蔬菜作物逆境生理与抗逆栽培研究进展
2022-11-21束胜胡晓辉王玉张润花袁颖辉陈长军施露郭世荣
束胜,胡晓辉,王玉,张润花,袁颖辉,陈长军,施露,郭世荣
(南京农业大学园艺学院,江苏 南京210095)
2020年我国蔬菜播种面积约2 200万hm2,其中设施蔬菜面积约410万hm2[1],蔬菜已成为我国第一大经济作物。然而,蔬菜生产中常常面临高温、低温、干旱、高湿、涝害、盐碱、弱光、病虫草害、连作障碍等不利环境条件,蔬菜作物的生长发育受阻,导致产量和品质下降。因此,急需解析蔬菜作物逆境生理和逆境调控机制,培育蔬菜作物抗逆品种,开发提高蔬菜植株抗性的技术和产品,建立完善的蔬菜可持续健康生产技术体系。本文首先阐述了逆境胁迫对蔬菜作物的影响以及蔬菜作物的抗逆生理,然后对抗逆栽培的主要品种、技术和产品等进行综述,最后对蔬菜抗逆栽培进行总结和展望,以期为蔬菜绿色、优质、高效、高产抗逆栽培提供理论依据和实践指导。
1 蔬菜逆境障碍因子及其危害
1.1 生物逆境因子
1.1.1 病原菌蔬菜病原菌主要包括真菌、卵菌、细菌和病毒,它们分别与寄主植物和环境因子互作,进而致病。
真菌病害占蔬菜病害的80%以上。灰葡萄孢菌(Botrytiscinerea)是引起园艺作物最重要的病害,为灰霉病的病原菌。研究发现黄瓜转录因子CsWRKY10通过降低活性氧含量和抑制茉莉酸(JA)介导的抗性信号通路,对黄瓜灰霉病菌的抗性起负调节作用,但增强了黄瓜对褐飞虱的抗性[2]。研究表明,叶面喷施人工合成的双链小RNA(dsRNA)能够抑制灰葡萄孢菌、菌核病菌(Sclerotiniasclerotiorum)、立枯丝核菌(Rhizoctoniasolani)和黑曲霉(Aspergillusniger)的致病性,但不能抑制胶孢炭疽病菌(Colletotrichumgloeosporioides)和致病疫霉菌(Phytophthorainfestans);用dsRNA处理土壤,能够抑制大丽轮枝菌(Verticilliumdahliae)的侵染;dsRNA防效取决于植株对其吸收率[3]。
卵菌主要包括疫霉和腐霉。疫霉菌引起的作物疫病曾被称为“植物瘟疫”,有160多种,能侵染蔬菜等上千种植物,具有发病快、变异快、流行快等特点,在生产上防控比较困难。疫霉菌侵染宿主过程中,质外体扮演着极其复杂和精确的“攻击防御”角色,病原菌攻击寄主采用“诱饵模式Decoy”[4],即疫霉菌分泌的糖基水解酶(XEG1),通过攻击寄主细胞壁,破坏植物的抗病性,但植物通过细胞膜免疫受体RXEG1来识别XEG1,是植物启动对疫霉菌免疫的分子基础;疫霉菌会利用XEG1的酶活性丧失突变体XLP1作为“分子诱饵”,来掩护XEG1对植物的攻击。进一步研究表明,疫霉菌通过N-糖基化修饰保护XEG1,从而逃避寄主植物大豆分泌的天冬氨酸蛋白酶(GmAP5)的识别以防护罩“Shield”形式保护XEG1免于被植物降解[5];疫霉菌分泌型分子诱饵XLP1则通过C端缺失逃避GmAP5的识别,与XEG1竞争性结合抑制子GmGIP1,从而掩护XEG1攻击植物免疫系统。大豆GmAP5是一类新的广谱抗病基因资源,对XEG1的深入认识,可为以分泌型核心毒性因子为分子靶标筛选杀菌剂提供理论依据。
由细菌丁香假单胞菌(Pseudomonassyringae)引起的黄瓜角斑病是设施栽培黄瓜的主要细菌性病害。研究表明从发病株形成的粒径1.1~4.7 μm气溶胶占整个气溶胶的72.16%,空气中84~179 CFU·m-3气溶胶是发病的阈值,为及时清除棚内病株提供理论支撑[6]。番茄细菌性叶斑病菌miR863-3p能够作用于植物免疫中的正向和反向调节因子,进而调控植物免疫反应,即侵染初期在miRNA水平上抑制非典型受体样假激酶基因(ARLPK1和ARLKP2)的表达;侵染后期,在蛋白水平上抑制对miRNA加工和合成关键蛋白的表达,从而关闭植物防卫反应,这有助于通过RNA沉默增强植物抗病性[7]。
负链RNA病毒是对蔬菜等作物生产具有严重威胁的植物病毒,例如番茄斑萎病毒,它能侵染1 000多种植物,对全球经济和园艺作物安全生产构成重大威胁。已成功建立植物多分体负链RNA病毒的反向遗传学体系,可用于病毒侵染和致病机制的深入研究,意味着植物多分体负链RNA病毒研究新时代的到来[8]。
1.1.2 害虫烟粉虱是被联合国粮农组织(FAO)认定的迄今唯一的“超级害虫”,在设施茄果类蔬菜生产中危害严重。Xia等[9]研究表明植物和动物之间存在功能性基因水平转移现象,并揭示了昆虫如何利用水平转移基因来克服宿主的防御,为探索昆虫适应性进化规律开辟了新的视角,也为新一代靶标基因导向的烟粉虱田间精准绿色防控技术研发提供全新思路。研究表明,烟粉虱磷脂酰乙醇胺结合蛋白(PEBP4)是导致番茄感染黄化曲叶病毒的关键因子,通过维持其细胞凋亡与自噬作用相互拮抗的中度免疫平衡,以达到高效传毒给番茄植株[10]。
小菜蛾是十字花科蔬菜作物的重要害虫,是最早被报道对Bt生物杀虫剂产生抗药性的农业害虫。小菜蛾昆虫激素含量升高及其激活的MAPK信号途径反式调控中肠受体和非受体同源基因的差异表达,从而使小菜蛾在维持正常生长发育的前提下对Bt Cry1Ac杀虫蛋白产生高抗性;小菜蛾MAPK信号途径调控的转录因子GATAd通过与Bt Cry1Ac杀虫蛋白受体基因(PxmALP)的启动子不同靶标位点结合,调控抗性小菜蛾PxmALP表达下调,从而参与小菜蛾Bt Cry1Ac杀虫蛋白抗性的形成[11]。
1.1.3 草害蔬菜地杂草种类繁多,危害严重的有40余种。稗草是世界性恶性杂草,被列为我国农田危害严重的杂草之首。在设施蔬菜栽培中,危害较严重的主要有马塘、狗尾草、刺苋和荠菜。草甘膦是一种灭生性除草剂,由于长期和大量施用已导致田间至少48种杂草对草甘膦产生了抗药性。五氟磺草胺自投入市场以来,在防治稗草方面起到了重要作用,但随着使用年限的增加,我国常见稗草已对五氟磺草胺产生抗药性,并呈现不断扩大的趋势,现已无法控制。研究发现,稗草中抗五氟磺草胺的主效基因为水稻细胞色素P450基因(CYP81A68),而CYP81A68启动子的甲基化是导致CYP81A68高表达的主要原因,研究结果为稗草抗除草剂及其转录调控机制提供新视野[12]。
1.2 非生物逆境因子
1.2.1 极端温度极端温度会导致蔬菜细胞膜透性增大,活性氧(ROS)大量生成,光合机构受损。高温会导致酶变性失活、蒸腾作用加剧、叶片失水萎蔫、花序脱落、果实灼伤等症状;而低温抑制呼吸代谢,根系吸水缓慢,叶片失绿、变黄,果实出现坏死斑。蔬菜在生殖生长阶段对温度尤为敏感,高温和低温均可显著影响番茄的开花数和坐果率,30 ℃就会使番茄花粉萌发率降低50%以上[13-14]。
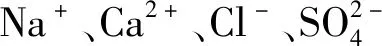
1.2.3 酸、碱胁迫酸性物质在叶片表面沉积直接引起叶片枯黄甚至坏死,落入地表导致土壤酸化,间接阻碍作物生长发育。酸、碱胁迫除了对蔬菜造成渗透胁迫、离子毒害,还会引起pH胁迫。酸、碱胁迫下蔬菜细胞内K+/Na+稳态被破坏,丧失细胞膜完整性。钟嘉文等[15]研究表明,当pH值为3.0时,生菜净光合速率降低17.0%,产量显著下降。此外,土壤pH值决定了营养元素是否处于可供蔬菜植株吸收利用的状态,因此,酸、碱胁迫容易引发蔬菜缺素症。
1.2.4 水分胁迫干旱胁迫下蔬菜由于吸水量小于蒸腾失水量,表现为叶片萎蔫、茎端下垂,细胞伸长,生长受到抑制。干旱胁迫下气孔开度降低以阻止水分散失,CO2吸收随之受限,且叶绿素降解加速,合成减慢,光合电子传递受阻,光合作用减弱。干旱胁迫还导致植物体内DNA、RNA和蛋白质等迅速分解,泛素化蛋白水平上升。
涝害对蔬菜的伤害主要是根际缺氧造成,缺氧阻碍了根系有氧呼吸,抑制根系生长,且无氧呼吸易积累乙醇、乳酸等有害物质,使得根尖发黑,根系腐烂;缺氧环境下蔬菜茎基部可产生多个不定根,光合色素含量迅速下降,叶片转黄、早衰甚至脱落。
设施栽培条件下,高湿环境极易引发瓜类霜霉病以及茄果类炭疽病、疫病和灰霉病等真菌性病害发生,导致产量损失40%~70%。高湿有利于缩短其孢子产生时间或提高其孢子萌发率,例如在相对湿度(RH)95%以上时,黄瓜、番茄叶斑病的棒孢菌孢子萌发率可达92.6%[16]。高温、高湿条件下,蔬菜叶片气孔开度减小,光合速率降低,生长速度减慢,抗氧化酶活性降低,同时显著抑制了生殖生长。此外,高湿环境下,植物蒸腾作用受到抑制,养分吸收受阻,易发生生理性病害。
1.2.5 光照强度胁迫在冬春季蔬菜栽培,尤其是设施栽培中弱光寡照常伴随低温胁迫,大气污染或设施覆盖材料透光性差等也会带来弱光逆境。弱光环境下,蔬菜净光合速率和光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)显著降低,同化产物合成量减少。研究发现,番茄在弱光处理5 d后叶绿素a和叶绿素b与对照相比分别降低12.28%和10.96%[17]。蔬菜对弱光会做出避荫响应,如叶片变薄、变大,节间距增大,茎秆和叶柄变细、变长,幼苗下胚轴增长,这些都有利于光截留最大化。此外,长期弱光导致同化产物分配改变或入库受阻,引起花芽分化、果实发育不良,最终影响果实产量和品质。相反,夏季蔬菜栽培可能遭受强光胁迫。强光会减少叶绿素含量,导致气孔限制增加,同时抑制PSⅡ的修复,并引起光氧化、D1蛋白周转、叶黄素循环和电子传递受阻,Rubisco酶活性降低,引发光抑制。强光下番茄α-生育酚和质体醌水平显著提高,以减缓强光对番茄造成的光抑制和光漂白[18]。
1.2.6 重金属胁迫重金属离子被大量吸收后会严重干扰蔬菜细胞内原有离子稳态,致使代谢紊乱。重金属可与蛋白质、核酸互作而破坏其结构或功能,蔬菜植株的光合、呼吸、蒸腾作用以及矿质营养代谢等均会受到抑制,蔬菜生长发育迟滞,甚至死亡。重金属也会抑制抗氧化系统,诱导ROS积累,加剧氧化胁迫。铜胁迫可诱导泛素化蛋白酶体失活,蔬菜无法清除体内的错误折叠或变性蛋白,使植株生育受阻[19]。
1.3 连作障碍
由于设施蔬菜栽培的半封闭性,加之复种指数高、作物种类单一和不合理的肥水管理,使得连作障碍频发。连作障碍造成植株生长发育不良,产量、品质下降,极端情况下,局部死苗,不发苗或发苗不旺;多数受害植物根系发生褐变、分支减少,活力降低,分布范围狭小,导致水分、养分的吸收能力下降。连作障碍在植物科属间存在显著的差异,茄科、豆科、十字花科、葫芦科和蔷薇科等蔬菜作物易发生连作障碍。种植5年以上的大棚、日光温室连作障碍达到80%,20年以上的连作障碍接近100%,严重制约设施蔬菜的可持续发展[20]。连作障碍形成的原因及危害主要有以下几个方面:
1.3.1 土壤理化性状恶化日光温室内连续10年种植甜瓜,土壤含盐量会增加数倍。过量施用化肥和有机肥,特别是生理酸性肥料和未完全腐熟的有机肥,破坏了土壤的缓冲性和离子平衡,最终导致土壤酸化。张福建等[20]研究发现,江西地区辣椒连作2年内的土壤pH值平均为5.85,而连作2年以上的pH值平均为4.95。番茄连茬种植后土壤pH值下降,硝态氮含量升高,根系淀积物增多,影响番茄的生长发育。随着土壤酸化加剧,pH值下降,会将K+、Ca2+、Mg2+从土壤胶体中置换出来,这些离子极易被淋溶,导致作物出现缺钙、缺镁的症状;而Mn2+和Al3+等离子在土壤中的含量增加,植株受到毒害。
1.3.2 有害微生物积累同一种作物长期连续种植导致土壤某些特定微生物富集,病原菌数量增加,而有益细菌种类和数量减少。土壤酸化会使土壤微生物的活动和繁殖受到限制,土壤微生物活性降低,影响有机质的分解和养分循环。土壤中的细菌、放线菌会随着番茄的连作而数量降低,尖孢镰刀菌等真菌数量增加;细菌和真菌比值、放线菌和真菌比值、微生物群落多样性降低。番茄连作使土壤中微生物群落组成改变,放线菌门和球囊菌门的相对丰度降低,而枝鼻菌属相对丰度升高。黄瓜连作根际土壤微生物区系发生明显改变,土壤微生物群落的多样性指数及丰富度也随着种植年限的增加而降低。番茄、甜椒、黄瓜等蔬菜作物连续种植3年后,每年将会减产10%~20%,同时猝倒病、枯萎病、根结线虫等病虫害逐年加重[21]。
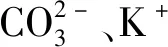
连作障碍显然属于土壤理化性状恶化、自毒物质危害的非生物和有害微生物侵染的生物混合胁迫作用的结果,产生原因错综复杂,需要深入研究。
2 蔬菜逆境适应性与抗逆栽培机制
2.1 形态结构适应性变化
非生物胁迫下植株形态结构和生理生化代谢会发生一系列的变化,以适应逆境胁迫。水涝胁迫下,耐涝性冬瓜于48 h后即可萌发不定根,且随水涝胁迫时间延长,不定根数量和长度明显增加,而涝敏感性品种不定根数量形成较少[23]。淹水胁迫会增加大豆下胚轴通气组织数量和面积,施用生长调节剂S3307进一步提高大豆对淹水逆境的适应性,增加通气组织数量和通气组织面积,以应对淹水胁迫对植株造成的缺氧胁迫[24]。而干旱胁迫下,由于气孔周围保卫细胞的膨胀压力而使气孔发生运动,并出现气孔开度降低,以减少水分的蒸发,提高蔬菜作物对干旱胁迫的适应性。Wang 等[25]研究发现大豆中受光诱导的TGACG基序结合因子(GmSTF3/4)和开花基因(GmFT)会从大豆的地上部移动到根系中,相互依赖地诱导根瘤器官发生。弱光胁迫下番茄幼苗叶片形态产生适应性变化,叶面积、茎叶夹角、茎叶垂角、垂度均显著提高,以响应弱光胁迫。
2.2 抗逆栽培机制
2.2.1 植物生长调节剂目前已报道的可提高植物抗逆性的外源物质主要有植物生长调节剂、渗透调节物质和非酶促抗氧化物质等三大类。植物生长调节剂是人工合成的一类与植物源激素具有类似生理和生物学效应的物质,主要包括多胺(PA)、油菜素内酯(BR)、脱落酸(ABA)、一氧化氮(NO)等,这些物质大多具有促进蔬菜植物生长发育、提高产量、改善品质以及增强植株抗逆性等功能。
1)PA 高等植物体内主要存在腐胺(Put)、亚精胺(Spd)和精胺(Spm),广泛参与植物生长代谢过程,如基因复制和表达,生长发育、衰老、生物膜稳定、酶活调节以及响应生物和非生物胁迫[26-27]。外源Put能够调节盐胁迫下黄瓜PSⅡ捕光天线蛋白的转录和翻译水平,提高热耗散能力[26];同时,Put可以诱导环式电子传递能力的加强,避免PSⅠ过度还原,从而缓解盐胁迫诱导黄瓜幼苗的氧化伤害[28]。高温胁迫下,喷施1 mmol·L-1Spd明显增加生菜叶绿素含量,减少胆色素原向尿卟啉原Ⅲ的转化过程,抑制原卟啉Ⅳ的过量积累,避免叶绿体的氧化损伤,有效减轻高温对生菜的伤害[29]。外源Spm可促进呼吸爆发氧化酶(RBOH)活性,使质外体产生更多的H2O2,激活抗氧化基因的表达,提高超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)和谷胱甘肽还原酶(GR)的活性,从而促进盐碱胁迫下番茄幼苗的生长,暗示Spm诱导的番茄幼苗对盐碱胁迫的耐受需要依赖于RBOH1的H2O2信号分子功能[30]。黄瓜植株经外源Spd处理后,诱导了RBOH1表达,促进了根系H2O2积累,而H2O2作为信号分子诱导自噬体相关基因(ATGs)表达和自噬体形成,吞噬了盐胁迫下受损的蛋白质或蛋白质聚集体以供进一步利用,从而提高黄瓜幼苗耐盐性[31]。
2)BR 外源施用BR显著减轻Cd胁迫诱导的番茄植株生长抑制,显著降低番茄各组织中Cd的积累,与钙、水杨酸复合施用,还能进一步减轻Cd胁迫[32]。BR预处理可以降低脂质过氧化对膜的损伤和ROS的产生,提高冷处理下冬萝卜和油菜的气孔导度、光合效率、抗氧化能力[33]。干旱胁迫下,外源施用BR通过降低MDA含量和增加脯氨酸含量以及抗氧化酶和硝酸还原酶的活性,减少产量损失,提高不同基因型豇豆的耐旱性[34]。BR类似物预处理可以改善干旱条件下植物叶片的水分状况,降低脂质过氧化水平,促进脯氨酸和总可溶性糖的积累,提高作物耐旱性。此外,BR对胁迫条件下蔬菜作物的营养品质也有较为明显的调控作用。研究表明,紫外线B(UV-B)抑制芽苗菜PSⅡ的活性,但BR处理能缓解UV-B引起的不利影响,提高豆芽黄酮、游离氨基酸和光合色素的含量,以及苯丙氨酸解氨酶(PAL)和2-二苯基-1-苦基肼(DPPH)自由基的清除能力,从而保持采后小黑豆芽的高营养成分[35]。BR处理可以抑制果实冷藏期间的冷害和果肉褐变,可能归因于BR能够保持细胞膜完整性、降低总酚含量以及降低PAL、多酚氧化酶(PPO)和过氧化物酶(POD)活性[36]。
3)ABA 研究表明,ABA处理显著提高番茄幼苗含水量和叶绿素水平,降低脯氨酸的积累,增强植株对干旱胁迫的适应性[37]。氢气通过调节番茄幼苗中ABA的生物合成和ABA信号转导基因的表达来增强抗旱性[38]。钴胁迫下,施用ABA通过激活番茄植株APX、SOD、POD和CAT活性,减轻氧化伤害,并能有效减少根和芽中钴含量的积累,从而提高植株对钴的耐受性[39]。Cd污染的土壤接种产ABA的细菌后,减轻了Cd 诱导的不结球白菜光合作用抑制和氧化损伤,增加了生物量及可溶性糖和可溶性蛋白质含量,降低Cd 在不结球白菜体内的积累,使受Cd污染的土壤种植蔬菜成为可能[40]。ABA处理可有效增强抗氧化能力,降低番茄幼苗ROS和MDA水平,提高脯氨酸、内源褪黑素(MT)、光合色素含量,促进氧化还原基因、耐盐相关基因表达上调,进而提高植株耐盐性[41]。
4)NO 低温胁迫下,喷施NO供体SNP能显著增加茄子幼苗的生物量、光合色素、抗氧化酶活性、可溶性蛋白和脯氨酸含量,显著降低MDA含量,提高茄子幼苗对低温胁迫的抗性[42]。NO预处理种子或叶面喷洒提高了缺水条件下西兰花植株的生物量、茎长、叶绿素含量、总酚类含量、总可溶性蛋白含量以及SOD和POD活性,还发现叶面施用NO比播种前施用在增强西兰花植物的耐旱性方面更有效[43]。研究发现外源NO通过降低不结球白菜种子的胚根和胚芽中Na+含量,提高K+和K+/Na+比值,来改善盐胁迫下种子发芽和幼苗生长[44]。外源NO能够稳定植物放氧复合体结构,提高质体醌A(QA)向次级电子受体质体醌B(QB)的电子传递速率及质体醌(PQ)接受电子的能力,使QA快速进入氧化状态,促进电子传递顺利进行,缓解低温胁迫诱导的光合抑制。NO处理显著提高Cd和Pb复合胁迫下植株抗氧化酶的活性以及Ca和K含量,抑制植株体内Cd和Pb水平,降低脯氨酸、MDA和H2O2含量,从而减轻Cd和Pb对辣椒植株的伤害[45]。
2.2.2 生物防治近年来,科研人员研究发现了大量天敌、微生物及其代谢产物,广泛用于设施蔬菜病虫害的生物防治,并取得了较好的防治效果。天敌生物种类多样,主要有天敌昆虫、捕食螨和昆虫病原线虫。通过多年研究发现,赤眼蜂对甜椒和番茄上的棉铃虫、小菜蛾有较好的防控效果,东亚小花蝽可有效防治甜椒苗上的蓟马,丽蚜小蜂、草蛉、瓢虫能够防治茄果类蔬菜和瓜类蔬菜上的白粉虱和蚜虫,潜叶蝇茧蜂能够防治斑潜蝇[46]。有益微生物及其制剂是改良连作土壤、提高作物抗病性和促进植株生长的有效途径。施用液态和固态微生物菌剂均能显著提高连作土壤脲酶活性,增加植株全氮含量,促进蔬菜产量和品质的提升。以生物菌作为底肥能促进辣椒的生长发育,改良果实品质性状,提高植株抗病性,增产可达20%以上[47]。根际微生物能与植物根系互作,诱导根系分泌一些代谢产物,影响根际土壤微环境,进而促进生长和提高抗病性。研究发现,从药用植物根系分离出的根系微生物,对番茄细菌性溃疡病表现出较高的抑制效果,使番茄发病率下降28.4%~50.0%[48]。植物促生菌NSY50通过改善新陈代谢及活化防御相关蛋白,缓解枯萎病菌对黄瓜植株的伤害[49]。
2.2.3 嫁接嫁接可通过增加叶片抗氧化物质活性和内源多胺含量以及促进脯氨酸和可溶性蛋白合成,降低ROS水平,增强重金属胁迫下植物幼苗的抗性。南瓜作为黄瓜的砧木,具有较高的Na+外排和根系保留能力,可降低盐分离子由根系向地上部的转运,同时促进植株对K+、Ca2+和Mg2+的吸收,改善离子含量比例,从而提高黄瓜植株的耐盐性[50]。番茄嫁接苗的耐盐性主要受砧木耐盐性的影响,其次为接穗,同时,其与番茄体内的氨基酸和活性氧代谢调控密切相关。淹水胁迫下,嫁接苦瓜比自根苦瓜具有更强的抗氧化能力和渗透调节能力、更低的氧化损伤以及更多的适应淹水的不定根结构,耐淹性强于自根苦瓜。低氮水平下,砧木嫁接提高了番茄植株氮的吸收和利用能力,促进不同氮水平的氮代谢,提高番茄产量[51]。砧木基因型的差异,对嫁接植株的耐受性也不同。耐盐性砧木嫁接与不耐盐砧木嫁接相比,叶绿素含量、光合作用能力和脯氨酸含量明显增加,植株的生物量和耐盐性能力更强。不同砧木嫁接均可明显提升复合盐碱胁迫下青椒幼苗根系活力、叶片渗透调节物质含量、抗氧化酶活性和AsA-GSH循环效率,降低活性氧积累水平和膜脂过氧化,从而缓解盐碱胁迫对青椒幼苗造成的损伤,其中以‘优壮’砧木效果最佳;将辣椒嫁接到合适的砧木上,可以克服高温胁迫诱导的氧化伤害,使植株具有更高的相对生长速率、叶面积和光合效率,有效减轻高温胁迫[52]。
2.2.4 抗性锻炼抗性锻炼是指植株经过前期适度的逆境处理后,对再次发生的逆境胁迫表现出较强的抗性,也称为逆境胁迫记忆。生产上,较为常见的是蔬菜育苗过程中的炼苗,一般通过放风、控温(夏季提高温度,冬季降低温度)、适当控水等措施对幼苗进行锻炼,使其定植后能够迅速适应栽培的不良环境,缩短缓苗时间,增强植株对设施春提前或秋延后环境的适应能力。研究表明,低温条件下炼苗,细胞通过质膜流动性和蛋白构象感受冷信号,提高信号途径中转录因子CBF和冷调节基因COR的表达;同时,诱导ROS、NO和Ca2+依赖的蛋白激酶等介导的信号转导途径,进而提高植株的抗逆性[53]。经过低温锻炼的植株在低温胁迫下较未经过预处理的植株具有较高的抗氧化酶活性,其编码基因表达与酶活性表达趋势一致。鹰嘴豆植株经低温锻炼后,幼苗电解质外渗率(ELI)、MDA含量相比蚕豆褐斑病处理组显著下降,同时增加了鹰嘴豆植物对蚕豆褐斑病的耐受能力[54]。冷等离子体种子处理诱导的 H2O2和ABA 级联信号上调了冷驯化的调节基因,有助于增加脯氨酸和可溶性糖等渗透调节物质的积累和抗氧化酶的活性,从而提高冷胁迫耐受性[55]。
2.3 其他
轮作和间作是有效避免前茬作物病虫害、营养元素失衡、改善土壤理化性状、调节土壤肥力的有效措施之一。设施春黄瓜与景天植物间作,然后再与菠菜和秋黄瓜轮作的模式,与常规轮作制度(大白菜和黄瓜轮作)相比,对春、秋黄瓜枯萎病的抑制率分别为28.4%和57.4%,产量分别提高35.2%和383%;同时,该模式改变土壤微生物群落结构,增强土壤酶活性,并能有效降低土壤总镉和植物可利用镉的含量[56]。豆科植物三叶草与花椰菜间作系统的残留物,可以为后续轮作作物生菜提供营养支持,可减少氮肥用量、降低生产成本和减轻环境污染[57]。白菜间作多叶白萝卜,可以明显减少白菜害虫的发生,尤其是嚼叶害虫如甜菜半环虫、夜蛾科和菜粉蝶[58]。
3 蔬菜抗逆栽培生产实践(技术)
在各种生物和非生物逆境胁迫下,蔬菜植物进化出一系列复杂的机制来应对逆境,但植物自身的应急反应并不能实现逆境下蔬菜的高效生产。因此,利用抗逆品种、嫁接技术、逆境锻炼诱导、施用外源调节剂、加强田间管理和设施栽培等方法是提高蔬菜作物抗逆性栽培的主要措施。
3.1 选育优质多抗蔬菜新品种
采用传统和分子育种的方法,成功选育出许多抗逆性强的蔬菜新品种,应用于生产以达到抗逆、高产、优质的目的。甘蓝根肿病的抗性是由多基因控制的数量性状遗传,但由于甘蓝抗性遗传规律较为复杂,获得纯系抗病材料的难度较大,因而育种进程缓慢,而白菜根肿病抗性基因之间相对保持独立,且已经发现8个抗根肿病基因,并选育出一些抗病材料,进行商品化应用[59]。培育出的新一代优质、抗病甘蓝新品种‘中甘628’等中甘系列品种可以抗枯萎病、抗根肿病、耐寒越冬,已推广种植 6.7 万 hm2以上[60]。培育的京春娃系列白菜品种具有抗病毒病、霜霉病和软腐病,抗抽薹能力强,品质佳等特点。培育的黄瓜‘津优316’品种耐低温、弱光,连续坐果能力强,产量突出。
3.2 嫁接栽培
嫁接是提高蔬菜作物抗逆性、抗病性的有效途径之一,在蔬菜生产中已广泛采用。我国蔬菜嫁接技术已在番茄、茄子、辣椒、黄瓜、苦瓜、西葫芦等蔬菜上得到普遍应用,对于防治瓜类和茄果类蔬菜枯萎病、黄萎病、根腐病、青枯病等土传病害,抵御高温、低温、高湿、干旱、盐渍等恶劣环境条件成效显著,是提高蔬菜植株系统抗性和克服连作障碍的有效措施之一。
嫁接后的黄瓜、西葫芦等瓜类蔬菜表现出较强的抗病性、抗寒性、抗盐性和抗旱性;嫁接黄瓜可防止枯萎病,嫁接茄子可防止黄萎病、根结线虫病,嫁接番茄可防止青枯病、枯萎病等发生,砧木嫁接后的植株具有优良的防土传病害效果,一般可达到89.6%以上的抗病性。在低氮条件下,嫁接可增强氮素的吸收和利用,也能够促进番茄植株光能的利用效率,维持植株健康正常生长[52]。
为适应蔬菜规模化种植需要大量嫁接苗,嫁接机应运而生。目前,国内的嫁接机尚处于起步阶段,华南农业大学的辜松教授成功研制出2JC系列的嫁接机,嫁接成功率可达90%以上。国家农业智能装备工程技术研究中心面向设施农业育苗生产,开发研制了TJ-800型瓜、茄科蔬菜自动嫁接机,适用于瓜、茄科蔬菜,生产率为800株·h-1,嫁接成功率可达95%以上。
3.3 逆境锻炼
逆境锻炼已成为蔬菜作物抗性诱导和提高的主要措施,在生产中广泛采用。冬春季黄瓜种子浸种后0 ℃处理1~2 d后,黄瓜植株抗冷性会增强,生长健壮,易成活,提早结瓜7~10 d,早期产量提高20%~30%,黄瓜幼苗定植前15 d,适当降低夜间温度可有效提高瓜苗耐低温能力,促进多花、早开雌花[61]。番茄开花前,对植株进行干旱预处理,可有效降低番茄遭受渍水胁迫引起的减产[62]。
3.4 施用生长调节剂
施用植物源生长调节剂可增强植株系统抗性,不断应用于蔬菜抗逆栽培之中。施用外源多胺可以有效提高番茄、黄瓜等蔬菜作物耐盐性。适宜浓度的外源ALA可以提高番茄低温耐性、黄瓜耐冷性和生菜的耐盐性[63-64]。外源水杨酸增强了西瓜耐低温胁迫能力,增强辣椒的抗旱性、抗盐性等。外源ABA增强番茄幼苗等作物的抗旱性,增强甘蓝型油菜对镉胁迫的抗性,增强番茄的耐盐性[65]。
叶面喷施氨基酸、甲壳素、海藻精等生物叶面肥,有利于养护叶片,避免卷叶、干叶和黄化失绿现象,可在苗期结合病虫防治进行;结合浇水可冲施养根素或激抗菌冲施肥等生物制剂,具有抗菌、驱虫、促进生根、补充营养、培育壮苗、增加产量的效果。外源施用γ-氨基丁酸(GABA)可以为幼苗提供氮素营养,作为氨基酸肥料广泛使用,可提高不同耐性植株的耐盐能力[66]。
3.5 加强田间管理
在整地时应考虑所种植蔬菜作物种类及品种的需求,采用不同的土壤种植方式和栽培密度,以缓解根际缺氧和养分亏缺胁迫。施肥时应适当增施磷、钾肥,多施有机肥,控制果菜类作物的负载量,维持植物营养生长与生殖生长的平衡,使其保持良好的营养状况,提高植株抗病能力[67]。应合理施用磷、钾肥,适当控制氮肥。磷可促进有机磷化合物的合成,提高植株的抗旱性;钾肥能改善植株体内的糖类代谢,降低细胞的渗透势,促进气孔开放,有利于光合作用,提高植株整体系统抗性。
生物炭可以提高土壤有机质及其他养分含量,改善土壤物理性状,调节土壤细菌群落结构,抑制病原菌发生,促进植株生长发育,进而提高植株的抗病能力[68]。
3.6 设施栽培
在低温季节,依靠玻璃、塑料薄膜覆盖保温,霜冻、寒流来临时实行熏烟、加温设施等提高温度,有效抵御低温胁迫。在夏秋季高温期,依靠湿帘、风机、遮阳等实施降温,创造凉爽环境。采用防虫网防虫、防病,采用防雨棚防台风、暴雨侵袭;增施CO2提高叶光合作用;应用无滴膜覆盖、清理覆盖物表面雾滴及灰尘等增强设施内光照强度,及时清理老病叶,增加植株间的散射光照,或张挂反光膜、安装补光灯来补充光照等措施,实现冬季、夏季逆境胁迫条件下蔬菜作物优质高效生产。
以新能源、新技术为依托的设施节能增温技术不断发展。孙先鹏等[69]使用太阳能蓄热联合空气源热泵系统进行日光温室加温,能够维持较高的室内空气和土壤温度,降低相对湿度,为作物生育提供适宜环境。何芬等[70]设计了基于碳晶电热膜的温室育苗加温保温系统,可有效提高育苗环境温度,且加温均匀性高,为培育高质量蔬菜种苗提供保障。孙维拓等[71]设计了一套日光温室空气余热热泵加温系统,白天富集室内空气余热储存于蓄热水池中,用于夜间温度较低时散热增温,可实现白天降温除湿、夜间加温降湿的效果。
3.7 化学、物理方法防治病虫草害
化学防治往往会对非靶标生物产生负面影响,需要与土壤改良剂、生态修复技术配套使用。氯化苦和1,3-二氯丙烯是常用的土壤熏蒸剂,其对土壤进行消毒后再施用土壤改良剂,可改善土壤环境,重建微生物群落结构,更利于作物生长发育。黄腐酸与土壤熏蒸剂棉隆配合施用,对真菌性病原菌的抑制效果更佳。Tian等[72]研究发现,土壤消毒后施用蚯蚓粪和生物炭可有效降低西洋参镰刀菌根腐病的发病率,并增加根际有益细菌的相对丰度。采用土壤高温高湿+石灰氮消毒技术杀灭土传病虫害,利用生物秸秆反应堆、基质土壤调理剂、生物有机菌肥和水肥一体化等进行土壤修复,结合水旱轮/间作新模式,可有效防控连作障碍[73]。
物理防治利用杀虫灯、粘虫板等防护用具,或使用热处理、射线处理等措施,实现驱避害虫、杀灭病菌的目的。马雨晴等[74]发现昆虫对不同波长LED灯具有选择性,可用于开发LED单波长诱虫灯,针对性诱杀害虫,降低对非靶标昆虫的伤害。吴霜等[75]对不同波段光谱诱虫灯的效果进行比较,发现440~445 nm蓝光灯诱集的菜地昆虫最多,提出根据昆虫活动规律,不同时段使用相应的诱虫灯以有效诱杀菜地害虫,保护天敌昆虫。
4 蔬菜抗逆栽培理论与实践展望
科研工作者和生产人员经过不懈努力在逆境生理、抗逆理论研究和抗逆栽培生产实践方面取得了长足进展,保障了蔬菜生产的正常进行和产品供给。今后将在以下方面展开蔬菜作物逆境生理与抗逆栽培研究。
4.1 围绕信号分子通路,阐明逆境伤害蔬菜作物的作用机制
众所周知,各信号分子、功能基因等组成了错综复杂的逆境信号转导网络。考虑到多重胁迫下各信号组分之间可能存在新的相互作用,学者们将目光从单一胁迫转向了多重胁迫,发现不同组合胁迫对蔬菜有着不同的效应。高温、干旱等可能增加蔬菜作物对病虫害的敏感性,且生物胁迫和非生物胁迫在蔬菜作物体内诱发的信号网络往往存在高度的交集。然而,研究者对于生物与非生物组合胁迫下蔬菜复杂信号通路的认知较为零碎。因此,应深入研究不同复合胁迫信号通路及其调控网络,以期为蔬菜抗性品种选育及抗逆栽培提供理论依据。
ROS具有双重作用,其处于低浓度时在各种生命活动和环境响应中扮演着重要的角色,高浓度时则对细胞产生毒害。ROS能将叶绿体和线粒体中的氧化还原状态信号传递到细胞核从而调控核基因的表达,但对胁迫下ROS如何调控核基因的表达、影响细胞周期调节和细胞程序性死亡(PCD)的认识仍有限,ROS的代谢机制和与其他信号组分的互作情况也需要深入研究。多组学的迅猛发展成为系统生物学繁荣的重要推动力,也为蔬菜的逆境生理研究提供了丰富的信息,然而如何从这些海量数据中筛选有效信息,并加以整理和比较,从而明确蔬菜胁迫伤害的成因,仍是亟待解决的重要问题。
4.2 挖掘优质抗逆基因,揭示蔬菜作物适应逆境的分子机制
为进一步阐明蔬菜作物逆境响应机制,应充分利用多组学原理和技术挖掘抗逆关键基因、蛋白和代谢产物,并鉴定和分析它们的功能,运用正向、反向遗传学手段相结合验证关键基因的功能,为抗逆新品种选育提供依据。
目前,已克隆出多个与蔬菜生长发育、抗逆性相关的基因,并发现这些基因过量表达或抑制表达,可在一定程度上提高非生物胁迫下蔬菜作物的抗性。科研工作者如何更加有效地应用这些基因,将其应用到蔬菜作物抗病和耐逆转基因研究中,是抗性转基因研究中的重要方向。大多数蔬菜作物响应胁迫不是由单个抗性基因决定,而是复杂的数量性状,并由多种微效抗性基因共同作用。病虫害是限制蔬菜栽培生产的关键因素,基因工程的应用为未来蔬菜病虫害防治提供了有效的技术手段,如抗病毒基因工程、抗真菌基因工程等技术,通过提高植物体内的壳体蛋白、几丁质酶等物质含量,从而提高植物对病虫害的系统防疫能力。
近年来,随着智慧植物工厂、表型组学等平台以及功能基因组学、基因编辑等生物技术的发展,为蔬菜抗性基因克隆、鉴定以及高效编辑作物抗性基因奠定基础,加速基因工程育种的进程。例如利用响应盐碱胁迫、高温、低温或抗病基因的启动子驱动的基因编辑技术,通过对某些基因进行特定编辑,进而提高蔬菜作物对非生物胁迫的耐受性。
4.3 探索农艺农技融合,重视以应用研究为基础的抗逆品种、技术和产品开发
在复杂多变的温、光、水、气、土、生物等蔬菜生育环境下,必须选择优良的品种,采取有效的技术,利用可行的产品,克服不利于蔬菜生育的逆境因子,创造有利于蔬菜生育的环境条件。
长期以来抗逆优质高产蔬菜新品种选育取得了明显进展。但需要特别重视抗新流行病害品种的研究,如番茄黄化曲叶病毒、十字花科蔬菜根肿病、甘蓝枯萎病、黑腐病等的抗病品种选育;要重视露地栽培时应对低温、高温等灾害性天气的蔬菜抗逆育种,以及设施栽培专用的耐弱光、抗高湿、抗高(低)温、抗盐碱、抗病蔬菜品种选育,需要选育适合设施长季节栽培的抗逆性强的蔬菜品种,需要选育耐裂球裂果、适合远距离运输、适合机械化收获的专用品种。
目前对嫁接技术的研究主要集中在提高接穗抗性方面,关于砧木品种研究较少,尤其是抗虫砧木的选育,需要加强对适宜砧木品种的研究;由于传统嫁接苗的生产需要大量劳动力,且手工嫁接工作效率低下,成活率不能保证,蔬菜嫁接自动化和嫁接机器人技术在未来的嫁接技术上前景广阔,需要不断开发并完善嫁接育苗机械配套装备。
施用植物源生长调节剂和微生物制剂是非常有效的促进蔬菜植株生育、提高植株抗逆性的方法。市场上已有多种油菜素内酯、多胺、5-氨基乙酰丙酸等外源生长调节剂产品,但质量参差不齐,有待改良和提高;芽胞杆菌等促生菌微生物制剂和功能性育苗基质等,使用效果不稳定;产品不能有效协调作物生长,提高作物品质;也存在相应自动化喷施设备不配套、劳动效率低等问题。因此,需要加强研制绿色环保、标准化、高效复合型、多功能助剂产品,同时着力研究自动化喷施设备,利于外源生长调节剂和促生菌微生物制剂的有效推广应用。
另外,抗逆优良品种与抗逆农艺技术、产品如何有效配套结合使用需要研究,亟待加强抗逆栽培技术创新和轻简增效技术集成。
