基于UPLC-MS非靶向代谢组学分析大鲵肉冷藏过程中代谢物的变化
2022-12-22刘俊霞兰阿峰裴金金陈德经金文刚
赵 萍,刘俊霞,兰阿峰,裴金金,陈德经,3,金文刚,3,*
(1.陕西理工大学生物科学与工程学院,秦巴生物资源与生态环境省部共建国家重点实验室,陕西 汉中 723001;2.陕西理工大学 陕西省资源生物重点实验室,陕西 汉中 723001;3.陕西理工大学 陕南秦巴山区生物资源综合开发协同创新中心,陕西 汉中 723001)
大鲵(Andrias davidianus),俗称“娃娃鱼”,属于动物界脊索动物门有尾目隐鳃鲵科,是世界上现存最大的两栖动物[1],迄今约有3.5亿 年的历史。大鲵具有极高的食用和药用价值,其皮肤、黏液、肌肉、油、骨骼等含有多种生物活性物质,具有抗氧化、抗肿瘤、抗菌、抗衰老等功效,是公认的宝贵生物资源,有“水中人参”之称[2]。随着大鲵养殖和繁殖技术的成熟,目前我国已在陕西汉中、湖南张家界、河南洛阳、重庆等多个省、市实现了规模化养殖[3]。近年来,研究人员已从大鲵的营养成分[4]、功能活性物质[5]、食用品质[6-7]、贮藏品质[8]等多方面进行了研究。但在大鲵肉贮藏保鲜方面研究较少,且主要集中在应用不同包装方式[8]、贮藏方式[9]、冻结方式[10]延长大鲵肉的货架期。
代谢组学是继基因组学、转录组学、蛋白质组学之后,系统生物学的重要组成部分,它是以高通量检测技术和多元数据处理为基础,通过研究小分子代谢物的变化,进而探究其代谢机制的新兴科学[11]。在食品领域代谢组学的主要分析技术包括核磁共振技术、气相色谱-质谱联用技术、液相色谱-质谱(liquid chromatographymass spectrometry,LC-MS)联用技术[12]等。其中LC-MS不需要对代谢物进行衍生化处理,即可进行定性和定量分析,具有分辨率高、灵敏度好、分析速度快等优点,适合沸点高、极性强的化合物分析[13],是目前通过非靶向确定代谢表型最广泛使用的方法[14]。超高效液相色谱(ultra-high performance LC,UPLC)是在高效液相色谱分离技术的基础上发展的,其在制造技术、扩散体积和耐受压力方面进行了优化,能在很大限度上发挥色谱柱的效能。其中Yu Qi等[15]基于UPLC-MS非靶向代谢组学揭示电子束辐照中华管鞭虾在冷藏过程中肌肉代谢的变化,获得两种潜在的生物标志物。Stella等[16]采用液相色谱-高分辨率质谱非靶向代谢组学技术对新鲜、冷冻、解冻鲈鱼进行研究,确定二十碳五烯酸和二十二碳六烯酸为最相关的预测因子,并以此建立模型以达到根据鲈鱼样品的贮藏条件进行分类的目的。Yu Qianqian等[17]利用LC-MS研究牛腰长肌和大腰肌之间肌肉的能量代谢,从代谢层面揭示肌肉到食用肉转化的内在机制。
低温保藏通常是既经济又实用的生鲜肉保鲜技术,同时也是开发其他保鲜技术的基础,降低温度一方面能减缓微生物的生长繁殖,另一方面可以降低酶活性[18]。但在冷藏过程中伴随着肌肉的僵直与解僵、蛋白质自溶、微生物生长繁殖等生化变化,进而导致肉的品质发生变化。课题组前期探究大鲵肉在冷藏过程中挥发性盐基氮(total volatile base nitrogen,TVB-N)、菌落总数等理化特性的变化并明确了冷藏期间主要致腐微生物菌属为假单胞菌属、气单胞菌属、Hafnia-Obesumbacterium和沙雷氏菌属[19-20]。在此基础上,本研究进一步采用UPLCMS非靶向代谢组学技术,结合多元统计方法筛选大鲵肉在冷藏过程中的差异代谢物,并分析差异代谢物的变化规律及相关代谢通路,为大鲵宰后肌肉代谢及冷藏期间品质调控提供一定参考。
1 材料与方法
1.1 材料与试剂
鲜活健康子二代大鲵3 尾,体质量(2.5±0.36) kg,购自汉中市龙头山大鲵养殖基地。
甲醇、甲酸、乙腈(均为色谱纯) 美国赛默飞世尔科技有限公司,L-2-氯苯丙氨酸 上海恒创生物科技有限公司。
1.2 仪器与设备
JXFSTPRP-24/32全自动样品快速研磨仪 上海净信实业发展有限公司;F-060SD超声波清洗机 深圳福洋科技集团有限公司;TGL-16MS台式高速冷冻离心机上海卢湘仪离心机仪器有限公司;LNG-T98冷冻浓缩离心干燥器 江苏苏州太仓市华美生化仪器厂;QE plus高分辨质谱仪、Dionex U3000 UHPLC超高效液相色谱仪美国赛默飞世尔科技有限公司;ACQUITY UPLC HSS T3(100 mm×2.1 mm,1.8 μm)色谱柱 美国沃特世公司。
1.3 方法
1.3.1 样品制备
鲜活大鲵经放血、热烫(95 ℃热水,处理约1 min)、刮黏液、去内脏和清洗后,用聚乙烯袋20 min内运回实验室。去头、皮、四肢、尾,取背肌切成约5.0 cm×2.0 cm×0.5 cm的肉块,放入黑色托盘中并用保鲜膜密封,置于4 ℃冰箱中冷藏。
以课题组前期对大鲵肉冷藏过程中TVB-N值和菌落总数等理化指标的研究结果为基础[19-20],分别于宰后冷藏第0(新鲜)、2(货架期之间)、4(货架期)、8天(腐败)时取样,每个取样时间点取6个平行,分别记为D0(D0-1、D0-2、D0-3、D0-4、D0-5、D0-6),D2、D4、D8的标记与D0相似。在取样时间点将样品取出,置于超净工作台上,去除保鲜膜,用事先灭菌的剪刀将肉样剪碎,混匀,用镊子将其放入15 mL离心管中,置于-80 ℃冰箱待用。
1.3.2 样品前处理
准确称取30 mg组织样品于1.5 mL EP管中,加入20 μL内标(0.3 mg/mLL-2-氯苯丙氨酸甲醇溶液)、600 μL甲醇-水溶液(4∶1,V/V),加入两个小钢珠。在-20 ℃冰箱中预冷2 min后,放入研磨机中60 Hz研磨2 min,经冰水浴超声提取(40 kHz、10 min)后,于-20 ℃冰箱中静置30 min。13 000 r/min、4 ℃离心10 min后,用注射器吸取150 μL上清液,使用0.22 μm的有机相针孔过滤器过滤后,转移到进样小瓶,置于-80 ℃冰箱保存。
质控(quality control,QC)样品由所有样品的提取液等体积混合制备而成。
1.3.3 UPLC-MS分析条件
UPLC条件:ACQUITY UPLC HSS T3色谱柱(100 mm×2.1 mm,1.8 μm);柱温:45 ℃;流动相:A:水(含0.1%甲酸),B:乙腈(含0.1%甲酸);流速:0.35 mL/min;进样体积:2 μL。梯度洗脱程序:0~2 min,95% A、5% B;2~4 min,95%~70% A、5%~30% B;4~8 min,70%~50% A、30%~50% B;8~10 min,50%~20% A、50%~80% B;10~14 min,20%~0% A、80%~100% B;14~15 min,0% A、100% B;15~15.1 min,0%~95% A、100%~5% B;15.1~16 min,95% A、5% B。
MS条件:采用电喷雾离子源,在正离子和负离子扫描模式下采集数据,喷雾电压分别为3 800 V和-3 000 V,其他参数正负离子模式下相同。毛细管温度:320 ℃;辅助器加热温度:350 ℃;鞘气流速:35 arb;辅助气流速:8 arb;离子透镜射频电压:50 V;质量扫描范围:m/z100~1 200;全MS分辨率:70 000;MS/MS分辨率:17 500;归一化碰撞能量:10、20、40 eV。
1.4 数据处理与分析
1.4.1 数据处理
在进行模式识别之前,原始数据使用Progenesis QI v2.3代谢组学处理软件进行基线过滤、峰识别、积分、保留时间校正、峰对齐和归一化处理,主要参数如下:母离子质量容差:5×10-6;子离子质量容差:10×10-6;子离子阈值:5%(物质峰的偏离程度)。化合物的鉴定基于精确质量数、二级碎片以及同位素分析,使用EMDB数据库进行定性。对得到的数据进行进一步处理,删除组内缺失值(0值)>50%的离子峰,将0值以最小值的一半替换,根据化合物定性打分,对定性得到的化合物进行筛选,筛选标准为36 分(满分60 分),36 分以下视为定性结果不准确并删除。最后将正负离子数据合并成一个数据矩阵表,该矩阵包含了从原始数据提取到的所有可用于分析的信息。
1.4.2 数据分析
以1.4.1节的数据矩阵为基础,利用SIMCA 14.1软件进行主成分分析(principal component analysis,PCA)、偏最小二乘判别分析(partial least squares-discriminant analysis,PLS-DA);通过t-检验和变异倍数(fold change,FC)分析比较两组之间代谢物数量差异;采用SPSS 25软件进行显著性分析,其中多重比较采用沃特-邓肯法,P<0.05表示差异显著;使用Origin 2021软件绘图,并利用京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)Pathway数据库(https://www.kegg.jp/kegg/pathway.html)进行代谢通路分析。
2 结果与分析
2.1 QC样品分析

图1 QC样品正离子(A)和负离子(B)基峰图Fig. 1 Positive ion (A) and negative ion (B) base peak chromatograms of QC samples
在检测过程中,QC样品用于评价质谱系统的稳定性,以获得可靠且高质量的代谢组学数据。如图1所示,QC样品的基峰图基线稳定,说明仪器数据采集稳定性很好。此外,正负离子模式下出峰的响应强度、峰面积及峰数量有所不同。
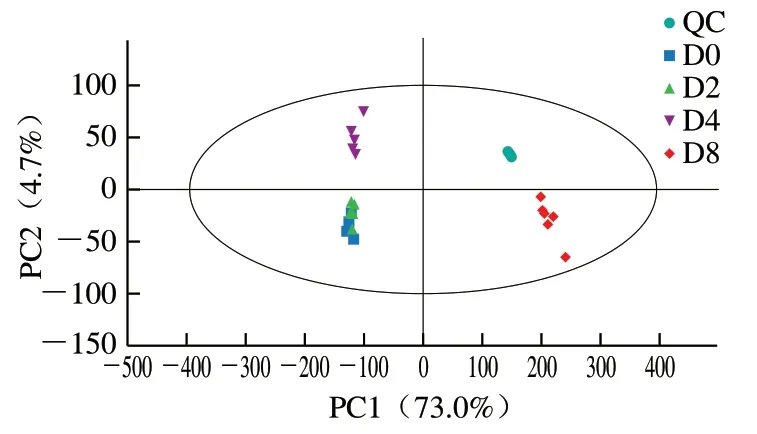
图2 QC组和实验组PCA得分图Fig. 2 PCA score plot of QC and experimental groups
对QC样品进行相对标准偏差(relative standard deviation,RSD)筛选,删除QC样品中RSD>0.4的离子峰,并采用PCA法反映组间和组内样品之间变异度的大小,同时对系统稳定性进行评价。如图2所示,在95%置信区间冷藏4、8 d样品组间分离度良好;而冷藏0、2 d样品出现组间重叠现象,说明冷藏0、2 d样品组间差异较小。各组内样品之间相互靠拢,表明样品组内变异度较小。QC样品紧密聚集在一起,表明实验稳定性和重复性较好,满足继续实验的条件。
2.2 不同冷藏时间大鲵肉的多元统计分析
2.2.1 不同冷藏时间大鲵肉的PCA

图3 不同冷藏时间大鲵肉样品的PCA得分图Fig. 3 PCA score plot of giant salamander meat samples with different refrigeration time
PCA因无外加人为因素,其模型反映了代谢组学数据的原始状态,有利于掌握数据的整体情况并对数据从总体上进行把握,尤其有利于发现和剔除异常样品,并提高模型的准确性。如图3所示,说明拟合性较好,Q2=0.724>0.5,且与相差较小,说明模型的稳定性较好。与冷藏0、2 d样品相比,冷藏4、8 d组内样品分散程度较高,说明(0~2)、4、8 d之间样品差异较大。
2.2.2 不同冷藏时间大鲵肉的PLS-DA

图4 不同冷藏时间大鲵肉样品PLS-DA得分图(A)和响应排序检验图(B)Fig. 4 PLS-DA score plot (A) and response permutation test plot (B) of giant salamander meat samples with different refrigeration time
非监督的PCA能够直观显示样品间的分组、变化趋势和相似性关系,但差异变量会分布在PC上,无法在后面的分析中将其剔除做更准确的分析,同时对相关性较小的变量不敏感[21]。PLS-DA是一种有监督的判别统计方法,能实现对样品类别的预测,加入了分组变量可弥补PCA的不足,模型中分别表示所建模型对X和Y矩阵的解释率,Q2表示模型的预测能力,这些值越接近于1表示模型的拟合度越好。如图4A所示,Q2的值分别为0.842、0.984、0.903,证明PLS-DA模型能更好地解释和预测样品之间的差异,模型的预测能力较好。为防止模型过拟合,对PLS-DA模型进行7次循环交互验证和200次响应排序检验考查模型的质量,其中R2、Q2为回归直线与y轴的截距值,R2表示模型能够解释的方差总和,Q2表示模型的预测能力,使用响应排序检验时,一般要求Q2小于零。如图4B所示,R2=0.692,Q2=-0.321,回归线呈向上的趋势,没有出现过拟合的现象,模型可靠,可用于后续的分析。
2.3 不同冷藏时间大鲵肉的单变量统计分析

图5 不同冷藏时间大鲵肉样品中差异代谢物火山图Fig. 5 Volcano diagrams of differential metabolites between giant salamander meat samples with different refrigeration time
单变量分析主要集中在单变量的描述(描述样品数据的集中和离散趋势)和统计推断(从样品资料推断总体情况)两个方面,用t-检验和FC分析比较两组之间的代谢产物数量差异。以P<0.05,log2FC绝对值>1为标准,筛选差异代谢产物。如图5所示,红色圆点代表在比较组中显著上调的代谢产物,绿色圆点代表显著下调的代谢产物,蓝色圆点代表不显著的代谢产物。筛选结果显示,冷藏0 d与2 d、2 d与4 d、4 d与8 d的样品中差异表达代谢物分别为1 223(上调和下调代谢物分别为603、620个)、1 780(上调和下调代谢物分别为1 379、401个)、16 761个(上调和下调代谢物分别为9 676、7 085个),显著差异代谢物的个数随冷藏时间的延长呈增加的趋势;3 组对比组样品中上调代谢物分别占其显著差异代谢物总数的49.30%、77.47%、57.73%,下调代谢物分别占其显著差异代谢物总数的50.70%、22.53%、42.27%。
2.4 不同冷藏时间大鲵肉中差异代谢物的筛选与鉴定
采用多维分析和单维分析相结合的方法,筛选组间差异代谢物。PLS-DA中,变量投影重要性指标(variable importance in projection,VIP)值可用来衡量各代谢物表达模式对各组样品分类判别的影响强度和解释能力。以PLS-DA模型PC1的VIP值≥2,t-检验的P≤0.001为标准进行进一步筛选,最终共得到125种差异代谢物,如表1所示。同时利用Origin绘制125种差异代谢物的热图,如图6所示,热图聚类分析可将冷藏0、2 d组聚为一类,4 d组聚为一类,8 d组聚为一类,冷藏8 d样品与其他组样品有明显差异。

图6 不同冷藏时间大鲵肉样品中差异代谢物热图Fig. 6 Heatmap of differential metabolites between meat samples of giant salamander with different refrigeration time
从表1观察到17种有机酸类及其衍生物,其中肌酸的VIP值最高为23.13,其次为葡萄糖酸(VIP 8.95)、2,3-二氨基水杨酸(VIP 8.66),其余有机酸类及其衍生物的VIP值均小于6。将17种代谢物划分为A1、B1两类,其中A1类化合物在冷藏8 d明显降低;与冷藏0 、2 d相比,Bl类化合物在冷藏4 d和8 d明显增加。对A1、B1类化合物进行累积统计分析,其变化趋势如图7所示,A1类化合物累积丰度在冷藏0~2 d出现小幅度的上升,冷藏2~4 d出现小幅度的下降,冷藏4~8 d快速下降;B1类化合物累积丰度在整个冷藏过程中较A1类化合物波动小,但在冷藏4 d明显向上波动。由表1可知,肌酸在脊柱动物体内,能够辅助为肌肉和神经细胞提供能量,由精氨酸、甘氨酸及甲硫氨酸合成,其丰度在冷藏0、2、4 d较高,在冷藏8 d显著下降(P<0.05)。葡萄糖酸是磷酸肌醇代谢的终产物,能进入磷酸戊糖途径参与代谢,其丰度在冷藏4 d达到最大值。课题组前期的实验结果表明,大鲵肉在冷藏后期(6、8 d)的优势菌属为假单胞菌属[20],由表1可以看出,在冷藏8 d绝大部分有机酸类及其衍生物(A1类化合物)和氨基酸类及其衍生物(A3类化合物)丰度明显降低,可能是由于假单胞菌属的首选碳源为有机酸和氨基酸,而不是葡萄糖[22]。
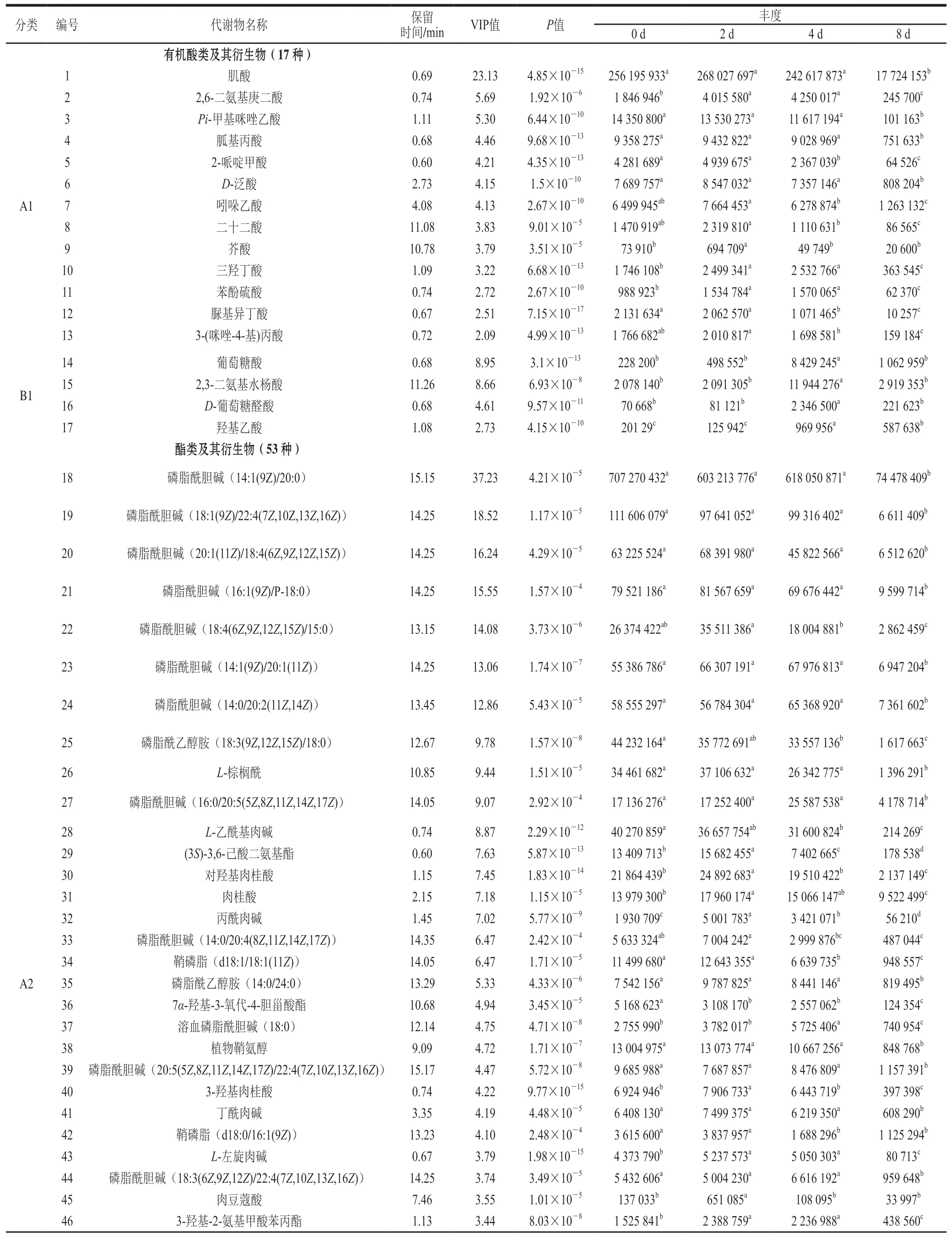
表1 不同冷藏时间大鲵肉样品差异代谢物筛选结果Table 1 Results of screening of differential metabolites in giant salamander meat samples with different refrigeration time

续表1

续表1

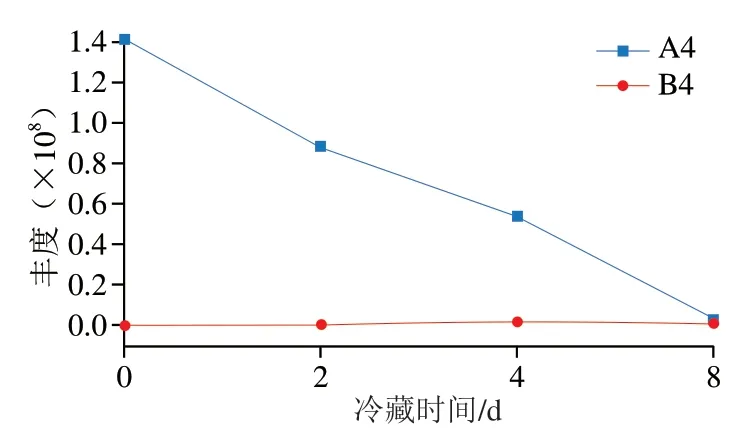
图7 大鲵肉冷藏期间各类差异代谢物的变化Fig. 7 Changes in abundance of differential metabolites in giant salamander meat during cold storage

图8 不同冷藏时间大鲵肉样品差异代谢物中6 类酯类化合物丰度Fig. 8 Abundance of six differential ester compounds in giant salamander meat samples at different cold storage time
酯类及其衍生物共53种,是不同冷藏时间大鲵肉样品中差异代谢物数量最多的一类化合物。所有酯类及其衍生物主要包括14种溶血磷脂酰胆碱类化合物(lysophosphatidylcholine,LysoPC)、13种磷脂酰胆碱类化合物(phosphatidyl choline,PC),2种磷脂酰乙醇胺类(phosphatidyl ethanolamine,PE),2种鞘磷脂类(sphingomyelin,SM),1种磷脂酸(phosphatidic acid,PA),1种溶血鞘磷脂(LysoSM)及一些其他的酯类化合物。如图8所示,PC、PE、SM的丰度随着冷藏时间的延长呈降低的趋势,其中,PC类的丰度在6 类酯类化合物中占有绝对优势;LysoPC类冷藏4 d丰度最高,其次为8 d, 0、2 d丰度较低。所有酯类化合物中PC(14:1(9Z)/20:0)、LysoPC(18:1(11Z))的VIP值最大,分别为37.23、30.63,其次为LysoPC(20:4(5Z,8Z,11Z,14Z))(VIP 22)、PC(18:1(9Z)/22:4(7Z,10Z,13Z,16Z))(VIP 18.52)、LysoPC(16:0)(VIP 16.48)、PC(20:1(11Z)/18:4(6Z,9Z,12Z,15Z))(VIP 16.24)、LysoPC(20:4(8Z,11Z,14Z,17Z))(VIP 15.93)、PC(16:1(9Z)/P-18:0)(VIP 15.55)、PC(18:4(6Z,9Z,12Z,15Z)/15:0)(VIP 14.08)、PC(14:1(9Z)/20:1(11Z))(VIP 13.06)、PC(14:0/20:2(11Z,14Z))(VIP 12.86),其余酯类及其衍生物的VIP值均小于10。将53种酯类及其衍生物划分为A2、B2两类,其中A2类化合物在冷藏8 d明显降低;与冷藏0 d和2 d相比,B2类化合物在冷藏4、8 d明显增加。如图7所示,A2类化合物累积丰度在0~4 d缓慢下降,4~8 d快速下降;B2类化合物累积丰度在0~2 d波动不明显,2~4 d显著上升,4~8 d显著下降。
脂质氧化是导致肉类中脂肪组织恶化的主要原因,在生物系统中,脂质通过光氧化、酶氧化和自氧化3种途径氧化[23]。PC又称卵磷脂,是生命体中分布最广泛的磷脂,由表1可知,PC(18:1(9Z)e/2:0)在冷藏4、8 d丰度较0、2 d高且变化明显(P<0.05);其他PC类化合物在冷藏0、2、4 d丰度较高,在冷藏8 d丰度显著下降(P<0.05),与Yu Qi等[15]的研究结果相似,PCs作为所有质膜的一部分,发挥着重要的作用,对新细胞的生长至关重要[24-25],多数PC在冷藏8 d显著下降(P<0.05)标志着大鲵肉品质的显著下降。同时由于PCs分子中常有软脂酸、硬脂酸、油酸、亚油酸、花生四烯酸等,因此,大鲵肉冷藏过程中,PC能在酶的作用下分解产生花生四烯酸参与没食子酸的代谢,也可直接在酶的作用下分解为α-亚麻酸、亚油酸等物质[15]。LysoPC是PC代谢的中间产物,除LysoPC(18:0)在冷藏0、2、4 d丰度较冷藏8 d高且变化显著(P<0.05),其余LysoPC冷藏4、8 d丰度较高。PE俗称脑磷脂,分子中的脂肪酸常有软脂酸、硬脂酸、油酸及少量花生四烯酸,PE与PC的性质相似,变化趋势也与PC相似,在冷藏0、2、4 d丰度较高,在冷藏8 d丰度显著下降(P<0.05)。PA是最简单的二酰基甘油磷脂,细胞膜的组成成分,在冷藏0、2、4 d丰度较高,在冷藏8 d丰度显著下降(P<0.05)。脂质氧化主要发生在脂肪酸中,而存在于细胞膜和亚细胞结构中的磷脂是脂质氧化的良好底物,这可能是多数PC、PA、PE等磷脂类化合物在冷藏8 d显著下降的主要原因[26]。SM是细胞膜与细胞生长、存活、死亡相关的结构中普遍存在的组成部分,同时也是控制免疫效应的信号代谢物[27],在冷藏0、2 d丰度显著高于冷藏4、8 d(P<0.05),表明随着冷藏时间的延长,大鲵肉逐渐丧失自身的免疫、抗菌等功能特性。
氨基酸类及其衍生物共25种,L-色氨醇的VIP值最高为17.67,其次为谷胱甘肽(VIP 10.12),L-组氨酸(VIP 9.10)、L-正亮氨酸(VIP 7.22),其他氨基酸类及其衍生物的VIP值均小于7。将25种代谢物划分为A3、B3两类,其中A3类化合物在冷藏8 d明显降低,B3类化合物在冷藏0、8 d丰度较高。如图7所示,A3类化合物与A1类化合物累积丰度变化趋势相似,即在冷藏0~2 d缓慢上升,2~4 d缓慢下降,4~8 d迅速下降;B3类化合物即亮氨酰-亮氨酸,在整个冷藏过程中波动幅度不大。
除亮氨酰-亮氨酸外,其余氨基酸类化合物在冷藏8 d丰度较低,表明大鲵肉在冷藏过程中营养流失。在冷藏前期内源性蛋白水解酶活性较高,肌原纤维蛋白被肌肉蛋白酶(钙蛋白酶、催化蛋白酶等)分解成多肽片段,然后由肽基肽酶(二肽基肽酶、三肽基肽酶、氨基肽酶等)分解产生游离氨基酸[28],在冷藏后期丰度显著下降,可能是由于以假单胞菌属为主的致腐菌属生长繁殖消耗[22]。通过KEGG代谢通路分析发现,L-精氨酸、鸟氨酸、L-谷氨酸主要参与必需氨基酸精氨酸的生物合成和分解代谢,在冷藏0、2 d丰度最高(表1);L-组氨酸、L-鹅肌肽主要参与组氨酸代谢,其中鹅肌肽为肌肽的甲基化衍生物,为组氨酸二肽,具有显著的抗氧化作用,是肉类的内源性抗氧化剂,对保持肉类品质[29]、形成良好的风味和细嫩的质地具有重要作用[30],这些物质的丰度随着冷藏时间的增加而降低(表1),表明大鲵肉自身抗氧化性降低;大部分氨基酸主要参与氨酰-tRNA生物合成和ABC-转运器。所有筛选出来的氨基酸类及其衍生物中没有明显的代谢物导致大鲵肉品质恶化,但游离氨基酸可能是细菌的生长促进剂[31]。
核苷酸类及其衍生物13种,别嘌呤醇核糖苷的VIP值最大为15.76,其次为次黄嘌呤(VIP 12.07)、阿糖肌苷(VIP 11.77)、肌苷(VIP 11.16),其余核苷酸类及其衍生物的VIP值均小于7。将13种核苷酸类及其衍生物分为A4、B4两类,其中A4类化合物在冷藏8 d明显降低,与冷藏0、2 d相比,B4类化合物在冷藏4、8 d明显上升。如图7所示,A4类化合物累积丰度在冷藏过程中呈降低的趋势,且趋势接近线性,而B4类化合物在冷藏4、8 d出现波动,但波动幅度不大。
核苷酸类及其衍生物中二磷酸腺苷和2'-脱氧鸟苷-5'-二磷酸三钠盐在冷藏0、2、4 d丰度无显著差异(P>0.05),在冷藏8 d显著降低(P<0.05)(表1)。在冷藏过程中三磷酸腺苷主要由磷酸肌酸途径和无氧糖酵解途径产生,随后三磷酸腺苷逐渐降解,降解的一般过程为[32]:三磷酸腺苷→二磷酸腺苷→单磷酸腺苷→次黄嘌呤核苷酸→肌苷→次黄嘌呤→黄嘌呤→尿酸。
随着三磷酸腺苷的降解,二磷酸腺苷、2'-脱氧鸟苷-5'-二磷酸三钠盐的丰度逐渐增加,并分别在冷藏2、4 d丰度达到最大值后逐渐降低。次黄嘌呤、肌苷、5'-肌苷酸在冷藏0 d丰度最高,之后显著降低(P<0.05),在冷藏8 d达到最小值;而黄嘌呤随着冷藏时间的延长呈现先增加后降低的趋势,在冷藏4 d达到最大值后降低,但冷藏8 d丰度仍大于冷藏0、2 d。次黄嘌呤已被确定为水产品新鲜度的潜在生物标志物[33],次黄嘌呤是一种与炎症和氧化应激相关的代谢标志物,能通过增加内源性黄嘌呤氧化酶活性提高抗菌性能,但在本实验中次黄嘌呤下调可能是大鲵肉细胞的抗菌特性减弱,同时作为三磷酸腺苷的分解产物,其下调也可能与三磷酸腺苷总量的减少有关[34]。
醇类及其衍生物3种,均在冷藏8 d丰度最低且变化显著(P<0.05);反-2-戊醇的VIP值最大为3.95,其次为氧化芳樟醇(VIP 3.04),雄甾烷-3α,17β-二醇(VIP 2.29)。其他类化合物共14种,以去甲基西酞普兰的VIP值最大为5.10,其余化合物的VIP值均小于4。尸胺(1,5-二氨基戊烷)在冷藏2 d开始被检出,与冷藏0 d无显著差异(P>0.05),在冷藏4 d显著增加(P<0.05)并达到最大值。这可能与该冷藏期微生物的种类和丰度有关,王纯纯等[35]研究表明,尸胺的产生与沙雷氏菌属、短乳杆菌、片球菌等菌属密切相关,同时假单胞菌属、乳酸杆菌等可促进尸胺等生物胺的形成,课题组前期对微生物多样性的研究结果揭示[20],在冷藏4 d沙雷氏菌属的丰度最高,除此之外假单胞菌属、Hafnia-Obesumbacterium等也有相对较高的丰度,这些菌属在冷藏4 d的变化有利于尸胺的形成。尸胺是一种对人体有害的化合物,但是它对人体健康的危害需要达到一定的量,同时尸胺作为一个单一的生物胺代谢物,不能单独作为评价大鲵肉品质的指标,仍需结合多个指标综合进行判定,但该物质在冷藏后期的增加,表明大鲵肉的品质发生了劣变。
2.5 不同冷藏时间大鲵肉中代谢通路富集分析
大鲵肉冷藏过程中品质的变化受多种物质和反应共同调控,不能仅从某一物质含量的高低进行整体判断,因此仍需进一步对其代谢通路进行分析。通过KEGG数据库比对,富集到80 条代谢通路中的差异代谢物共144种,其中差异显著(P<0.05)的代谢通路共19 条,富集到这19 条代谢通路的差异代谢物共54种,基本信息如表2所示,差异代谢通路的总体情况如图9所示。

表2 不同冷藏时间大鲵肉中差异代谢通路基本信息Table 2 Basic information of differential metabolic pathways in giant salamander meat at different cold storage time

续表2

图9 不同冷藏时间大鲵肉中差异代谢通路富集气泡图Fig. 9 Enrichment analysis of differential metabolic pathways in giant salamander meat at different refrigeration time
气泡图中,颜色由黄到蓝表示P值依次降低;气泡越大,表明富集到该通路的差异代谢物数目越多。由表2及图9可知,差异极显著的前5 条通路分别为组氨酸代谢、精氨酸和脯氨酸代谢、嘌呤代谢、精氨酸生物合成、鞘脂类代谢;富集差异代谢物数量最多的5 条通路分别为嘌呤代谢、精氨酸和脯氨酸代谢、组氨酸代谢、赖氨酸降解、鞘脂代谢;富集的差异代谢物中L-谷氨酸、L-精氨酸、L-亮氨酸、组氨酸、鸟氨酸、酮戊二酸、二磷酸腺苷、三磷酸腺苷等参与多条差异代谢通路。所有代谢通路中氨基酸类代谢通路最多,为7 条。
2.6 相关性分析
2.6.1 代谢通路与微生物、理化指标进行相关性分析
对19 条差异显著(P<0.05)的代谢通路与课题组之前的研究结果(大鲵肉在相同冷藏条件下的致腐微生物菌属、TVB-N值、菌落总数)进行Pearson相关性分析[19-20],结果如图10所示。结果表明,磷酸戊糖途径与其他代谢途径之间均无显著相关(P>0.05);与铁死亡、谷胱甘肽代谢、坏死性凋亡、嘌呤代谢通路呈显著相关(P<0.05)的代谢通路较少;而与氨基酸类代谢通路(组氨酸代谢、精氨酸代谢、赖氨酸降解、牛磺酸和次牛磺酸代谢、缬氨酸、亮氨酸和异亮氨酸生物合成)和翻译通路(氨酰-tRNA生物合成)显著(P<0.05)相关的代谢通路较多。除嘌呤代谢、坏死性凋亡、谷胱甘肽代谢、铁死亡代谢通路、磷酸戊糖途径以外,其余代谢通路与TVB-N值均显著(P<0.05)负相关。除溶酶体、铁死亡、促性腺激素信号通路、磷酸戊糖途径以外,其余代谢通路与菌落总数均显著(P<0.05)负相关。致腐微生物总体与代谢通路之间的相关性较差,仅坏死性凋亡与气单胞菌属、Hafnia-Obesumbacterium之间显著(P<0.05)负相关,但菌落总数与多条代谢通路的相关性较强,可能是由于代谢物的富集受多种微生物共同作用,单一微生物发挥的作用较小。综上并结合表2,组氨酸代谢、精氨酸和脯氨酸代谢、精氨酸生物合成、赖氨酸降解、牛磺酸和次牛磺酸代谢、缬氨酸、亮氨酸和异亮氨酸生物合成、氨酰-tRNA生物合成、鞘脂类代谢、细胞间隙连接、神经活性配体-受体相互作用代谢通路与大鲵肉品质的变化关系密切,目前,也有研究指出嘌呤代谢和组氨酸代谢是与腐败显著相关的代谢通路[15,36]。
2.6.2 代谢物与微生物、理化指标进行相关性分析
选取19 条差异代谢途径中VIP值≥3、P<0.05的代谢物与课题组之前的研究结果(大鲵肉在相同冷藏条件下的致腐微生物菌属、TVB-N值、菌落总数)进行Pearson相关性分析[19-20],结果如图11所示。

图10 不同冷藏时间大鲵肉中差异代谢通路、微生物及理化指标Pearson相关性分析Fig. 10 Pearson correlation analysis of differential metabolic pathways, microorganisms and physicochemical indexes in giant salamander meat at different refrigeration time

图11 不同冷藏时间大鲵肉中差异代谢物、微生物及理化指标Pearson相关性分析Fig. 11 Pearson correlation analysis of differential metabolites, microorganisms and physicochemical indexes in giant salamander meat at different refrigeration time
由图11可知,次黄嘌呤、肌苷、谷胱甘肽、5'-肌苷酸、鸟苷之间相关性较强;肌酸、L-组氨酸、L-谷氨酸、组氨酸、1-甲基-L-组氨酸、鸟氨酸、L-精氨酸、植物鞘氨醇、L-脯氨酸、二磷酸腺苷之间相关性较强,其都与TVB-N值显著(P<0.05)负相关,以肌酸、L-组氨酸、组氨酸、1-甲基-L-组氨酸、植物鞘氨醇的相关性最强;(3S)-3,6-己酸二氨基酯、SM(d18:1/18:1(11Z))、2-哌啶甲酸、鸟嘌呤之间相关性较强。L-谷氨酸与菌落总数之间具有极显著(P<0.01)相关,肌酸、L-组氨酸、组氨酸、鸟氨酸、L-精氨酸、植物鞘氨醇、(3S)-3,6-己酸二氨基酯、SM(d18:1/18:1(11Z))、2-哌啶甲酸与菌落总数之间具有显著(P<0.05)相关。与理化指标相比,致腐微生物与差异代谢物之间的相关性不强,其中鸟嘌呤与假单胞菌属之间具有显著(P<0.05)负相关,(3S)-3,6-己酸二氨基酯、SM(d18:1/18:1(11Z))、2-哌啶甲酸、鸟嘌呤与气单胞菌属、Hafnia-Obesumbacterium之间具有显著(P<0.05)负相关。综上并结合表2,肌酸、L-组氨酸、L-谷氨酸、组氨酸、鸟氨酸、L-精氨酸、植物鞘氨醇可作为大鲵肉冷藏过程中品质变化的潜在生物标记物。
3 结 论
采用UPLC-MS非靶向代谢组学技术对大鲵肉冷藏过程中(4 ℃,0、2、4、8 d)肌肉小分子代谢物进行研究。通过多元统计分析表明,在冷藏过程中,冷藏0、2 d组间和组内差异较小,冷藏4、8 d组间和组内差异相对较大;单变量统计分析表明,随着冷藏时间的延长,上调代谢物的个数呈增加趋势,下调代谢物的个数呈先减少后增加的趋势;以VIP值≥2、P≤0.001为筛选标准,共筛选出125种差异代谢物,包括有机酸类及其衍生物(17种)、酯类及其衍生物(53种)、氨基酸类及其衍生物(25种)、核苷酸类及其衍生物(13种)等;对差异代谢物的变化规律进行分析发现,大部分差异代谢物在冷藏8 d丰度明显下降;代谢通路富集及Pearson相关性分析表明,组氨酸代谢、精氨酸和脯氨酸代谢、精氨酸生物合成、赖氨酸降解、牛磺酸和次牛磺酸代谢、缬氨酸、亮氨酸和异亮氨酸生物合成、氨酰-tRNA生物合成、鞘脂类代谢、细胞间隙连接、神经活性配体-受体相互作用代谢通路与大鲵肉品质的变化关系显著;肌酸、L-组氨酸、L-谷氨酸、组氨酸、鸟氨酸、L-精氨酸、植物鞘氨醇可作为大鲵肉冷藏过程中品质变化的潜在生物标记物。该研究为大鲵肉宰后肌肉代谢及冷鲜肉品质控制提供了参考。
