棘茎楤木和树参对林窗、林缘和林下生境的光合生理响应
2022-12-28李孝初王盼康华靖郭书亚周钰鸿潘德月陈子林肖琛闻
李孝初,王盼,康华靖,郭书亚,周钰鸿,潘德月,陈子林,肖琛闻
(1.宁波鄞州中铭农业科技有限公司,浙江 宁波 315040; 2.磐安县中药创新发展研究院,浙江 磐安 322300;3.温州市农业科学研究院,浙江 温州 325006; 4.浙江师范大学 植物学实验室,浙江 金华 321004;5.浙江大盘山国家级自然保护区管理局,浙江 磐安 322300; 6.浙江省农业科学院,浙江 杭州 310021)
棘茎楤木(Araliaechinocaulis)属五加科楤木属,在我国甘肃、陕西、福建等地区均有分布[1],生于森林、灌丛或林缘路边,垂直分布从海滨至海拔3 000 m左右。树参(Dendropanaxdentiger)为五加科树参属植物,在我国南方地区分布广泛,可生长在海拔10~2 000 m的常绿阔叶林或灌丛中[2]。前者具有镇痛消炎、祛湿活血之效[3],同时也具有重要的营养价值,茎皮中含墩果酸、刺囊酸及马栗树皮素二甲酯等物质。后者的根[4]、茎[5]、叶[6]均有重要的药用价值,可治偏头痛、风湿痹痛等症[7],嫩叶芽可以腌制、爆炒、凉拌等多种方式食用[6],味道鲜美,营养价值高。国内有关棘茎楤木和树参的研究集中于幼苗繁育[8]、生长发育规律[9-10]、栽培技术[11]等方面。
发展林下经济正成为充分利用林地资源、促进林业发展的一个新方向。利用林下特有生境种植药用和食用植物,可提高林地资源利用效率,缓解耕地不足,增加森林经济效益。光照是影响林下植物生长和产量的主要因子[12]。若光照过弱,则光合产物积累较少,影响植株生长;而光照过强,则会出现光抑制而降低光合作用,进而抑制植株的生长。探明棘茎楤木和树参的适宜光照范围及生理特性对其规模化推广种植,批量化育苗等具有重要意义。为此,本试验设置了林窗、林缘和林下3种生境,拟从光合气体交换及叶绿素荧光参数两个角度,探讨棘茎楤木和树参的适宜光强及光合生理特征,以期为其林下种植推广提供参考。
1 材料与方法
1.1 试验地点与处理设计
2020年2月在浙江省磐安县祠下村杉木林林窗(A)、林缘(B)和林下(C)生境下分别种植棘茎楤木和树参,日常实行统一管理。于2020年8月下旬进行试验测量。每个生境选择5~7株植物,每株植物选取1片顶部完全展开的叶子待测。
1.2 测定指标
利用便携式光合仪(LI-6400-40, LI-COR INC., USA)于晴天9: 00—16: 30进行数据测量。数据测量前进行开机预热及各项检查。在自然光下诱导1.5~2.0 h后,采用开放式气路,流速为400 μmol·s-1。根据预测量仪器中叶室温度(Tblock)的动态变化,设定叶室温度约为29.47 ℃(大气温度约为30.48 ℃左右),空气相对湿度控制在45%~70%,光合有效辐射强度(PAR)梯度设置为:1 400、1 200、1 000、800、600、400、200、150、100、50和0 μmol·m-2·s-1,应用CO2注入系统提供400 μmol·mol-1稳定的CO2。在此条件下同时测定植物叶片的气体交换参数和荧光参数。应用自动测量程序(FlrPn-Cacurve)进行数据测量。每次程序记录最小等待时间为2 min,最大等待时间为3 min,数据记录前仪器均自动进行参比室和样品室之间的匹配。同样,外界温度、湿度、光强等环境因子数据也采用便携式光合仪自带的传感器获得。
1.3 数据分析
根据光合计算软件4.1(井冈山大学生命科学学院/数理学院),应用光响应机理模型拟合电子传递速率对光强的响应曲线以及光合速率对光强的响应曲线。SPSS 19.0版本(SPSS, Chicago, IL)软件进行显著性分析。每个生境选取5个重复,采用Excel 2010作图。
2 结果与分析
2.1 3种生境下的光强和温度动态变化
由图1可看出,以8:00—14:00的数据为例,A生境下的光强和温度整体处于最高,其次是B生境,C生境下的光强和温度则整体处于最低,说明林窗、林缘和林下生境具有较大差异。
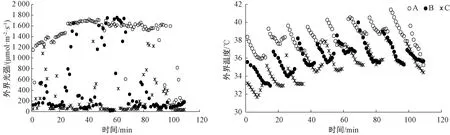
图1 3种生境(A、B和C)下外界光强及温度的动态变化
2.2 不同生境下棘茎楤木和树参叶片气体交换参数的响应
2.2.1 净光合速率对光强的响应
由图2可知,低光强(小于200 μmol·m-2·s-1)下,不同生境下的棘茎楤木和树参叶片的净光合速率均随PAR的增加几乎直线增加;在高光强(>800 μmol·m-2·s-1)下,其净光合速率则均有明显下降趋势。不同生境下的光合速率值整体上有所差异。

图2 不同生境下棘茎楤木和树参叶片的净光合速率对光的响应曲线
利用光响应机理模型拟合不同条件下光合作用对光的响应曲线,得到最大净光合速率(Pnmax)、净光合速率对应的饱和光强(Isat)、光补偿点(Ic)和暗呼吸速率(Rn)等光合参数见表1。由表1可知,拟合曲线所得到的拟合值与其所对应的实测值高度符合,其确定系数均大于0.90。从最大光合速率来看,C生境下的棘茎楤木和树参光合速率显著较高,分别为19.325和12.451 μmol·m-2·s-1。而同一树种在不同生境的饱和光强均较为接近,棘茎楤木和树参的光饱和点分别约为900和800 μmol·m-2·s-1。另外,不同生境下棘茎楤木和树参叶片的光补偿点均较低,其范围分别为19.590~28.392 μmol·m-2·s-1和13.200~23.829 μmol·m-2·s-1。

表1 不同生境下棘茎楤木和树参叶片光合作用对光的响应曲线拟合结果
2.2.2 蒸腾速率及气孔导度对光强的响应
由图3可知,棘茎楤木叶片蒸腾速率随着光强的增加而上升,高光强下趋于稳定;而树参叶片蒸腾速率随着光强的增加而呈上升趋势。不同生境下棘茎楤木叶片的蒸腾速率整体上均存在显著差异,以C生境下的值为最高;同样,树参叶片的蒸腾速率也以C生境下较高。由图4可知,在较低光强(<400 μmol·m-2·s-1)下,不同生境下棘茎楤木叶片气孔导度均随着光强的增加先直线上升,而后均呈缓慢下降趋势;而不同生境下树参叶片气孔导度的变化趋势有所差异。另外,同一植物不同生境下叶片的气孔导度整体上均存在显著差异,且均以C生境下的值为最高。
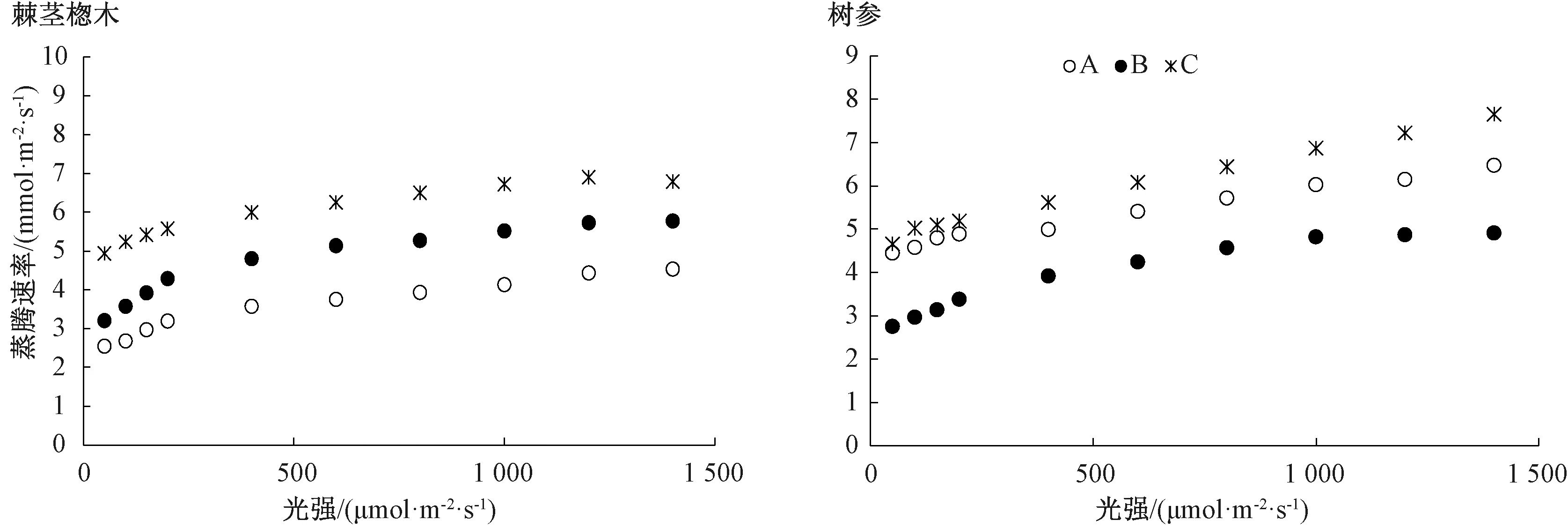
图3 不同生境下棘茎楤木和树参叶片的蒸腾速率对光的响应曲线

图4 不同生境下棘茎楤木和树参叶片的气孔导度对光的响应曲线
2.3 不同生境下棘茎楤木和树参叶片叶绿素荧光参数的响应
2.3.1 电子传递速率对光强的响应
由图5可知,PAR<400 μmol·m-2·s-1时,棘茎楤木和树参叶片的电子传递速率均随PAR的增加几乎呈直线增加;之后电子传递速率均在达到最大值后发生较为明显的PSⅡ动力学下调现象。PAR大于600 μmol·m-2·s-1时,不同生境下棘茎楤木叶片的电子传递速率存在显著差异;当光强高于500 μmol·m-2·s-1时,B和C生境下棘茎楤木叶片的电子传递率则有显著差异;C生境下棘茎楤木和树参的最大电子传递速率均为最高(P<0.05),分别为145.263和99.053 μmol·m-2·s-1。同样,两个树种叶片的最大电子传递速率所对应的饱和光强也均以C生境下的值为最高,分别为1 044.110和847.615 μmol·m-2·s-1(表2)。

图5 不同生境下棘茎楤木和树参叶片的电子传递速率对光的响应曲线

表2 不同生境下棘茎楤木和树参叶片光合电子传递速率对光的响应曲线结果
2.3.2 实际光化学量子产量(ΦPSⅡ)和天线转化效率(F′v/F′m)对光强的响应
由图6可知,不同生境下棘茎楤木和树参叶片ΦPSⅡ均随着光强的增加而下降。与B和C生境相比,A生境下棘茎楤木叶片ΦPSⅡ值整体较低,在相对较高光强(>400 μmol·m-2·s-1)下存在显著差异;而不同生境下树参叶片的ΦPSⅡ值均较为接近。不同生境下棘茎楤木和树参叶片F′v/F′m对光强的响应见图7。不同生境下棘茎楤木和树参叶片F′v/F′m均随着光强的增加而下降。A和C生境下,棘茎楤木叶片之间的F′v/F′m值在光强大于400 μmol·m-2·s-1时存在显著差异;不同生境下树参叶片之间的F′v/F′m值整体上无显著差异,只有在光强大于1 000 μmol·m-2·s-1时A生境下的F′v/F′m值显著较低。

图6 不同生境下棘茎楤木和树参叶片的实际光化学效率对光的响应曲线

图7 不同生境下棘茎楤木和树参叶片的F′v/F′m 对光的响应曲线
2.3.3 光化学猝灭系数(qP)和非光化学猝灭系数(qN)对光强的响应
由图8可知,不同生境下棘茎楤木和树参叶片qP均随着光强的增加而整体呈直线下降。不同生境下,棘茎楤木和树参(除高光强下树参A和B生境之间)叶片的qP整体上均无显著差异。由图9可知,不同生境下棘茎楤木和树参叶片qN均随着光强的增加而下降,A和C生境下,棘茎楤木叶片的qN值整体上无显著差异,而当光强较高时(≥400 μmol·m-2·s-1)树参叶片qN值整体上差异显著。

图8 不同生境下棘茎楤木和树参叶片的光化学猝灭系数对光的响应曲线

图9 不同生境下棘茎楤木和树参叶片的非光化学效率对光的响应曲线
3 小结与讨论
光是影响植物光合作用的重要因子。不同的光照条件会影响到植物的光合生理特性,以及叶绿素、可溶性糖、蛋白质含量和各种酶的活性,最终会影响到植物的生长,体现在苗高、地径、根重及生物量的变化[13-16]。本试验的环境因子监测表明,林窗生境下的光照强度相对较高,林缘其次,林下生境的光照强度最低,这一结果与前人的研究结果相似[17]。研究不同光环境下植物的光合特性可了解其对光照强度变化的生理适应性。柯世省等[18]研究表明,云锦杜鹃(Rhododendronfortunei)树苗对林窗和林缘不同强度光照存在不同的生理响应,林缘是林下云锦杜鹃最适宜的转移生境。
明确植物的光饱和点对植物的高效栽培至关重要。本文应用光响应机理模型可很好地拟合不同生境条件下棘茎楤木和树参叶片电子传递速率和净光合速率对光的响应曲线,而且得到的主要光合参数与实测值高度符合(R2>0.99),这与其他学者的结论相同[19,21]。光合速率对光的响应曲线结果表明,棘茎楤木和树参的光饱和点分别约为900和800 μmol·m-2·s-1;电子传递速率对光的响应曲线结果表明,棘茎楤木和树参的光饱和点范围分别为867.957~1 044.110 μmol·m-2·s-1和755.893~847.615 μmol·m-2·s-1,两个角度拟合的结果一致性较好。由此可见,棘茎楤木和树参均为半阳生植物,其最适宜光强约分别为900和800 μmol·m-2·s-1。这与吴晓云等[22]的相关研究结果基本一致。林下种植物种选择关键是其对林下弱光环境的适应能力[12]。光合速率对光的响应曲线拟合结果表明,棘茎楤木和树参叶片的光补偿点较低(约20 μmol·m-2·s-1),说明棘茎楤木和树参利用弱光的能力较强。另外,B和C生境下棘茎楤木和树参叶片光补偿点显著降低,说明棘茎楤木和树参可通过生理调节主动适应弱光环境。这与杨亚男等[23]对四季桂(Osmanthusfragrans)的研究结果相一致。由此可见,棘茎楤木和树参适合林下种植。
蒸腾作用是植物对水分吸收和运输的主要动力,可促进植物体对矿质元素的吸收,在高温条件下可通过水分蒸发带走大量热量,降低叶片的温度,从而减少高温对叶片的灼伤。熊慧等[24]发现,生境对植物气孔密度和长度有显著影响,由于林下光照强度低,相对湿度高与土壤含水量高,所以林下植物以较低的蒸腾成本可获得较大的光合效益,这与本试验结果一致。3种生境下的棘茎楤木和B生境下树参的蒸腾速率都随光照强度的增加而上升,随后趋于稳定,而A和C生境下树参的蒸腾速率在光照强度为1 500 μmol·m-2·s-1时未达到最大值。不同生境下棘茎楤木和树参叶片的蒸腾速率都存在显著差异,两种植物C生境下的蒸腾速率均处于最高。叶片气孔导度表示气孔张开的程度,这影响着植物的光合作用,蒸腾作用。本研究发现不同生境下棘茎楤木和树参叶片的气孔导度整体上均存在显著差异,以C生境下的值为最高。
光化学淬灭系数qP反映PSⅡ天线色素吸收的光能用于光化学电子传递的份额,并进一步反映PSⅡ反应中心的开放程度和电子传递活性的大小。苏金等[25]对不同遮阴下紫珠(Callicarpabodinieri)的光合特性研究表明,与全日照相比,遮阴处理会显著降低叶片的qP值。本试验结果显示,棘茎楤木和树参叶片qP在不同生境下整体上均无显著差异。另外,除用于光合作用的光能外,还有一部分光能可通过非光化学猝灭(qN)过程保护PSⅡ反应中心免受因吸收过多光能而引起的光氧化和光抑制伤害。本试验结果表明,不同生境下棘茎楤木叶片的qN值整体上无显著差异,而光强相对较弱的生境(B和C)下树参叶片qN值整体相对较高,说明棘茎楤木以及B和C生境下树参叶片具较好的耗散过量能量能力。ΦPSⅡ的明显降低是光抑制发生的可靠判据[26]。不同生境下的结果显示,A生境下棘茎楤木叶片ΦPSⅡ值整体较低,B和C生境下的ΦPSⅡ值极为接近;而3种生境下树参叶片的ΦPSⅡ值均较为接近。由此可见,A生境下的棘茎楤木发生了一定程度的光抑制。
综合本试验结果表明,林下生境的棘茎楤木和树参叶片的最大光合速率、蒸腾速率、气孔导度,以及最大电子传递速率、ΦPSⅡ和F′v/F′m值整体上均显著高于林窗和林缘生境;同时,在林窗和林缘生境下,棘茎楤木和树参也具有较强的耗散过量能量能力。结合叶片光饱和点和光补偿点范围,认为棘茎楤木和树参均属于半阳生树种,且两者对弱光的利用能力较强,适合林下种植。建议在浙中低海拔山区种植时,生境光强为800~900 μmol·m-2·s-1最为适宜。
