叶。
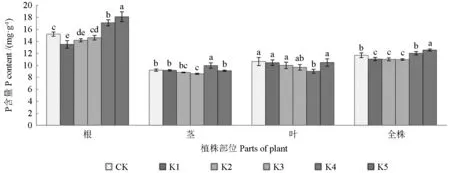
图4 不同钾水平毛竹幼苗各部位P含量对比Fig.4 Comparison of the P content in different parts of Phyllostachys edulis seedlings treated with different potassium levels
2.7 不同钾水平对毛竹幼苗各部位钾含量的影响
由图5可以看出,随着钾水平降低,毛竹幼苗根、叶以及全株的钾含量均呈下降趋势。其中,K2、K3、K4和K5水平根和叶的钾含量较CK均显著上升(P<0.05),而K1水平根和叶的钾含量与CK的差异均不显著;CK、K1、K2和K3水平全株的钾含量较K4和K5均显著降低(P<0.05),而K4和K5之间全株的钾含量差异则不显著。就茎而言,其钾含量随着钾水平下降呈先升后降的趋势,其中,钾含量最低和最高者分别为CK和K4水平,二者与其他钾水平间的差异均为显著(P<0.05)。

图5 不同钾水平毛竹幼苗各部位K含量对比Fig.5 Comparison of the K content in different parts of Phyllostachys edulis seedlings treated with different potassium levels
2.8 钾水平、生长指标、叶片生理指标间的相关分析
如表4所示,钾水平与MDA含量和可溶性糖含量显著负相关。苗高与地径、主根长显著正相关,与叶面积、平均节间距、总根长、总根表面积、总根体积和根尖数极显著正相关,与可溶性蛋白含量极显著负相关。地径与平均节间距、总根长、主根长、总根表面积、总根体积显著正相关,与根尖数极显著正相关。叶面积与平均节间距、总根长和根尖数极显著正相关,与主根长、总根表面积和总根体积显著正相关,与MDA含量显著负相关,与可溶性蛋白含量极显著负相关。平均节间距与总根长、主根长、总根表面积、总根体积和根尖数极显著正相关,与MDA含量和可溶性糖含量显著负相关,与可溶性蛋白含量极显著负相关。总根长与主根长、总根表面积、总根体积和根尖数极显著正相关,与MDA含量和可溶性糖含量显著负相关,与可溶性蛋白含量极显著负相关。主根长与总根表面积、总根体积和根尖数极显著正相关,与MDA含量和可溶性蛋白含量显著负相关。总根表面积与总根体积和根尖数极显著正相关,与MDA含量、可溶性蛋白含量和可溶性糖含量显著负相关。总根体积与根尖数极显著正相关,与MDA含量和可溶性蛋白含量显著负相关。根尖数与可溶性蛋白含量显著负相关。MDA含量与可溶性蛋白含量和可溶性糖含量显著正相关。
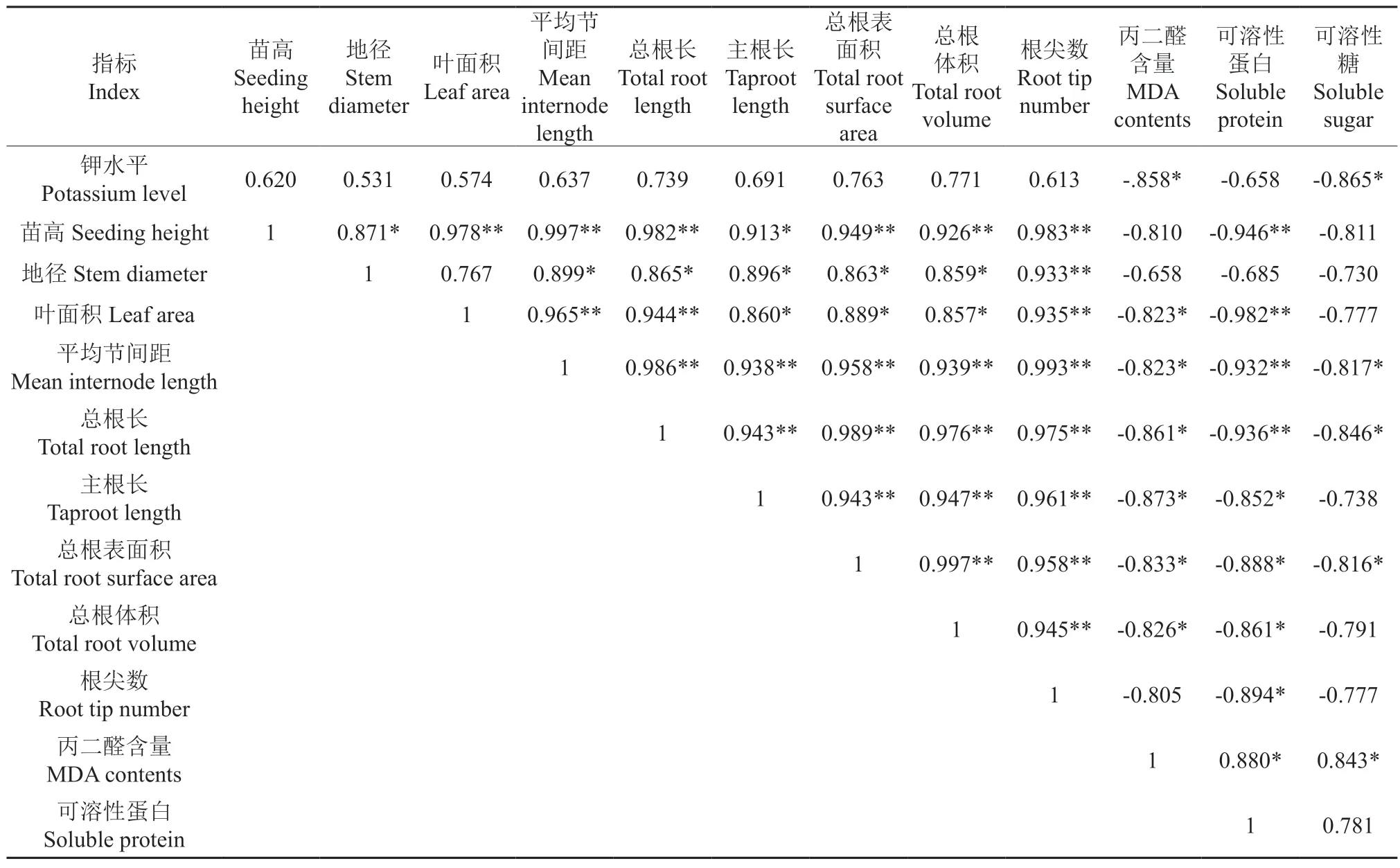
表4 钾水平、生长指标、叶片生理指标间的相关系数†Table 4 Correlation coefficients between potassium level, growth index and leaf physiological index
3 讨 论
除种子萌发时期自身供给外,植物所需的钾绝大部分是从环境中获取的,环境中钾的含量在很大程度上影响着植物的生长发育。刘晓伟等[21]研究发现,低钾会引起小麦叶长、叶面积、株高、总根长、总根表面积等形态指标降低。刘芳等[22]研究表明,缺钾会显著降低香蕉Musa acuminata的总根长、总根表面积和总根体积。高伟勤等[23]研究发现,无钾、低钾和高钾均会降低枳Poncirus trifoliata的株高、茎粗、叶片数以及叶、茎和根的生物量。本研究中,供钾水平低于1 mmol/L时,毛竹幼苗的苗高、地径、平均节间距、总根长、主根长、总根表面积、总根体积和根尖数均显著降低(P<0.05),高于1 mmol/L时幼苗的苗高、地径、平均节间距、主根长和根尖数均显著降低(P<0.05)。表明钾水平过低和过高均会限制毛竹幼苗的生长,这与以往的研究结果一致。
在正常情况下,植物细胞内活性氧(ROS)的产生与清除处于动态平衡状态,当遭受逆境时,该平衡会被打破,细胞内产生大量的ROS,如果产生的ROS不能被有效地清除,会导致膜脂过氧化,最终产生MDA[24-25]。在一定含量范围内,可通过测定MDA含量来判断植物膜脂过氧化的程度[26]。本研究中,随着钾水平的下降,毛竹幼苗叶片MDA含量呈上升趋势,说明钾水平下降会加剧毛竹幼苗叶片膜脂过氧化程度,该结果与夏乐[27]对玉米、杨日才[28]对香蕉的研究结果一致。为防止膜脂过氧化,植物演变出了一套高效的酶促清除活性氧系统,可通过增加体内SOD、POD和CAT等保护酶活性来清除ROS,减少生物膜的损伤[29-30]。况帅等[31]对烟草Nicotiana tabacum、Miao等[32]对大豆Glycine max(L.)Merr.的研究均表明,低钾会引起植株体内MDA含量上升,使SOD、CAT和POD活性显著增加。本研究中,当钾水平低于1 mmol/L时毛竹幼苗叶片SOD、POD和PPO的活性均显著增加(P<0.05),高于1 mmol/L时仅SOD和PPO活性显著增加(P<0.05)。说明毛竹幼苗各保护酶间的协调表达因钾水平的变化而异。推测该差异可能是因为毛竹幼苗为应对不同程度活性氧毒害而采取了不同措施。随着钾水平下降MDA含量不断上升,说明钾越缺乏毛竹幼苗叶片受活性氧毒害越严重,此时多种酶的协调表达可能更利于其清除过量的活性氧,以防止膜脂过氧化加剧。
一般情况下,K+是植物调节细胞膨压的主要物质,对维持细胞正常生理功能具有重要作用[33]。K+缺乏时会引起细胞膨压下降,导致生物膜和细胞器受损,最终扰乱细胞正常的代谢活动。为维持细胞正常生理功能,植物可通过改变其他渗透调节物质的含量来调节细胞膨压[34]。可溶性糖和可溶性蛋白是植物体内重要的渗透调节物质,其含量变化与植物的抗逆性密切相关[35]。缺钾条件下,不同植物种类体内可溶性糖和可溶性蛋白含量的变化存在差异。吴敏等[36]对巴西橡胶Hevea brasiliensis、闫洪奎等[37]对玉米的研究均表明,缺钾时植株叶片的可溶性蛋白含量降低,可溶性糖含量则上升。黄海云[38]对花生、饶宝蓉等[39]对香蕉的研究则表明,缺钾会引起植株叶片可溶性蛋白含量提升,李兴涛等[40]研究表明,缺钾会降低大豆叶片可溶性糖含量。本研究中,当钾水平为0、0.005和0.01 mmol/L时,毛竹幼苗叶片可溶性蛋白含量较0.1、1和2 mmol/L的显著上升;钾水平为0~0.1 mmol/L时,幼苗叶片可溶性糖含量较1 mmol/L和2 mmol/L的显著提升。这表明增加叶片可溶性蛋白和可溶性糖的含量是毛竹幼苗防止钾缺乏导致细胞膨压下降的一种重要方式。
植物对钾、氮和磷的吸收存在一定的互作效应[41-42]。在供钾量变化时,因不同植物偏好不同,其体内氮、磷、钾含量的变化存在差异。刘晓伟等[21]对小麦的研究表明,低钾处理会导致其地上部和根的氮、磷、钾含量降低;盆栽试验表明,缺钾显著提高了香蕉地上部、根系和全株的氮、磷含量,而钾含量则显著降低[22];习金根等[43]研究发现,施钾显著提高了剑麻Agave sisalanalPerr.ex Englem地上部氮、磷、钾含量和根系的钾含量,而根氮含量显著降低。本研究中,当钾水平为0~0.1 mmol/L时,毛竹幼苗根、茎、叶以及全株的钾含量均显著低于1或2 mmol/L的钾含量,但根、茎、叶以及全株的氮含量均显著增加。表明缺钾会使毛竹幼苗体内钾含量相应减少,这与以往的研究结果一致。而钾水平下降导致植株氮含量增加,可能是低钾条件下毛竹幼苗受渗透胁迫时可溶性蛋白含量增加的结果,或是缺钾时K+与NH4+之间的拮抗作用减弱促使毛竹幼苗增加对营养液中NH4+的吸收。另外,钾水平下降还改变了毛竹幼苗体内的磷含量,主要表现为根系和全株磷含量明显下降,说明高钾水平对毛竹幼苗磷的吸收具有促进作用,但茎和叶中的磷含量变化规律并不明显,可能是毛竹幼苗为满足地上部的生长需求将磷优先向其供应所致。
有研究表明,不同钾水平下植物生长指标和生理指标之间存在一定的相关性[44]。罗汉东等[45]指出,不同供钾水平下油茶Camellia oleifera的地径与株高呈极显著正相关(P<0.01)。赵泽茹等[46]对烟草的研究表明,钾水平与可溶性糖含量的相关性不显著,而与可溶性蛋白含量极显著正相关(P<0.01)。本研究中,钾水平与毛竹幼苗叶片MDA和可溶性糖含量呈显著负相关(P<0.05),即钾水平下降会诱导毛竹幼苗叶片MDA和可溶性糖含量增加,说明缺钾会对毛竹幼苗活性氧代谢和渗透调节产生影响。在前述生长指标中,除叶面积与地径的相关性未达显著水平外,其余生长指标两两之间呈显著正相关(P<0.05)或极显著正相关(P<0.01),表明毛竹幼苗根、茎、叶的生长对钾水平变化的响应具有一致性。叶片MDA含量与叶面积、平均节间距、总根长、主根长、总根表面积和总根体积均显著负相关(P<0.05);可溶性蛋白含量与苗高、叶面积、平均节间距和总根长极显著负相关(P<0.01),与主根长、总根表面积、总根体积和根尖数显著负相关(P<0.05);可溶性糖含量与平均节间距、总根长和总根表面积显著负相关(P<0.05)。上述结果说明钾缺乏引起叶片MDA、可溶性蛋白和可溶性糖含量的增加,会限制毛竹幼苗根、茎和叶的生长,推测是MDA对叶片的毒害以及光合产物向可溶性蛋白和可溶性糖的转化使物质积累减少,限制了幼苗叶形态的建成,这在一定程度上减弱了幼苗的光合能力,进而影响茎和根的生长。
植物的生长发育以养分的吸收利用为基础,研究植物钾素营养状况与植株生长以及氮、磷、钾吸收利用的关系,可在一定程度上为植物的养分管理提供指导。本研究在毛竹幼苗期考察不同钾水平对其生长和生理生化方面的影响,对毛竹实生苗的钾素管理有一定的参考价值,但由于试验是通过水培的方式进行,其研究结果与实际生产应用仍存在一些差距。此外,本试验仅对毛竹苗期对钾素的需求规律和相应效应进行探讨,而关于毛竹其他生长阶段对钾素的需求特征、不同供钾量毛竹体内养分吸收利用的动态变化、低钾胁迫下毛竹的分子响应机理等目前仍未明朗,这一系列问题将是下一步的研究方向。
4 结 论
本研究表明,在一定范围内钾水平的上升有利于促进毛竹幼苗地上部分和根系形态的建成,但超过该范围时继续添钾对毛竹幼苗的促长效益不大,甚至会抑制其生长。本试验条件下,钾水平为1 mmol/L时毛竹幼苗地上部分和根系的生长状况最佳。供钾不足会导致毛竹幼苗叶片MDA含量积累并对其造成渗透胁迫,严重影响叶片的生理健康;但毛竹幼苗可通过叶片各项保护酶的协同作用和增加其他有机溶质的含量来减轻缺钾引发的负面效应。缺钾还会影响毛竹幼苗体内氮、磷、钾的平衡,主要表现为缺钾时根、茎、叶以及全株的氮含量升高,而根、茎、叶以及全株的钾含量降低,并且根的磷含量也降低。可见适宜的钾水平不仅能促进毛竹生长,还能改善植株的生理状况。基于此,建议在生产实践中注意合理适量地对毛竹施用钾肥,既能避免钾素资源浪费,还能达到增产和增效的目的。
