乌虾酱长期发酵过程中细菌群落演替及对生物胺形成的影响
2023-01-07邓义佳王润东张宇昊李学鹏季广仁励建荣
邓义佳,王润东,张宇昊,李学鹏,季广仁,励建荣,*
(1.西南大学食品科学学院,重庆 400715;2.渤海大学食品科学与工程学院,辽宁 锦州 121013;3.川渝共建特色食品重庆市重点实验室,重庆 400715;4.锦州笔架山食品有限公司,辽宁 锦州 121007)
虾酱又名“虾膏”,是我国沿海地区、新加坡和马来西亚等东南亚国家常用的调味料,以乌虾、蜢子虾和对虾加工副产品为原料,添加15%~25%的食盐自然发酵而成[1-2]。虾酱的品质与发酵周期密切相关,随发酵时间延长,虾酱形态和风味逐渐优化,经过3 a以上的长期发酵,虾酱可呈现均匀的深褐色糊状,有醇类、醛类和酯类等风味物质形成,具有独特口感和极佳风味[3-5]。
微生物群落是虾酱品质提升的关键因素,分离菌株如乳杆菌属(Lactobacillus)、交替假单胞菌(Pseudoalteromonas)和葡萄球菌属(Staphylococcus)促进蛋白质分解形成呈味氨基酸,增加虾酱鲜味[6],调节产酸细菌如腐生葡萄球菌(S. saprophyticus)、木糖葡萄球菌(S. xylosus)和格氏乳球菌(Lactococcus garvieae)相对丰度能够抑制虾酱腥臭和氨味形成[7]。然而,自然发酵虾酱中微生物群落复杂多样,通过纯培养识别的微生物种类仅占实际种群总量的极少部分,且目前仅针对短期发酵虾酱中的细菌多样性进行探索[8-9],对于长期发酵过程中细菌群落演替规律尚不清楚。此外,发酵食品中普遍存在脱羧酶阳性菌群,它们能够转化食品中的游离氨基酸,降解为腐胺、尸胺、组胺、精胺和亚精胺等生物胺。消费者误食过量的生物胺,产生头痛、恶心和痉挛等症状,造成严重的食品安全隐患[10]。研究发现,肠球菌属(Enterococcus)、乳杆菌属和乳球菌属是豆腐乳中组胺合成菌群[11],假单胞菌(Pseudomonas)、弧菌(Vibrio)和葡萄球菌与腌制海鲈鱼中尸胺积累密切相关。近年来,国内外学者指出虾酱中也存在生物胺超标问题,即使添加茶多酚等抑胺剂也无法降低其含量[12-13]。因此,阐明虾酱长期发酵过程中细菌群落演替与生物胺的关联、挖掘关键菌群成为解决虾酱产品中生物胺超标问题的新策略。
本研究以发酵1、2、3、8 a的乌虾酱(学名糠虾酱)为研究对象,使用色差仪和描述性定量分析(quantitative descriptive analysis,QDA)法感官评价不同发酵阶段的虾酱品质;采用Illumina MiSeq高通量测序分析不同发酵阶段虾酱中细菌群落结构及多样性变化规律;运用高效液相色谱(high performance liquid chromatography,HPLC)检测虾酱长期发酵过程中6种常见生物胺含量变化;经Pearson相关性分析探究细菌群落结构对生物胺形成的影响,识别形成生物胺的特征菌群,为精准防控长期发酵虾酱中生物胺积累提供标靶菌群信息。
1 材料与方法
1.1 材料与试剂
乌虾酱 辽宁省锦州笔架山食品厂。将捕捞的乌虾修剪洗净后放入发酵罐(陶罐高0.57 m、口径0.50 m、质量75 kg),加入虾体质量16%的食盐搅拌均匀后,用草盖封口置于室外自然发酵,温度控制在10~30 ℃。选取发酵1、2、3、8 a的乌虾酱各(200±10)g,并随机从同一年份的不同发酵缸中收集3 份平行样品,使用无菌采样袋(230 mm×140 mm)运送至渤海大学水产品贮藏加工与安全控制实验室,于4 ℃冷藏备用。
DNeasy PowerSoil Kit(细菌基因组DNA提取试剂盒)德国Qiagen公司;聚合酶链式反应(polymerase chain reaction,PCR)扩增试剂盒 生工生物工程(上海)股份有限公司;AMPure XP 核酸纯化试剂盒 美国贝克曼库尔特有限公司;腐胺(≥98.4%)、尸胺(≥98.1%)、组胺(≥98.3%)、酪胺(≥99.5%)、精胺(≥97.2%)、亚精胺(≥99.2%)、丹磺酰氯、脯氨酸、乙酸铵(均为色谱纯) 美国Sigma-Aldrich公司;乙腈、甲醇、丙酮、碳酸氢钠、氢氧化钠(均为分析纯) 辽宁省沈阳化学试剂厂。
1.2 仪器与设备
Illumina MiSeq高通量测序仪 美国Illumina公司;QIAcube HT高通量核酸纯化仪 德国Qiagen公司;Biofuge Stratos冷冻高速离心机、Nanodrop 2000紫外-可见分光光度计 美国Thermo Fisher Scientific公司;FireReader XS凝胶成像系统 英国UVItec公司;Qubit 2.0荧光计 美国Life Technologies公司;GI54DS立式高压蒸汽灭菌锅 福建省致微(厦门)仪器有限公司;StepOne Plus实时荧光定量PCR仪 新加坡ABI公司;LC-2030高效液相色谱一体机 日本岛津公司;Millipore超滤系统 美国Millipore公司;CR-10 Plus色差仪 日本Konica Minolta公司。
1.3 方法
1.3.1 虾酱色泽测定
对不同发酵阶段的虾酱外观拍照记录,采用色差仪对虾酱色泽差异进行分析。
1.3.2 虾酱感官评价测定
采用QDA对不同发酵期的虾酱进行感官评价。评定小组为20 名经过感官培训后的成员,根据样品的风味特征(虾味、鲜味、腥味、咸味、酸味、氨气味、发酵味)进行评分,使用10 分制评分感官强度(0 分表示没有,10 分表示极度强烈)。
1.3.3 细菌群落结构分析
1.3.3.1 虾酱中细菌群落总DNA提取
采用细菌基因组DNA提取试剂盒提取不同发酵阶段虾酱中细菌总DNA,随后取50 ng DNA使用1%琼脂糖凝胶电泳进行质量检测,分析DNA的完整性和相对浓度。
1.3.3.2 DNA的扩增与纯化
采用细菌通用引物341F/806R(341F:CCTAYGGGRBGCASCAG;806R:GGACTACNNGGGTATCTAAT)对16S rDNA V3-V4可变区片段进行PCR扩增,第1轮扩增程序为:95 ℃预变性3 min,98 ℃变性10 s,55 ℃退火30 s,69 ℃延伸30 s,共循环30 次,最后68 ℃再延伸3 min。第1轮PCR产物用AMPure XP核酸纯化试剂盒纯化,再用Qubit 2.0定量。第2轮扩增程序为:95 ℃预变性3 min,98 ℃变性10 s,58 ℃退火30 s,72 ℃延伸30 s,共循环15 次,最后72 ℃再延伸5 min。产物进行纯化后经PCR仪定量,用Illumina MiSeq高通量测序仪测序。
1.3.3.3 测序数据分析
原始测序数据经拼接和过滤得到有效数据,然后利用FLASH v1.2.11软件将双末端测序得到的成对reads进行组装,得到高变区的tags。利用USEARCH v7.0.1090软件将组装好的tags在97%相似度下聚类,得到物种分类的操作分类单元(operational taxonomic unit,OTU)序列,并绘制稀释曲线和物种多样性曲线。利用Mothur v1.31.2软件进行主成分分析(principal component analysis,PCA)、α多样性指数组间差异分析(t检验),包括ACE指数和Chao1丰度指数、Simpson指数、Shannon多样性指数、PD_whole_tree和Observed_species指数。利用QIIME v1.80软件中Unweighted UniFrac计算不同样品间的β多样性值,测定样品间群落组成的差异性。
1.3.4 长期发酵虾酱中生物胺测定
参照陈功等[14]的方法提取不同发酵阶段虾酱样品中生物胺,采用内标法和HPLC测定生物胺含量,并根据仪器条件稍作改进。HPLC条件:Agilent C18色谱柱(4.6 mm×250 mm,5 μm);柱温37 ℃;进样量5 μL;流速1 mL/min;流动相:A为0.01 mol/L乙酸铵缓冲液-乙腈(90∶10,V/V);B为0.01 mol/L乙酸铵缓冲液;梯度洗脱程序:0~18 min,40%~0% A、60%~100% B;19~24 min,0% A、100% B;25~35 min,0%~40% A、100%~60% B。各色谱峰在254 nm波长处检出。
1.4 数据处理
采用IBM SPSS statistic 22.0软件进行数据差异分析,Excel 2010软件进行线性回归分析,Origin 2018软件进行绘图。细菌多样性数据通过SPSS 22.0软件进行单因素方差分析(P<0.05,差异显著)。利用Pearson相关性检验分析标准化后变量间的相关性。
2 结果与分析
2.1 虾酱感官特征分析
如图1A所示,发酵1 a的虾酱呈褐色,表观呈不规则颗粒状;随发酵时间延长,虾酱颜色逐渐变深;发酵8 a的虾酱呈深褐色、光滑、外观无颗粒状。由图1B可知,发酵1~8 a期间亮度(L*)值逐渐降低,发酵8 a样本的L*值低至26.77;虾酱发酵1~3 a,红度(a*)值逐渐减小,但发酵8 a虾酱的a*值增加;黄度(b*)值在发酵过程中呈先上升后下降趋势,在发酵8 a达到最低。
由图2可知,发酵1~8 a的虾酱风味存在明显差异。随发酵时间延长,虾酱的虾味、腥味和氨气味均降低,在发酵8 a达到最低;鲜味在发酵2 a达到最大值,随后逐渐降低;此外,虾酱中酸味较低、咸味较高。

图1 不同发酵年份的虾酱外观(A)和色泽分析(B)Fig. 1 Appearance (A) and color parameters (B) of shrimp paste samples during fermentation
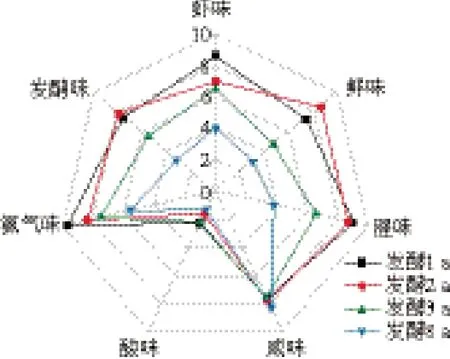
图2 不同发酵年份的虾酱滋味QDA雷达图Fig. 2 QDA radar chart of the odor and taste of shrimp paste fermented for different periods
2.2 虾酱长期发酵过程中细菌多样性变化
2.2.1 Illumina MiSeq测序数据统计及质控分析
利用16S rRNA测序技术研究1、2、3 a和8 a发酵虾酱细菌群落结构和多样性变化。由图3a可知,随测定序列数增加,所有样品可观测OTU数量不断上升;当测序深度大于25 000时,各样品中观测的OTU数量逐渐趋于平缓,样本菌落覆盖率达98.9%。由图3b可知,不同发酵阶段虾酱样品中细菌群落的相对丰度随可观测OTU数量的增加逐渐趋于稳定;当样品的可观测OTU数量大于200时,所有样品的多样性曲线趋于平滑,表明虾酱样品中细菌群落分布趋近均匀,群落结构无偏向性。从不同发酵阶段虾酱中测得总tag数分别为48 515±9 423、42 799±7 651、53 984±6 894和56 504±2 789,各筛选出(41 206±6 328)、(39 875±8 432)、(50 163±8 999)条和(55 132±5 172)条优化序列;各组有效序列数均远大于测序深度阈值(25 000),4 组虾酱样品细菌群落覆盖率达98.1%~99.5%,且测得的序列分别聚类于(1 502±87)、(2 008±113)、(1 611±202)、(515±19)个OTU(图4)。
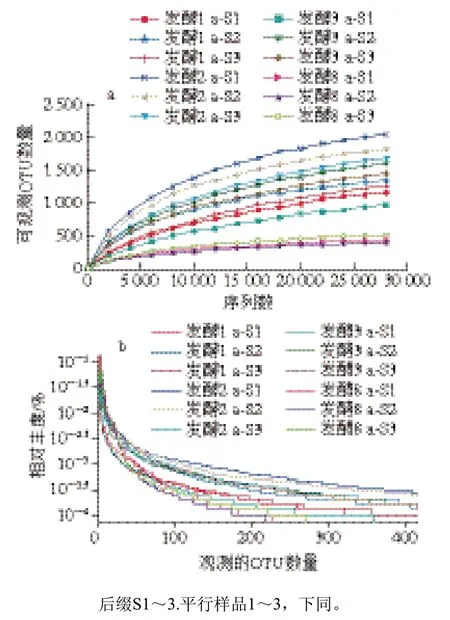
图3 不同发酵阶段虾酱的细菌稀释曲线(a)和多样性曲线(b)Fig. 3 Rarefaction (a) and diversity (b) curves of bacterial communities in shrimp paste at different fermentation stages

图4 虾酱长期发酵过程中细菌16S rRNA测序数据统计Fig. 4 Statistical analysis of 16S rRNA sequencing data of bacteria in shrimp paste at different fermentation stages
2.2.2α多样性分析
虾酱样品经α多样性指数分析,得到组内菌群多样性指数和丰富度指数,数据经Wilcox秩和检验和Tukey检验分析组间物种的多样性差异。ACE指数和Chao1指数反映样品中群落的丰富度。由图5可知,不同发酵阶段虾酱细菌群落的ACE指数和Chao1指数变化规律一致,随发酵时间延长,虾酱中细菌多样性逐渐增加,在发酵2 a时细菌多样性最高,表明细菌群落中物种数量较多,在发酵3、8 a时菌群多样性显著降低。Shannon指数和Simpson指数综合体现物种丰富度。发酵1 a虾酱的Shannon指数和Simpson指数与其他组相比较小,表明菌群丰富度较低,发酵2 a菌群丰富度值达到最大值,随后发酵时间延长,群落丰富度逐渐降低。此外,虾酱样品中Observed_species指数和进化距离的多样性评估指数(PD_whole_tree)也与多样性显著相关,均在发酵2 a达到最大值,在发酵8 a时最低,表明长期发酵使虾酱细菌群落单一化。
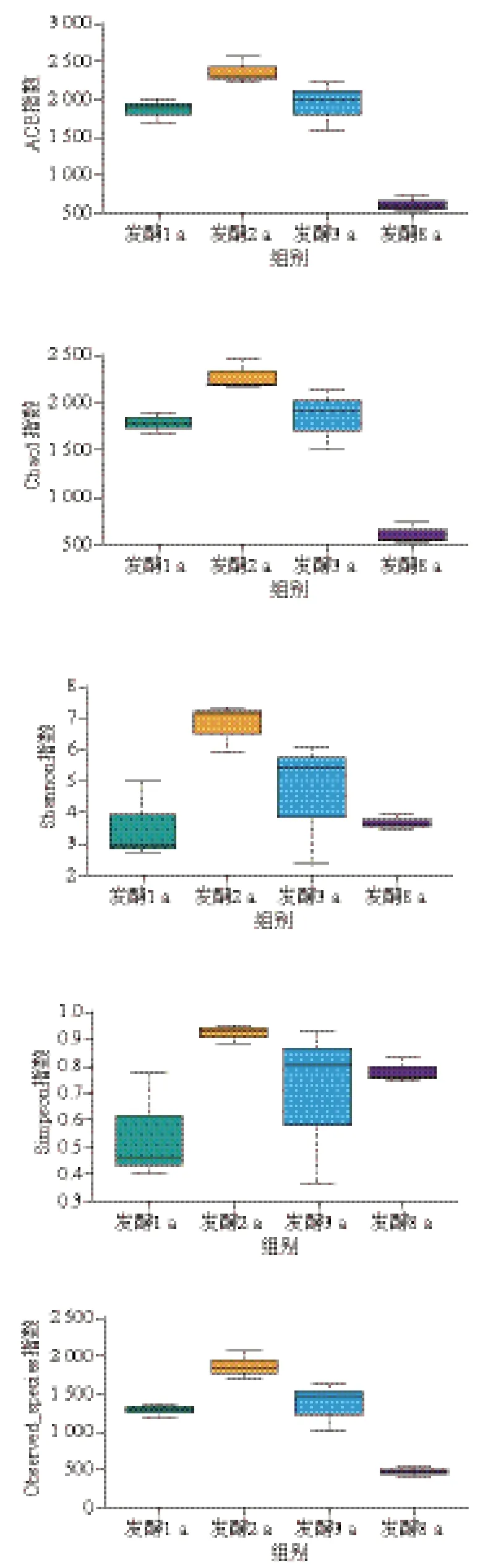

图5 不同发酵阶段虾酱细菌α多样性分析Fig. 5 α Diversity analysis of bacterial communities in shrimp paste at different fermentation stages
2.2.3β多样性分析
基于Unweighted UniFrac距离衡量法进行β多样性分析,比较不同发酵阶段虾酱细菌多样性差异。由图6a可知,4 个发酵阶段虾酱样品β多样性指数显示,发酵1~3 a虾酱组样品的相异指数为0.415~0.581,表明3 组样品间物种多样性较小;8 a发酵组与1~3 a发酵组的相异指数范围为0.642~0.774,说明发酵8 a虾酱的细菌群落与中短期发酵具有显著差异。由图6b可知,基于对细菌属水平聚类分析,发现1~3 a发酵组细菌种类均聚在同一分支,8 a发酵组样品单独聚为一类,此结果与β多样性指数结果一致。表明虾酱前期发酵(1~3 a)的细菌群落结构与后期发酵(8 a)的群落结构差异较大,而在1~3 a内的发酵阶段,各组群落结构差异较小。这说明在前期发酵虾酱中细菌群落结构变化较大,后期发酵时间越长,细菌群落结构趋于稳定。
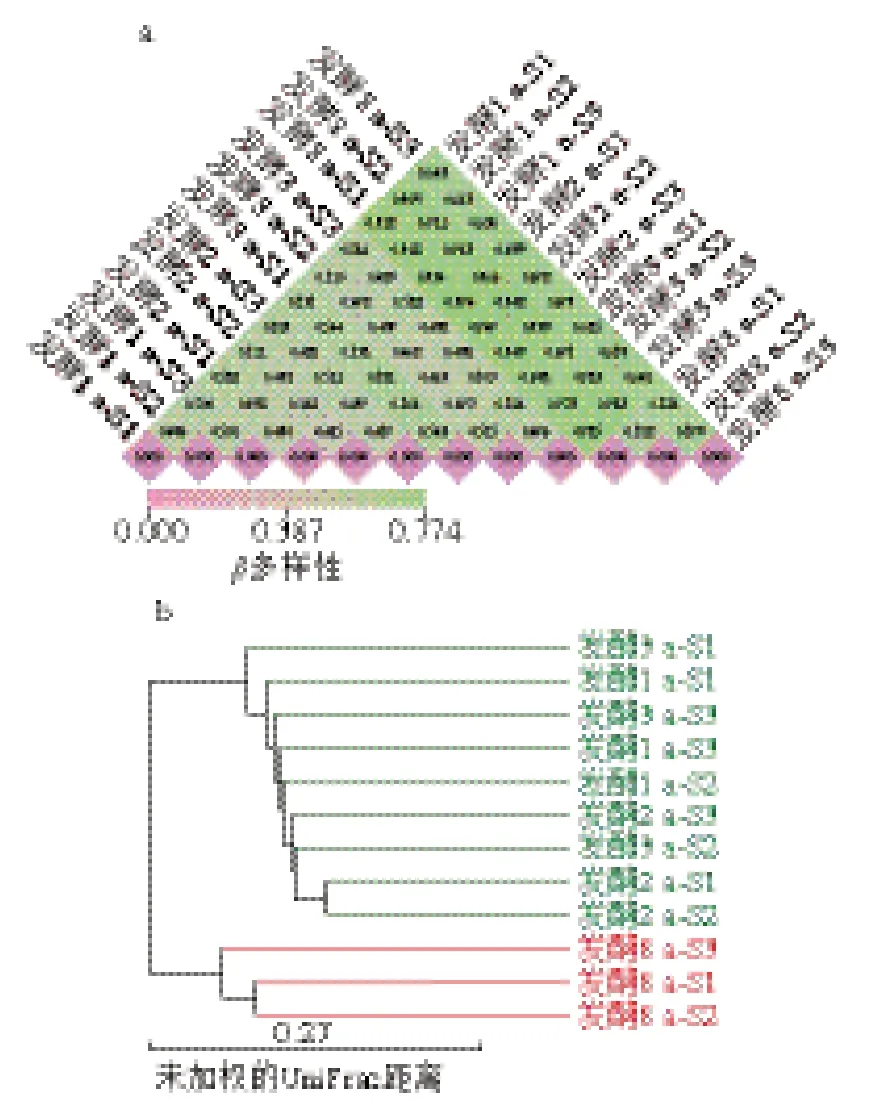
图6 不同发酵阶段虾酱细菌β多样性指数(a)和聚类分析(b)图Fig. 6 β Diversity index (a) and cluster analysis (b) of bacterial communities in shrimp paste at different fermentation stages
2.2.4 Venn图和PCA
对不同虾酱组中测得在相似水平为97%以上所有OTU数目的相似性和重叠性进行归类,得到组间差异Venn图,如图7a所示,4 个发酵阶段共有的细菌种类为161 个,分别占1、2、3、8 a发酵组细菌总数的41.6%、33.5%、38.6%和69.6%,其中1、2、3 a发酵组共同含有282 个OTU,表明这3 组的细菌群落相似度较高。进一步对不同发酵阶段独有OTU数量分析可知:2 a发酵组(58 个)>3 a发酵组(43 个)>1 a发酵组(26 个)>8 a发酵组(24 个),说明发酵2 a虾酱独有的细菌种类最多,随着发酵时间延长,虾酱中细菌群落结构逐渐相似。采用PCA识别虾酱样品的组内和组间差异性,根据各样品在PCA得分图中的分布区域和距离,判定样品间相似度。由图7b可知,PC1和PC2对区分样本差异的贡献率分别为53.19%和26.85%,总贡献率>80%,表明这两个因素能够较好地反映样品中生物多样性特征。4 个发酵阶段虾酱样品主要分布在第1、3、4象限,其中2 a发酵组的组内平行样本间距离较远,表明平行样本的细菌群落波动性大,细菌群落发生剧烈更替;而1、3、8 a发酵组的组内平行样本间距离较近,说明样本平行性较好,细菌群落构成相对稳定;此外,1 a和3 a发酵组的组间样本距离较近且有重叠,说明二者细菌群落构成相似。2 a和8 a发酵组的组间样本距离较远,并与1 a和3 a发酵组无重叠,表明2 a和8 a发酵虾酱菌群丰度差异显著。
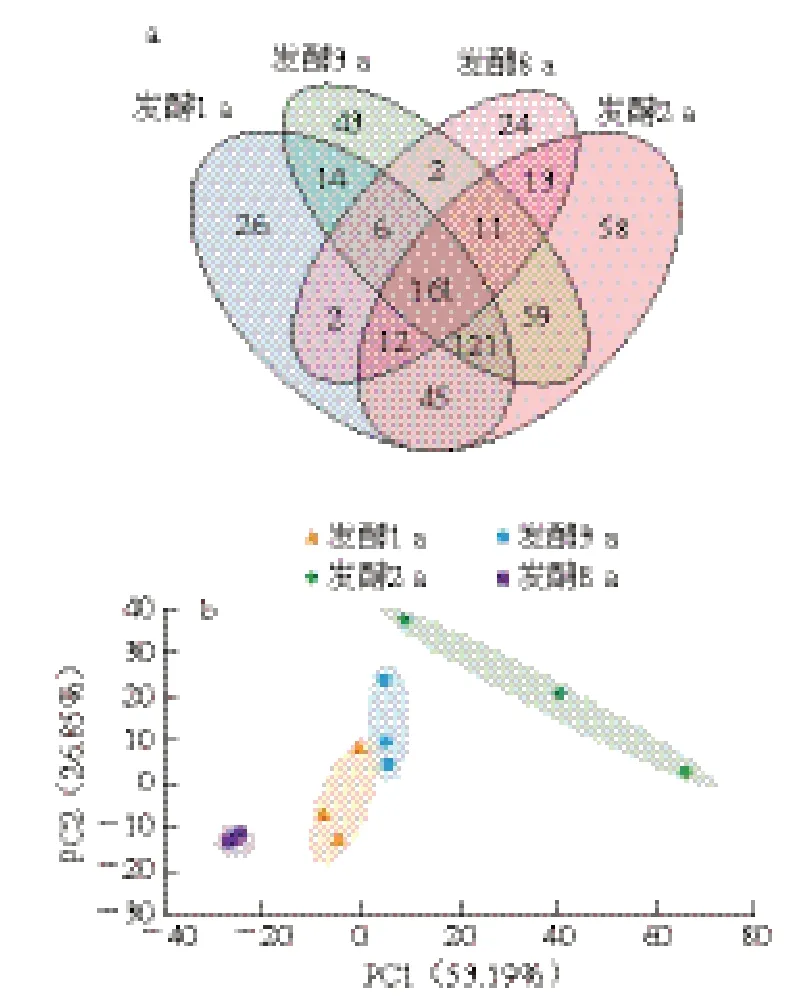
图7 不同发酵阶段虾酱细菌群落Venn图(a)和PCA得分图(b)Fig. 7 Venn diagram (a) and PCA score plot (b) of bacterial communities in shrimp paste at different fermentation stages
2.3 虾酱长期发酵过程中细菌群落的演替分析
2.3.1 门和属水平细菌群落结构变化
由图8a可知,在细菌群落门水平上,1 a发酵组的优势菌群分为3 类,包括:厚壁菌门(Firmicutes,82.35%)、变形菌门(Proteobacteria,11.52%)和放线菌门(Actinobacteria,3.55%),占细菌群落总平均丰度97.37%;发酵2 a时,样品中优势菌群显著增加到10 类,新增菌群为拟杆菌门(Bacteroidetes,3.08%)、酸杆菌门(Acidobacteria,1.7%)、髌骨细菌门(Patescibacteria,1.22%)、绿弯曲门(Chloroflexi,0.66%)、芽单胞菌门(Gemmatimonadetes,0.44%)、疣微菌门(Verrucomicrobia,0.4%)和装甲菌门(Armatimonadete,0.3%),而厚壁菌门、变形菌门和放线菌门分别下降至41.11%、9.92%和2.88%;发酵3 a的虾酱样品中无明显新增菌门,绝大多数菌门的相对丰度降低,仅厚壁菌门增加至59.1%;发酵8 a后,核心优势菌门为厚壁菌门和变形菌门,分别占相对丰度的46.91%和52.42%,其他菌门的相对丰度均显著降低。
由图8b可知,在细菌属水平上,共鉴定出相对丰度高于0.1%的33 个菌属,1 a发酵组中的优势菌属为:四联球菌属(Tetragenococcus,71.7%)、嗜冷杆菌属(Psychrobacter,2.73%)、芽孢杆菌属(Bacillus,1.77%)、劳尔氏菌属(Ralstonia,1.96%)、假单胞菌属(3.62%)、庞蒂亚克杆菌属(Pontibacter,2.61%)、链球菌属(Streptococcus,2.58%)、肠球菌属(2.55%)、嗜冷杆菌属(Psychrobacter,1.73%)、乳杆菌属(1.32%)、玫瑰变色菌属(Roseovarius,1.17%)和副球菌属(Paracoccus,0.25%)等;当虾酱发酵2 a时,四联球菌属相对丰度降低为54.5%,而肠球菌属、嗜冷杆菌属、链球菌属、假单胞菌属相对丰度明显上升,同时出现新优势菌属包括棒状杆菌属(Corynebacterium_1,1.66%)、拟杆菌属(Bacteroides,0.78%)和不动杆菌属(Acinetobacter,0.92%);发酵3 a时,四联球菌属相对丰度为50.56%,优势菌属以假单胞菌属(4.12%)、弧菌属(1.48%)和玫瑰变色菌属(1.26%)为主;发酵8 a虾酱样品中细菌群落构成与1~3 a样品存在明显差异,其中四联球菌属相对丰度显著降低至31.36%,假单胞菌属和弧菌属的相对丰度显著增加至23.69%和8.49%。因此,不同发酵时期在属水平下虾酱的菌群多样性具有显著差异,四联球菌属和链球菌属为虾酱整个发酵过程的优势菌属。
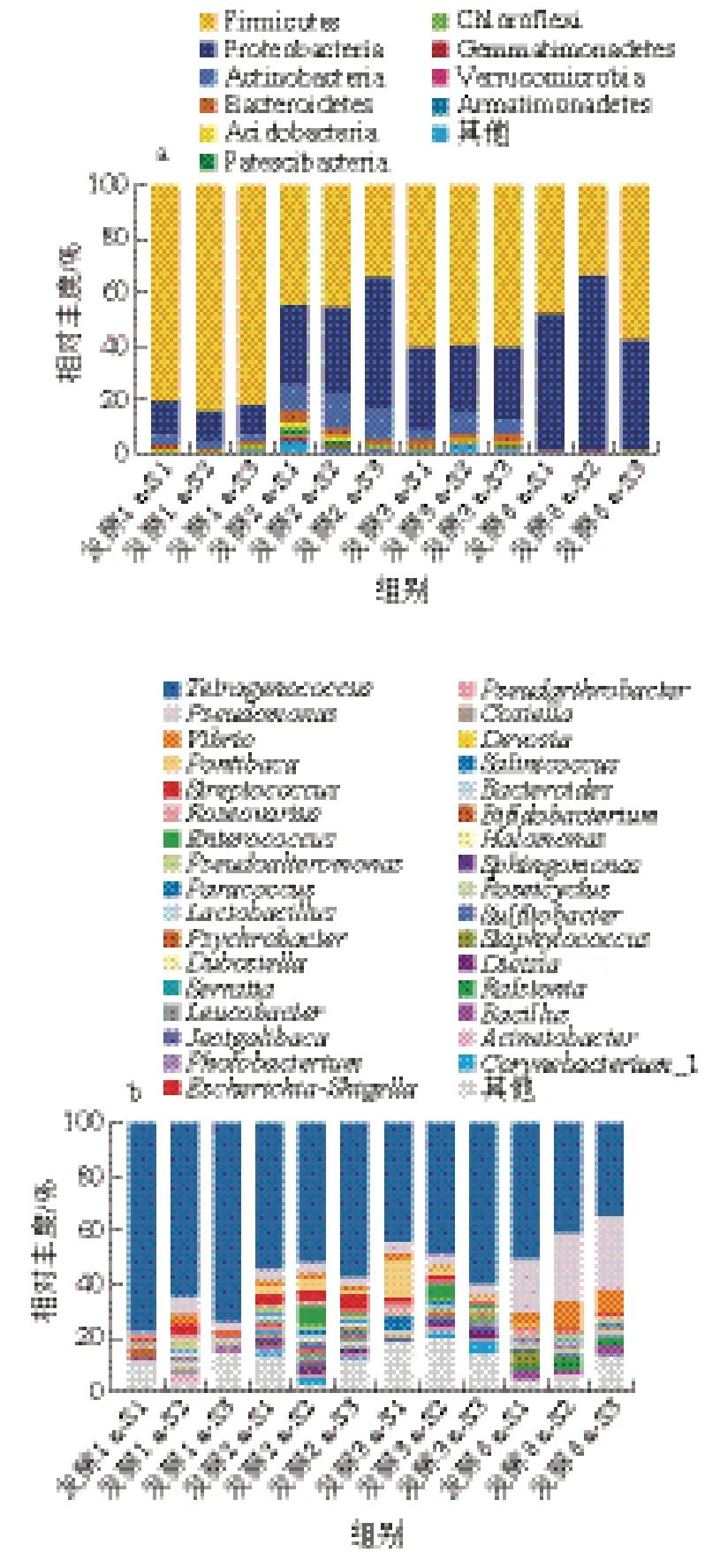
图8 不同发酵阶段虾酱菌群在门(a)和属(b)水平的构成图Fig. 8 Phylum-level (a) and genus-level (b) composition of bacteria communities in shrimp paste at different fermentation stages
2.3.2种水平细菌群落多样性分析
根据高通量测序结果在种分类水平上筛选相对丰度大于0.1%的可识别细菌群落组成,共得到14种,如表1所示,穆氏葡球菌(T. muriaticus)为发酵1 a虾酱中的优势菌种,相对丰度达65.81%,随发酵期延长相对丰度整体呈降低趋势,在发酵8 a相对丰度高度显著降低且仅为5.48%(P<0.001)。而产蓝假单胞菌(P. syncyanea)和嗜盐四联球菌(T. halophilus)、瓦拉尼链球菌(S. varani)和血光杆菌(P. sanguinicancri)在8 a发酵组中相对丰度均显著升高(P<0.05),分别为(31.47±4.11)%、(28.49±2.34)%、(0.48±0.09)%和(0.92±0.25)%。此外,大部分丰度较高的细菌在发酵2 a的虾酱中达到峰值,如粪肠球菌(E. faecalis)、嗜酸乳杆菌(L. acidophilus)、暹罗盐球菌(S. siamensis)、J. dankookensis、立克次氏体(C. cheraxi)、丝状白杆菌(L. holotrichiae),随后随发酵周期延长,相对丰度降低。
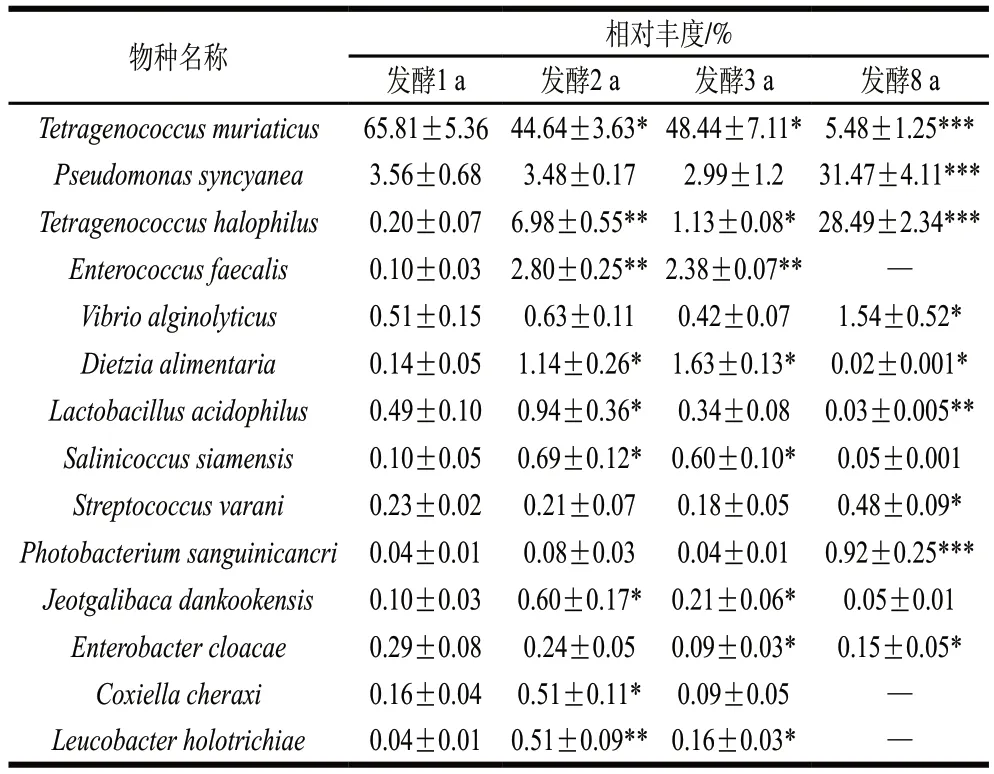
表1 不同发酵阶段虾酱种水平上相对丰度大于0.1%可识别的细菌组成Table 1 Identifiable bacterial composition with relative abundance greater than 0.1% at specie level in shrimp paste at different fermentation stages
2.4 虾酱长期发酵过程中生物胺含量变化
2.4.1 不同发酵阶段虾酱中生物胺含量分析
6种生物胺混标经丹磺酰氯衍生后均在进样35 min内出峰,且色谱峰峰形对称完整、无重叠,表明洗脱程序可将生物胺较好分离。对不同发酵年份的虾酱生物胺含量分析可得(表2),1 a发酵组仅有腐胺和尸胺;2 a和3 a发酵组检出的生物胺种类一致,均含有腐胺、尸胺和组胺;8 a发酵组生物胺种类多达6种,包括:腐胺、尸胺、组胺、酪胺、亚精胺和精胺,说明随着发酵时间延长,虾酱中生物胺种类逐渐增加。腐胺在1 a发酵组中含量最低,为(1.65±0.67)mg/kg,随发酵时间延长,腐胺含量在2~3 a发酵中显著升高(P<0.05),在8 a发酵组中含量高度显著提高(P<0.001),达到最高值(57.08±8.31)mg/kg;尸胺在1 a发酵组中检出含量最高(53.13±5.83)mg/kg,在2 a和3 a发酵组中均降低,分别为(41.11±10.75)mg/kg和(43.79±8.99)mg/kg,在8 a发酵组中降低至(35.67±6.84)mg/kg;组胺在1 a发酵组中未检出,在2、3、8 a发酵组中含量逐渐增加,在8 a发酵组中达到最大值(14.41±2.28)mg/kg;酪胺、亚精胺和精胺仅在8 a发酵组中检出,含量分别为(11.94±1.21)、(0.88±1.37)、(0.98±0.11)mg/kg。表明除了尸胺外,其他5种生物胺含量都随着发酵期延长而不断积累,发酵期越长,虾酱中总生物胺含量越高。
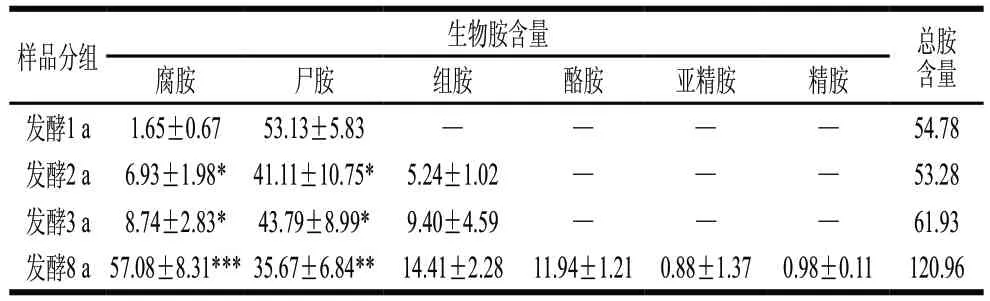
表2 不同发酵阶段虾酱的生物胺含量变化Table 2 Changes in contents of biogenic amines in shrimp paste at different fermentation stages mg/kg
2.4.2 细菌群落结构对虾酱中生物胺形成的影响
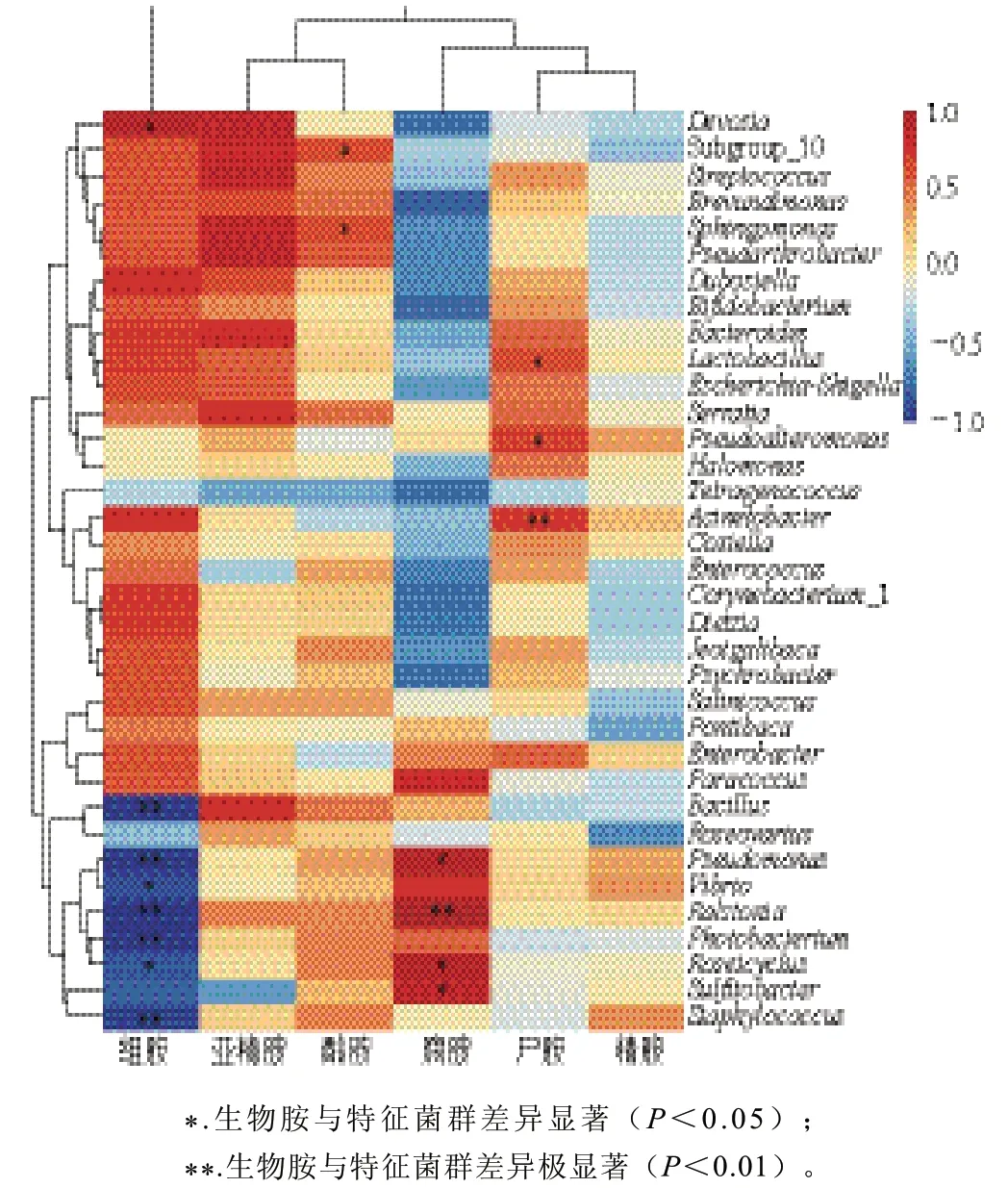
图9 虾酱中优势菌群与生物胺之间的Pearson相关性分析Fig. 9 Pearson’s correlation analysis between dominant bacterial communities and biogenic amines in shrimp paste
基于Pearson相关性检验,对虾酱中生物胺与细菌群落关联度分析。由图9可知,虾酱中组胺、酪胺、腐胺和尸胺与发酵1~8 a阶段的优势菌群密切相关,而亚精胺和精胺与细菌群落无明显关联。组胺含量变化与德沃斯菌属(Devosia)呈显著正相关(P<0.05),与芽孢杆菌属、劳尔氏菌属、发光杆菌属(Photobacterium)、葡萄球菌属呈极显著负相关(P<0.01);腐胺含量变化与劳尔氏菌属呈极显著正相关(P<0.01),与假单胞菌属、慢生盐场菌属(Roseicyclus)、亚硫酸杆菌属(Sulfitobacter)呈显著正相关(P<0.05)。尸胺主要与不动杆菌属呈极显著正相关(P<0.01),与乳酸菌属和假交替单胞菌属呈显著正相关(P<0.05)。酪胺含量变化主要与鞘氨醇单胞菌属(Sphingomonas)呈显著正相关(P<0.05)。芽孢杆菌属、假单胞菌属和葡萄球菌属是发酵1 a的虾酱优势菌群,与组胺呈极显著负相关(P<0.01),表明其可抑制组胺形成,因此在1 a发酵虾酱中未检出组胺。与酪胺、腐胺和尸胺呈显著正相关的优势菌群主要在2~8 a发酵的虾酱中丰度较高,在这3种生物胺的形成上发挥重要作用。
3 讨论与结论
经高通量测序发现,虾酱在长期发酵过程中,具有较厚细胞壁、可产生内生孢子的厚壁菌门逐渐演替为优势菌门,主要包括四联球菌属、葡萄球菌属、芽孢杆菌属、链球菌属和乳杆菌属等。也有报道指出,乳杆菌属具有产胺酶基因,与啤酒发酵过程中的酪胺、组胺、腐胺和尸胺的产生显著相关[15]。葡萄球菌在发酵香肠中被检测是酪胺的主要产生菌[16]。本研究发现,虾酱中细菌群落多样性在发酵2 a达到峰值,随后逐渐降低,适应性较弱的细菌群落逐渐消失,抗逆性强、嗜盐性强和代谢能力较弱的革兰氏阳性菌占优势,如嗜酸乳杆菌和暹罗盐球菌等。嗜酸乳杆菌与虾酱尸胺增加显著相关,在其他虾酱发酵过程中也有报道[10],这可能与嗜酸乳杆菌具有较强的赖氨酸脱羧酶活性有关。前期发酵1~3 a的虾酱,经PCA表明,细菌群落组成具有一定相似性,而发酵8 a的菌落结构具有特殊性,与1~3 a发酵虾酱的共有菌群显著降低;β多样性显示1~3 a发酵虾酱聚类同一分支,而发酵8 a单独为一分支,表明在前期发酵过程中,细菌群落演替较为频繁,菌群无显著差异性,长期发酵后,群落结构趋于稳定。
目前普遍认为,虾酱发酵时长有利于风味物质形成,主要是由于部分菌属分解蛋白质形成特殊风味,Prihantoab等[17]发现双歧杆菌属、片球菌属和魏斯氏菌属等乳酸菌,能产生丙氨酸、缬氨酸、天冬氨酸、苏氨酸等多种氨基酸,有利于提供发酵食品的滋味,但部分乳酸菌如干酪乳杆菌和嗜酸乳杆菌具有组氨酸和酪氨酸脱羧酶,对组胺和酪胺的形成具有重要影响[18]。此外,虾酱发酵8 a后假单胞菌属和弧菌属的丰度增加。假单胞菌属是水产食品常见的腐败菌和致病菌,相关性分析发现其与尸胺增加显著相关。另外,假单胞菌属和弧菌属会分解蛋白质产生硫化氢气味和腥臭味等不良气味,影响虾酱的风味和安全性[19-20]。因此,虾酱长期发酵过程中应预防环境中腐败菌和致病菌污染。
虾酱由小虾在腌制过程中增加盐分可抑制细菌繁殖引起的腐败,但长期存在的脱羧酶阳性菌群对氨基酸的转化和水解,是产生尸胺、腐胺、组胺等有害生物胺的主要途径[21]。在发酵过程中,主要存在的生物胺为尸胺和腐胺,且尸胺仅发酵1 a含量已超过我国水产品中生物胺限量标准(50 mg/kg)[22]。目前,生物胺超标一直是发酵水产品的棘手问题,Li Wenya等[23]对我国渤海湾的食品工厂发酵虾酱调查发现总胺含量为18.20~469.02 mg/kg,尸胺和组胺严重超标。Ma Xinxiu等[24]调查12 个月发酵鱼的产胺量,发现组胺含量高达55 mg/kg,且生物胺总量增加。虾酱长期发酵使腐胺和尸胺生成显著增加,且发酵8 a虾酱中含有腐胺、尸胺、组胺、酪胺、亚精胺和精胺,可能是由于发酵期产酸微生物活跃,通过糖酵解将原料中的碳水化合物分解为磷酸、乳酸等酸性代谢物质使虾酱pH值降低,导致大量脱羧酶阳性菌群为维持pH值的相对平衡,将部分氨基酸脱羧转化为生物胺[25-26]。Pearson相关性分析发现,假单胞菌属、慢生盐场菌属和亚硫酸杆菌属与腐胺呈正相关;不动杆菌属、乳酸菌属和假单胞菌属与尸胺呈正相关。研究发现,产胺能力较强的多为抗逆性较强的菌属,具有较好适应环境的能力。Jančová等[27]调查奶业厂废水中的葡萄球菌属、乳球菌属、肠球菌属、科氏菌属、不动杆菌属、假单胞菌属和肠杆菌属等的产胺能力,发现所有菌株均表现脱羧酶基因阳性,且能产生4~8种生物胺。
针对发酵水产品中生物胺含量超标的问题,主要通过添加甘氨酸、茶多酚和姜辣素等天然活性物质,或添加微生物发酵剂降低虾酱体系中微生物的生长繁殖和消减胺类物质[26,28],或从虾酱中筛选分离生物胺降解菌如维吉芽孢杆菌和盐反硝化枝芽孢杆菌等,再接种于发酵虾酱,降低生物胺的产生[29-30]。在本研究中,穆氏葡球菌为虾酱发酵期间细菌占比最大的优势菌,在泰国发酵鱼露[31]和日本发酵水产品[32]中也均有检出,属嗜盐发酵乳酸菌。Sang Xue等[33]针对虾酱源穆氏葡球菌进行安全性评估,发现与组胺产生无显著相关性,且对抗生素有弱抗性、无溶血活性和不产生生物被膜,产生脂肪酶和蛋白酶,可作为虾酱发酵期间控制组胺的潜在优势菌株。另外,嗜盐四联球菌在发酵8 a丰度显著增加,研究发现其具有较强的抗酸胁迫性和热适应性,为水产品发酵制品中的优势菌株[34]。Kim等[35]从发酵鱼中分离出此菌,经基因组测序未发现生物胺产生相关基因,接种发酵鱼可显著抑制尸胺的生成。因此,开展虾酱中优势菌属的降胺能力研究,有助于筛选潜在的功能性发酵菌株,对于功能菌种干预生物胺形成的机理,未来研究中将结合生物胺相关理化指标和菌群代谢特征进行深入探究。
