黑莓果实发育过程中蔗糖磷酸合成酶基因的表达分析
2022-11-29严志祥杨海燕樊苏帆吴文龙闾连飞李维林
严志祥,杨海燕,樊苏帆,吴文龙,闾连飞,李维林*
(1.南京林业大学林学院,南方现代林业协同创新中心,江苏 南京 210037;2.江苏省中国科学院植物研究所,江苏 南京 210014)
目前,分子育种是经济林研究的重要技术手段之一,具有广阔的发展前景[1]。蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)是由2个或4个亚基构成的二聚体或四聚体[2],其主要催化尿苷二磷酸葡萄糖(UDPG)和果糖-6-磷酸(F-6-P)反应生成6-磷酸蔗糖,而后进一步被蔗糖磷酸化酶(SPP)水解为蔗糖和磷酸根离子。SPS与SPP在植物中通常以复合体的形式存在,所以蔗糖生成是不可逆的[3]。SPS是蔗糖合成路径上的关键酶之一,在植物组织器官中广泛存在,调控光合产物在蔗糖和淀粉间的分配,同时是蔗糖合成的关键限速酶。SPS在植物生长发育、果实成熟、细胞壁合成、果实品质形成和同化产物的积累与分配等方面具有重要作用。此外,SPS还对植物的抗逆性有一定影响[4]。
在植物中发现的A、B、C、D 4个SPS基因亚族[5]中,除了D亚族仅存在于单子叶植物中[6],其他亚族成员在单、双子叶植物中均有发现。A亚族广泛存在于植物的各个组织中,B亚族主要存在于植物的生殖器官,C亚族主要存在于植物的成熟组织中。目前,在拟南芥(Arabidopsisthaliana)中共发现4个SPS基因,分别为SPSA1、SPSA2、SPSB和SPSC,其中SPSA1主要在光合作用合成蔗糖过程中发挥重要作用[7],SPSA1参与花粉末期蔗糖合成[8],SPSA1和SPSA2在成熟蜜腺中大量表达,表明其与花蜜合成关系密切[9]。在烟草(Nicotianatabacum)中,SPSB在生殖器官中大量表达[10];SPSC在拟南芥及烟草中均有发现,其主要负责夜间蔗糖合成以及维持淀粉正常运输[10]。SPS还能促进光合产物向蔗糖流动而降低淀粉的合成。已有研究表明,在荔枝(Litchichinensis)假种皮发育过程中LcSPS4与蔗糖含量呈现相同的变化趋势[11]。也有研究发现,甜瓜(Cucumismelo)中CmSPS1随着果实发育表达量不断提高,暗示其影响甜瓜果实发育和蔗糖代谢[12]。Verma等[13]对不同品种甘蔗(Saccharumofficinarum)的研究发现,高糖品种中SPS基因表达量更高,进一步证明SPS与蔗糖含量呈正相关。
黑莓(Rubusspp.)属蔷薇科悬钩子属灌木[14],是近年风靡全球的第3代水果,其果实糖含量不显著,口感较酸。黑莓中糖的种类及含量直接决定其品质。蔗糖是主要的可溶性糖之一,它的运输和分配不仅决定着植物生长发育的整个过程,也决定着经济器官的产量和质量,同时它改善了水果的味道,是水果甜味最重要的组成部分。SPS是蔗糖合成的关键限速酶之一,其活力大小与蔗糖合成呈显著正相关[15]。目前关于黑莓中SPS基因与蔗糖合成的分子机制研究的相关报道鲜见。本研究利用生物信息学分析方法, 基于前期通过转录组测序信息克隆得到的3个SPS基因,对其编码的蛋白结构、进化关系以及在植株内的表达情况等进行研究,以期为黑莓中SPS家族主要成员相关功能的深入研究提供参考。
1 材料与方法
1.1 研究材料
黑莓品种‘宝森’(‘Boysenberry’)栽培于江苏省南京市溧水区白马黑莓科研基地。试验于2019年4月下旬至6月上旬进行。选择10株生长发育良好的3年生健康植株,在植株达到开花的旺盛期[15],标记当日植株盛开的发育良好花朵,从花后21 d直至花后39 d,每7 d随机采集1次试验果实样品,同步收集当日发育良好的叶片样品,进行SPS基因的表达分析和SPS酶活性测定。分别于盛花期采集植株的根、茎、叶和花, 在液氮中冷冻而后存于-80 ℃待测。
1.2 黑莓SPS基因表达分析方法
1.2.1 黑莓SPS基因的序列及系统进化分析方法
利用美国国家生物信息中心(https://www.ncbi.nlm.nih.gov/)网站提供的相关信息进行SPS的cDNA序列比对;氨基酸序列同源性多重比对利用Clustal X 2.0进行。使用MEGA 7.0软件中的邻接法(neighbour-joining, NJ)构建氨基酸序列进化树,其中校验参数bootstrap为1 000次重复。
1.2.2 黑莓SPS蛋白的特性和结构分析方法
通过SMART网站(http://smart.embl.de/)分析蛋白保守功能域;利用MEME网站(http://meme-suit.org/tools/meme)分析黑莓SPS蛋白的保守元件,元件注释利用InterProScan 网站(http://www.ebi.ac.uk/interpro/search/sequence-search)进行;使用ExPASy Proteomics Sever(http://expasy.org/)网站进行黑莓SPS编码的蛋白氨基酸序列分子量和等电点预测;通过Prabi网站(https://prabi.ibcp.fr/htm/site/web/home)及Swissmodel网站(https://swissmodel.expasy.org/)对各SPS基因所编码的氨基酸序列进行推导,预测蛋白质的二级和三级结构;利用Genedoc软件进行氨基酸序列比对分析。
1.2.3 黑莓SPS基因qRT-PCR分析方法
RNA提取试剂盒购自北京百泰克公司,分别提取‘宝森’根、茎、叶片、花和果实的总RNA,利用上海浦迪生物科技有限公司反转录试剂制备cDNA,根据试剂盒说明,1 μL RNA 反转录成10 μL cDNA。通过qRT-PCR检测蔗糖代谢相关基因在不同组织中的表达水平,各基因序列特异性引物见表1。qRT-PCR 扩增体系为20 μL,其中cDNA 1 μg,TB GreenTMPremix ExTaqTM(TaKaRa)10 μL,引物各0.5 μL,扩增程序为:95 ℃ 2 min,95 ℃ 10 s,60 ℃ 10 s,72 ℃ 15 s,延伸6 s;40个循环。根据定量扩增曲线达到平台期时的Ct值,以黑莓Actin基因(GenBank No.HQ439556)作为内参。

表1 qRT-PCR引物序列
1.2.4 黑莓SPS酶的活性测定
SPS活性测定采用上海优选生物科技有限公司试剂盒。将每mg组织蛋白每min催化产生1 μg蔗糖定义为1个酶活力单位,μg/(min·mg)。测定3次重复。
1.3 数据处理
所有试验数据均进行 3 次生物学重复及技术重复,采用Excel 2016 及DPS 7.05软件进行数据统计和差异显著性分析,采用 SPSS 22.0 以 Pearson 相关模型分析基因表达量和酶活力的关系。
2 结果与分析
2.1 黑莓SPS基因系统发生分析
利用NCBI BLAST工具对克隆获得的3条cDNA核苷酸序列片段进行分析,发现其编码的是不同形式的SPS,将序列信息提交至GenBank数据库中,分别命名为RuSPS1(MT682348)、RuSPS2(MT682349)和RuSPS3(MT682350)。对RuSPSs基因编码的蛋白进行分析发现,其氨基酸长度为851~1 068 aa,分子质量为93.98~119.94 ku,等电点从5.96到6.16不等,可以看出RuSPS编码的蛋白等电点均<7,表明黑莓SPS蛋白主要以弱酸性形式存在(表2)。

表2 黑莓RuSPS基因基本信息
黑莓RuSPS基因编码蛋白都具有SPS蛋白的蛋白调控位点,即SPS蛋白与14-3-3 调控蛋白作用的磷酸化位点以及SPS响应渗透压的磷酸化修饰位点[16]。通过NCBI Blast 工具检索发现黑莓SPS的氨基酸序列与其他植物的氨基酸序列有着广泛的相似性,如RuSPS1与苜蓿(MedicagosativaAAK09427.2)一致性为76%,相似性为84%;RuSPS2与沙梨(PyruspyrifoliaBAG30918)一致性为68.7%,相似性为73.9%;RuSPS3与桃(PrunuspersicaEMJ26601.1)和月季(RosachinensisXP_024192491.1)一致性分别为91.3%和96.2%,相似性分别为96.4%和97.9%。
利用23个基因编码的氨基酸序列与3条RuSPSs氨基酸序列进行系统发生分析,显示其序列相关性,发现RuSPS1、RuSPS3 之间进化距离较近(图1a),与RuSPS2距离较远。可以看到RuSPS1与AtSPS1距离较近,RuSPS3与PpySPS距离较近,同属A亚族,RuSPS2和NtSPS B、AtSPS3属于B亚族(图1)。

Cm.甜瓜(Cucumis melo ABF47344.1);Sl.番茄(Solanum lycopersicum,SlSPS NP_001233920.2,SlSPS4 XP_004250761.1,SlSPSB AFD64638.1);Ib.番薯(Ipomoea batatas AAL34531.1);Ac.中华猕猴桃(Actinidia chinensis AAL86360.1);At.拟南芥(Arabidopsis thaliana,AtSPS1 NP_197528,AtSPS2 NP_196672,AtSPS3 NP_171984,AtSPS4 NP_192750);Hb.橡胶(Hevea brasiliensis XP_021661770.1);Mi:芒果(Mangifera indica BAM37540.1);Ms.苜蓿(Medicago sativa AAK09427.2);Ppy.沙梨(Pyrus pyrifolia BAG30918); Sb.高粱(Sorghum bicolor ACX94229);Vv.葡萄(Vitis vinifera AAW82754.1);Nt.烟草(Nicotiana tabacum NtSPSC ABA64520,NtSPSB ABA64521);Zm.玉米(Zea mays NP_001105694.1);Os.水稻(Oryza sativa BAA08304.1);Pt.毛果杨(Populus trichocarpa XP_002328899.1);Pp.桃(Prunus persica EMJ26601.1);Rc.月季(Rosa chinensis XP_024192491.1)。
2.2 黑莓SPS蛋白氨基酸序列分析
利用Meme软件在黑莓SPS蛋白中预测出10个保守元件。元件1~3、6和8~10在所有SPS蛋白中均被预测到;除RuSPS1外元件5在所有蛋白中均存在,RuSPS3中缺失元件7和元件4;元件10在C家族中未发现(图1b),RuSPS1和RuSPS3在保守元件中具有较大的特异性。根据 InterProScan 注释显示,元件 1~4 都是糖基转移酶蛋白,其余都是SPS 相关蛋白。
已有研究表明在植物中SPS 蛋白普遍存在3 个相对保守的磷酸化位点,分别为光/暗调控位点(Ser-158),14-3-3调控蛋白结合位点(Ser-229)和渗透压调控位点(Ser-424);同时具有2个保守的蛋白结构域,分别是位于C端的SPP-like 结构域(SPP-like domain)以及位于N 端的葡糖基转移结构域(glycosyltransferase domain)[17]。
对分别来自拟南芥、沙梨、烟草、高粱和黑莓的3个SPS亚家族共11个氨基酸序列进行了多重序列比对(图2),分析了它们的序列相似性、SPS蛋白结构域以及磷酸化位点。通过对不同家族成员SPS的比较可以看到,在所有SPS基因编码的氨基酸的N端,除RuSPS3前段缺失之外,其余前23个氨基酸残基几乎是保守的,但是它们的调控及结合位点序列区域存在较明显的差异。可以看到所有A亚族二分体核定位序列均表现为KK(11aa)KRR,而在B亚族和C亚族中表现为KK(12aa)RR。DTGGQVKY是6-磷酸果糖的共同识别位点,在所有的SPS中几乎都是保守的。14-3-3和尿苷二磷酸葡萄糖结合位点相互重叠,在A亚族成员包含14-3-3结合位点(HXRXXSXP, ser229),但B和C亚族成员缺失,与6-磷酸果糖结合位点一样,尿苷二磷酸葡萄糖结合位点在所有的SPS中都很保守,这反映了它们在功能上的一致性。A和C亚族都遵循BHXBXXS序列进行渗透调节,但在B亚族中丝氨酸则被天冬氨酸取代;同时3个RuSPS基因编码的SPS蛋白均具有保守的葡糖基转移结构域和SPP-like 结构域。

箭头以内序列表示二分体核定位序列。从上至下3处矩形框依次表示3个保守的磷酸化位点:6-磷酸果糖结合位点、14-3-3 调控蛋白结合位点(Ser-229)/尿苷二磷酸葡萄糖结合位点和渗透压调控位点(Ser-424)。SPP-like结构域和葡萄糖基转移结构域如实线所示。The sequence within the arrow represents the bipartitenuclear localization sequence; The three rectangular frames from top to bottom represent three conserved phosphorylation sites: fructose 6-phosphate binding site, 14-3-3 regulatory protein binding site(Ser-229)/UDP-Glc binding site and osmotic pressure regulatory site(Ser-424),the glucosyltransfer domain and the SPP-like domain were marked with black lines, respectively.
2.3 黑莓SPS蛋白结构预测及分析
通常,蛋白质分子的多肽链经过盘曲折叠后形成相对稳定的空间结构,从而形成独特的活性以及物理和化学性质,因此蛋白质二级结构的预测与空间结构分析对于了解其功能有重要的意义。蛋白质二级结构是指多肽链中主链原子以氢键构成的局部空间排布,常见的二级结构元件有α-螺旋、β-折叠、延伸链和无规则卷曲等。黑莓SPS蛋白的二级结构如SOPMA网站预测结果(图3)所示,可以看到α-螺旋及无规则卷曲是3个SPS蛋白最多的结构元件,而β-折叠及延伸链随机分布于整个蛋白质中。统计结果表明,RuSPS1蛋白由39.84%的α-螺旋、39.84%的无规则卷曲、13.97%的延伸链和6.34%的β-折叠组成,RuSPS2蛋白由40.54%的α-螺旋、39.61%的无规则卷曲、13.11%的延伸链和6.74%的β-折叠组成,RuSPS3蛋白由37.13%的α-螺旋、41.48%的无规则卷曲、15.16%的延伸链和6.23%的β-折叠组成。SWISS-MODEL在线预测结果表明,RuSPS蛋白质的三级结构主要由α-螺旋和无规则卷曲组成(图4),与二级结构预测结果一致。

蓝色blue.α-螺旋α-helix;绿色green.β-折叠β-fold;紫色purple.无规则卷曲irregular curl;红色red.延伸链extension chain。

不同颜色代表肽链的结构特征: 从N-端延(深蓝色)伸至C-端(红色)。Color shows the order of peptide chain, starting as dark blue from the N-terminus, going to red at the C-terminus.
2.4 黑莓RuSPS基因的表达分析
2.4.1 在不同组织中的表达
为了进一步了解RuSPS基因在黑莓组织器官中的表达特性,以黑莓RuActin基因为内参,对RuSPS1、RuSPS2和RuSPS3在‘Boysen’品种不同组织中的表达量进行比较分析。由结果(图5A)发现,3个基因在根、茎、叶、花、果实中表达量存在显著差异,其中RuSPS1在叶片和成熟果实中高度表达,RuSPS2在发育成熟的果实中有大量的表达,在其他器官中表达量较低,提示RuSPS2可能与黑莓果实发育有关,RuSPS3在各器官中的表达均较高,花和果实中的表达量稍高但与其他组织相比差异不显著。3个RuSPS基因在各种组织中有不同程度的表达,推测它们的功能是维持各种组织对蔗糖的基本需要。

不同小写字母表示在不同组织或不同发育阶段的表达量在5%水平差异显著。The lowercase letters indicate signifference at 0.05 levels in different tissues or at different stages.
2.4.2 在不同发育时期果实中的表达
‘宝森’品种果实在开花后9 d形成小核果,在21 d时果实开始膨大,果实完全成熟需历时39 d。对黑莓3个RuSPS基因在果实发育过程中即花后21~39 d中的表达量进行了比较分析(图5B),结果发现:RuSPS1表达量从果实发育早期逐渐上升,至中期27 d达到最高,而后表现为下降趋势;RuSPS2在果实发育早期和中期表达量变化不明显,在39 d时表现出较高的表达量,但与其他时期相比差异不显著;RuSPS3在果实发育过程中表达量呈线性增长,直至发育成熟时达到峰值。此外,可以看到除RuSPS1在27 d达到峰值,其余2个基因均在果实发育后期表达量较高,而在早期或中期表达量较低。
2.4.3 在叶片中的表达
对黑莓果实不同发育时期对应叶片中的RuSPS基因进行荧光定量分析发现,3个基因在果实发育进程各个时期对应的叶片中均有表达,且表达量在果实成熟期最大,分别较21 d时增加了59.7%、73.5%和59.4%(图5C)。RuSPS1在叶片中的表达量随着果实发育进程而减少,至果实成熟期突然上升;RuSPS2和RuSPS3在叶片中的表达量表现为花后27 d 前即果实发育早期和中期逐渐上升,在发育后期(花后33 d)有所下降,在果实完全成熟时表达量达到最高。
2.5 不同发育阶段黑莓果实及叶片中SPS酶活性的变化
在果实及叶片不同发育时期进行SPS酶活性的测定发现,果实及叶片中SPS活性与其RuSPSs基因的表达模式呈正相关。在果实中,SPS活性在花后21~27 d时显著上升,提高了197%,到花后33 d时略微下降,而后在花后39 d时达到最高值486.77 μg/(min·mg)。叶片中的SPS活性在花后21 d时低于果实,而后迅速上升;在花后33 d时迅速下降,最后在花后39 d时达到最高。SPS活性在叶片中的变化趋势与RuSPS2及RuSPS3表达模式一致。为了进一步解释黑莓SPS基因表达量与蔗糖磷酸合成酶不同发育阶段之间的关系,分别以叶片和果实4个发育阶段酶活力及该部位不同SPS的表达量的值进行了相关性分析,结果表明叶片中蔗糖磷酸合成酶活性与RuSPS2显著正相关(P<0.05,),而果实中蔗糖磷酸合成酶活性与RuSPS1显著负相关(P<0.05,表 3),提示叶片中SPS活性的变化一定程度上是由RuSPS2调控,而果实中SPS活性的变化则是由RuSPS1调控。
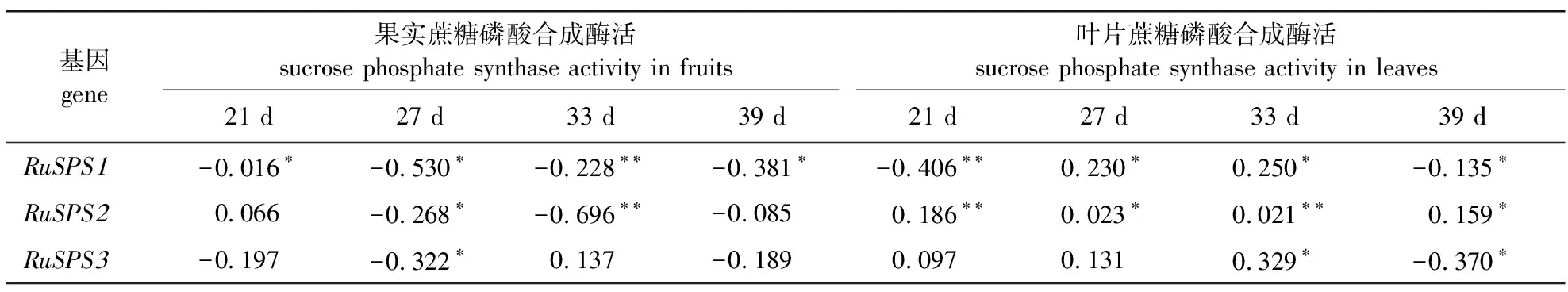
表3 不同发育时期黑莓果实及叶片中RuSPS表达量与SPS酶活性相关性分析
3 讨 论
本研究分析了黑莓SPS基因的系统发育关系、氨基酸序列、保守元件、不同组织器官和不同发育时期的表达差异。从黑莓品种‘宝森’中克隆分离了3个SPS基因家族成员,分别命名为RuSPS1(MT682348)、RuSPS2(MT682349)和RuSPS3(MT682350)。氨基酸聚类分析表明,RuSPS1和RuSPS3属于典型的A亚族,RuSPS2属于B家族,这与大多数植物中含有3~5个SPS基因的研究结果一致[18],尚未发现属于其他亚族的成员。目前认为,SPS活性受到转录水平和蛋白磷酸化共同调控。RuSPS1与AtSPS1中的Ser-424位点发生相同位点氨基酸突变,与聚类分析结果一致,表明其功能高度类似。此外,14-3-3 蛋白的磷酸化也参与了SPS活性调控[19],其通过与Ser-229 蛋白磷酸化位点互作抑制其自身表达,从而促进SPS活性升高和蔗糖积累[20]。氨基酸序列比对显示黑莓SPS蛋白的Ser-424、Ser-158、14-3-3蛋白磷酸化位点相对保守,RuSPS1前6个氨基酸突变,以及RuSPSs在结构域和磷酸化位点上的非保守替换并且由于点突变的累计,缺失和插入主要局限于N端和C端。这些变化都可能影响它们的结构和功能,从而改变它们的活性。本研究还预测了黑莓SPS蛋白的空间结构,所有的这些序列特征和空间特征可能会影响黑莓SPS蛋白的整体催化活性和功能专一性。
在本研究中,黑莓不同亚族SPS基因表达显示了不同的时间和空间特异性。B亚族RuSPS2基因在成熟叶片中表达量较高,表明其可能参与了源器官中蔗糖的合成,这与之前在水稻中OsSPS1的研究结果相似[21],但在甘蔗中则得到了相反的结果,SofSPSB在成熟叶片中表达量甚微[22]。当黑莓果实发育39 d时,RuSPS2在果实中表达量是叶片中的2.46倍,说明蔗糖合成在果实中更密集。B亚族SPS基因在叶片及成熟果实中高表达,暗示着它们在‘源’‘库’关系、器官光合产物的积累方面可能起着重要作用,但B亚族SPS在更多植物中的表达模式还有待进一步研究。A亚族SPS基因被认为是植物重要的管家基因[23],常常在植物的各个组织器官中均有较高的表达。黑莓A亚族SPS基因RuSPS1在被检测的各个组织中均有较高的表达,仅在花中表达量较低,推测它的功能是维持各种组织对蔗糖的基本需要。黑莓中A亚族及B亚族SPS基因表达量在果实及叶片发育过程中都随着果实成熟度增加而上调,与黑莓果实及叶片中SPS活性的变化趋势一致,这些结果表明黑莓SPS基因家族在黑莓果实发育过程中发挥重要作用,并影响植物的蔗糖的合成和积累,但它们的表达与黑莓‘源’‘库’器官间的联系仍需更深入的研究。
