‘京红’桃芽变缝合线局部早熟分子机制的探究
2022-11-26刘璐琪申艳红宋立琴石天磊武军凯张立彬
刘璐琪,申艳红,宋立琴,石天磊,武军凯,张立彬
(河北科技师范学院 园艺科技学院,河北省特色园艺种质挖掘与创新利用重点实验室,河北秦皇岛 066004)
桃果实是典型的呼吸跃变型果实,采后迅速软化、腐烂变质,特别是有些桃品种果实缝合线局部早熟,主要表现是缝合线局部凸起、软化、变色,当整个果实达到适采成熟度时,因该部位过软而无法运输销售。缝合线软化是当今桃生产中存在的重要问题之一,有些品种发病率超过50%,严重影响品质和商品价值[1-2]。有研究发现,缝合线软化部位的木质部和韧皮部面积相对较大,维管束发达粗大,糖代谢和相关酶活性都较旺盛[3-4]。桃缝合线部位的生长素(IAA)和玉米素(ZT)含量与缝合线局部软化具有相关性[5]。学者们对导致缝合线软化的原因进行了分析,有学者认为属于生理性障碍,与缺钙有关[6]。刘志民等[2]认为,与品种关系较大,‘大久保’、‘燕红’等品种发病率较高,种仁充水、呼吸加强使其提早释放乙烯,是缝合线软化的主要原因。目前,关于桃缝合线早熟的相关研究大都局限于生理生化水平,相关分子机理研究还鲜见报道。
本课题组在‘京红’桃中获得了一个芽变。该芽变具有两个特性,一是晚熟,突变体比野生型的成熟期约晚2周;二是芽变缝合线部位具有局部早熟特性,缝合线部位比果面部位提前两周变红,是研究缝合线局部早熟的好材料。基因差异表达是调节桃果实不同部位成熟进程差异的根本原因,通过转录组测序技术挖掘重要功能基因已经被广泛应用。本研究以‘京红’野生型果实为对照,采用转录组测序技术对芽变果实的缝合线和果面部位差异表达基因进行比较,挖掘桃果实缝合线提前成熟相关基因,分析其表达规律,从分子水平上分析果实不同部位成熟调控分子机制。
1 材料和方法
1.1 试验材料及取样
供试材料为桃品种‘京红’[Prunuspersica(L.) Batsch. cv. Jinghong wild type,JHW]及其芽变(Jinghong mutant,JHM)的果实,采自河北省秦皇岛市抚宁区桃园。在桃花后40、60、66、73、82和90 d进行果实采样,当天运回实验室进行处理。每份样品选择果实大小一致、无病虫害、无损伤的9个果实进行硬度测定。然后,将9个果实分为3组,将每组3个果实削去果皮后,将缝合线部位和果面部位的果肉分别切成小块后混匀,样品分别编号为WSP(JHW缝合线果肉)、WP(JHW果面果肉)、MSP(JHM缝合线果肉)、MP(JHM果面果肉),后续用于花青素含量的测定以及转录组测序。
1.2 果实硬度的测定
桃果实硬度采用TA.XTC-18质构分析仪(上海保圣实业发展有限公司)进行测量,选用TA/2N型号探头,TPA模式,设定触发点30 gf,目标量30 gf,测试速度2 mm/s,得出测定参数。随机选取9个果实测定缝合线和果面处硬度,用探头在削去果皮后的果肉处进行测定,每个部位测定3次。
1.3 果肉花青素含量的测定
果肉花青素含量的测定参照杨夫臣等[7]的方法。取1.0 g果肉样品,液氮研磨成粉末后,置50 mL离心管中,加0.1 mol·L-1HCl溶液15 mL,置32 ℃温箱中浸提8 h,期间摇动数次,离心,取上清液装入光径为1 cm的比色杯中,于530 nm下比色,记录吸光度值(OD)。3次重复。
1.4 转录组测序及差异表达基因筛选
将花后66 d的4个果实样品(WSP、WP、MSP、MP)送到广州基迪奥生物科技有限公司,使用第二代高通量Illumina HiSeqTM2500测序技术进行转录组测序,对得到的数据进行过滤得到clean data后,将reads比对到参考基因组(基因组版本:GCF_000346465.2)上,并利用Cufflinks组装转录本,得到已知转录本与新转录本。基因表达量的计算使用FPKM(fragments per kilobase of transcript per million fragments mapped reads)法,按照edgeR的一般过滤标准[log2|fold change(FC)| >1 & FDR (false discovery rate) <0.05]筛选差异表达基因,将筛选得到的差异表达基因做GO(gene nntology)和KEGG(kyoto encyclopedia of genes and genomes)富集分析。
1.5 差异表达基因的实时荧光定量表达分析
以北京博迈德生物技术有限公司的植物RNA提取试剂盒分别提取‘京红’及其晚熟芽变果面及缝合线处的RNA,逆转录为cDNA。参照候选基因序列设计荧光定量引物(表1),以桃PpTEF2为内参,20 μL荧光定量PCR反应体系为:2.0 μL cDNA,0.5 μL上游引物,0.5 μL下游引物,10 μL SYBR Premix Ex TaqTM(TaKaRa,日本),7.0 μL灭菌水。反应程序为:95 ℃退火30 s,95 ℃ 5 s,60 ℃ 15 s,39次循环。使用购自美国伯乐有限公司的荧光定量PCR仪(Bio-Rad CF CONNECTTMReal-Time System)检测基因的表达量。2-ΔΔCT法[8]分析基因表达的相对变化。进行3次生物学重复测定,采用Excel软件作图。

表1 荧光定量引物序列
2 结果与分析
2.1 桃果实形态、硬度及花青素含量随发育期变化特征
‘京红’野生型(JHW)及其芽变(JHM)不同发育时期的果实形态比较可以看出,芽变果实具有晚熟特性,比野生型果实晚成熟约2周;除此之外,芽变果实缝合线部位比果面部位局部早成熟。在花后66 d,野生型果实约为七成熟,此时果皮已转白,缝合线部位和部分果面均为红色;而花后66 d的突变体果实,果面仍为绿色,缝合线部位为红色,此时其果面与缝合线部位成熟度差异较大,用于后续转录组测序(图1,A)。随着果实的生长发育,JHW和JHM果肉硬度均逐渐降低,尤其在花后66 d开始明显下降;同时,两种材料果面硬度均大于相应时期缝合线部位的硬度,芽变果面和缝合线硬度基本大于同期的野生型(图1,B)。另外,如图1,C所示,随着桃果实的逐渐成熟,野生型和突变体果实果面以及缝合线部位的花青素含量均呈上升趋势;在花后66 d开始明显上升,且野生型始终高于突变体,突变体中又表现为缝合线始终高于果面。

WP. 野生型果面;WSP. 野生型缝合线;MP. 突变型果面;MSP. 突变型缝合线;下同
2.2 转录组数据质量分析与基因注释
将花后66 d的‘京红’野生型及其芽变果实的果面和缝合线4个样品(WP、WSP、MP、MSP)进行转录组测序,转录组测序数据组装和注释情况见表2。其中,去除接头、Ploy A、低质量序列以及核糖体RNA后,4个样品WP、WSP、MP、MSP的干净读长(Clean Reads)分别为56 527 760、50 253 068、54 834 138、54 811 850,GC含量分布在45.79%~46.06%之间,并且Q20均大于98%,表明测序数据质量优良,可以进行后续试验。对所有样品检测基因数目进行统计,共检测到20 108个基因,占桃参考基因组基因总数(23 128)的86.94%。

表2 测序数据组装和注释统计表
2.3 差异基因数目统计与Pathway功能显著性富集分析
以FDR<0.05且|log2FC|>1为条件筛选差异基因。如图2所示,WSP相对于WP、MSP相对于MP的差异表达基因分别为1 297和1 889个;其中,WSP相对于WP的差异表达基因中561个基因上调表达,736个基因下调表达;MSP相对于MP的差异表达基因中1 308个基因上调,581个基因下调。由此可见,突变体缝合线与果面之间差异表达基因数目远多于野生型,尤其是上调表达基因数量大大增加,这些基因在缝合线与果面之间的差异表达促进了缝合线部位的提早成熟。另外,对突变体缝合线与果面的1 889个差异基因进行分析发现,其共参与112种代谢通路,其中Top 20的代谢通路见图3,主要富集在代谢途径(metabolic pathways)、次生代谢产物的生物合成(biosynthesis of secondary metabolites)、植物激素信号转导(plant hormone signal transduction)、苯丙素的生物合成(phenylpropanoid biosynthesis)等代谢通路。因此,‘京红’桃突变体果实缝合线比果面提早软化和变红,与这些代谢途径、次生代谢产物的生物合成、植物激素信号转导、苯丙素的生物合成有关。

图2 差异基因数目统计

图3 MSP与MP之间KEGG富集前20代谢途径气泡图
2.4 桃缝合线部位提前成熟相关候选基因的筛选与分析
比较突变体桃果面与缝合线部位间差异基因的表达,筛选出24个缝合线提前成熟相关的候选基因,包含5个细胞壁降解相关基因、9个色素合成相关基因、5个乙烯合成与信号转导相关基因、3个生长素应答基因、2个NAC转录因子(表3)。

表3 桃果实缝合线和果面的差异表达基因
首先,细胞壁降解相关基因中包括4个果胶降解酶基因和1个纤维素降解酶基因,分别为果胶酯酶(pectinesterase domain-containing protein,PME)、果胶裂解酶(pectate lyase 5,PL5)、内聚半乳糖醛酸酶(endo-polygalacturonase,PG)、β-半乳糖苷酶(beta-galactosidase protein,BGAL)和木葡聚糖内转糖基酶/水解酶(xyloglucan endotransglucosylase/hydrolase protein 33,XTH33)基因,它们在突变体缝合线部位的表达量显著高于果面中的表达量。
其次,色素合成相关基因包含2个卟啉叶绿素合成途径基因,分别为血红素加氧酶(heme oxygenase 1,HO1)和原叶绿素酸酯氧化还原酶(protochlorophyllide reductase,PORA)基因;5个花青素合成途径基因,分别为查尔酮合酶(chalcone synthase,CHS)、查尔酮异构酶(chalcone flavonone isomerase,CHI)、黄烷酮3-羟化酶(flavanone 3-hydroxylase,F3H)、二氢黄酮还原酶(dihydroflavonol 4-reductase,DFR)、原花色素双加氧酶(leucoanthocyanidin dioxygenase,ANS)基因;2个花青素合成调节转录因子,分别为MYB转录因子(R2R3 MYB transcription factor 10,MYB10)、bHLH转录因子(transcription factor bHLH111-like isoform X1,BHLH)。叶绿素合成途径关键基因PORA在突变体缝合线部位表达量低于果面部位,而5个花青素合成途径基因与2个转录因子的表达趋势则与PORA相反,在缝合线部位表达量显著高于果面处,这与缝合线部位红色、果面部位绿色密切相关。
再次,乙烯在桃果实成熟的过程中起关键作用,共筛选出5个乙烯合成与信号转导相关基因。2个乙烯合成关键基因分别为ACC合成酶(1-aminocyclopropane-1-carboxylate synthase,ACS1)、ACC氧化酶(1-aminocyclopropane-1-carboxylate oxidase,ACO1)基因,在突变体缝合线部位表达量是果面部位的数倍,说明缝合线部位乙烯合成速率显著高于果面;3个乙烯转导相关基因分别为1个乙烯受体(ethylene receptor 2,ETR2)、2个乙烯应答因子(ethylene-responsive transcription factor,ERF),它们在突变体缝合线部位和果面部位的表达差异也达到显著水平。
另外,有研究表明,生长素可能是桃果实成熟期乙烯释放的先决因素,筛选的3个Aux/IAA家族基因在缝合线部位表达量显著高于果面部位。此外,还筛选了2个参与果实成熟调控的NAC转录因子——NAC2和NAC72,二者具有相似的表达趋势,在缝合线部位的表达量均低于果面部位,NAC2在缝合线部位的表达量约为果面部位的1/2。
进一步对差异表达基因进行了热图分析(图4)可知, 24个差异表达基因的表达模式聚类为三大组。一组是PORA、NAC72、NAC2和ERF105,这4个基因在野生型缝合线和果面的表达量相当,而在突变体缝合线部位的表达量显著高于果面;一组是花青素合成相关基因BHLH、MYB10、ANS、CHI、CHS、F3H、DFR,以及细胞壁降解基因BGAL、XTH33,这一大组基因在野生型桃果实的缝合线和果面表达量较低且无显著差异,而在突变体缝合线部位大量表达,且表达量显著高于果面;另一组是剩余的11个基因,它们在野生型桃果实中的表达量较高,在突变体缝合线与果面中差异表达。
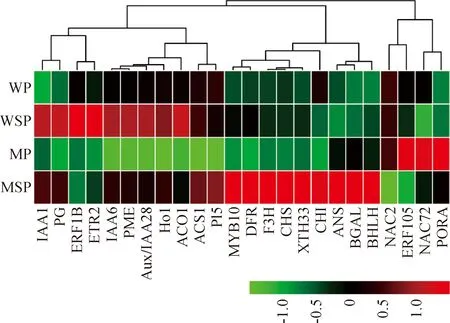
图4 差异表达基因的热图分析
2.5 转录组数据验证
对12个差异表达候选基因进行了荧光定量验证,结果如图5所示,对比荧光定量与RNA-seq两种技术获得的差异基因表达趋势,二者基本一致,这进一步证明了转录组数据的可靠性。
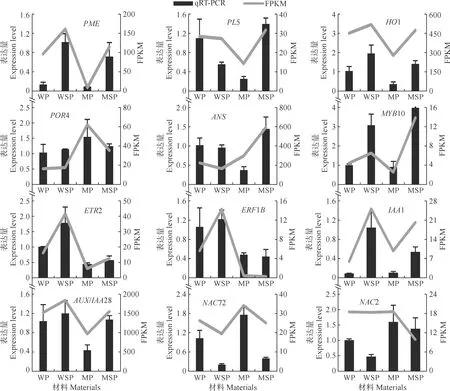
图5 用荧光定量PCR分析12个差异表达基因的相对表达量
3 讨 论
纤维素和果胶的降解是导致果实软化的直接原因。刘志民等[2]发现,缝合线易软桃品种在果实成熟初期缝合线部位的果胶酶活性较高,说明果胶降解在缝合线软化中起作用。通过比较‘京红’突变体缝合线与果面部位基因的差异表达,筛选了5个细胞壁降解相关基因,其中PME、PL5、PG和BGAL参与果胶降解,在缝合线部位的表达量显著高于果面数倍,尤其是PG基因在突变体缝合线的_fpkm值高达360,是突变体果面_fpkm(44)的8倍,说明缝合线提前软化与PG基因的高表达密切相关,PG在多种果实软化中的重要作用已经被证实[9-10]。另外,纤维素降解酶基因XTH33在缝合线中的表达量也显著高于果面,超过了10倍,说明在缝合线果胶降解的同时也伴随着纤维素降解。
桃果实颜色与叶绿素、类胡萝卜素和花青素含量相关[11-12]。在桃果实成熟过程中,绿色逐渐褪去,红色逐渐出现,说明桃果实转色伴随着花青素含量增加和叶绿素的减少。本研究筛选出了9个色素合成相关基因。其中,HO1和PORA为卟啉与叶绿素代谢途径基因,随着果实成熟叶绿素合成关键基因PORA表达量降低,减少了叶绿素合成;同时,CHS、CHI、F3H、DFR、ANS为花青素合成途径基因,在突变体缝合线处表达量为果面处的2倍以上,其中CHS与DFR基因的MSP/MP_fpkm倍数最高,分别为17和10,说明CHS和DFR基因的高表达促进了桃缝合线部位提早变红,这与Tsuda等[13]的CHS和DFR为花青素合成关键基因的结果一致。另外,前人已经证实,R2R3-MYB和bHLH类转录因子在花青素合成中具有重要调控作用[14-15]。在本研究中,MYB10与bHLH在‘京红’野生型缝合线与果面部位的表达量相当,而在突变体的两部位间表达量差异显著,变化趋势与花青素合成途径基因一致,推测它们是桃花青素合成调控的重要转录因子。
桃是呼吸跃变型果实,乙烯含量增加会促进果实迅速成熟衰老[16-17]。在桃缝合线易软品种中,缝合线处的乙烯含量明显高于果面处[18-19]。ACC合成酶(ACS1)与ACC氧化酶(ACO1)是乙烯合成过程中的限速酶,本研究中ACS1与ACO1在突变体缝合线部位的表达量是果面部位的3倍,显著提高了缝合线部位的乙烯合成速率;同时还检测到差异表达的乙烯受体基因ETR2,以及2个乙烯应答因子ERF1B和ERF105。研究表明,有些ERF参与了果实成熟过程,甜橙CitERF6通过上调叶绿素降解基因CitPPH的表达促进果实褪绿;桃PpeERF2、番木瓜CpERF9通过抑制相关细胞壁降解酶基因的表达,推迟果实软化[20-21]。本研究筛选的ETR2与ERF基因在桃突变体缝合线成熟过程中的具体作用还需进一步研究。除了以上基因,有些生长素相关基因也通过调控乙烯释放量参与果实成熟调控。在桃果实成熟过程中,ACS1与生长素合成限速酶基因YUCC11的表达趋势相同[22];生长素原初响应基因(Aux/IAA)与ACS1、YUCC11基因共表达,可能参与了桃果实成熟调控[23]。本研究中的3个Aux/IAA家族基因在突变体缝合线部位的表达量高于果面,与ACS1和ACO1表达趋势一致,这与前人研究结果相符。
此外,桃缝合线部位的早熟也与其特殊的结构相关。桃果实为真果,由单心皮雌蕊发育而成,雌蕊由叶子演化而来,缝合线相当于叶子的叶缘位置,发育为种子的胚珠着生在缝合线上。在缝合线易软桃品种中缝合线部位维管束粗大发达,韧皮部和木质部面积也相对较大,腺腔发达[3]。
综上所述,‘京红’桃芽变缝合线部位比果面部位局部早成熟约2周,在花后66 d,缝合线与果面部位差异表达基因1 889个,从中筛选到24个缝合线早熟相关基因,包含5个细胞壁降解相关基因,9个色素合成、调控相关基因,5个乙烯合成与转导相关基因,3个生长素应答基因和2个NAC转录因子。通过分析这些基因的差异表达推测缝合线部位局部早熟的原因。桃种仁产生的乙烯从缝合线向周围扩散,促进ACS1和ACO1等基因的转录,合成了较多乙烯,乙烯又进一步调控该部位PG、XTH33、CHS、DFR等细胞壁降解与色素合成相关基因的表达,导致该部位的提前成熟。
