烟草浸提液对番茄晚疫病菌的化感抑制效应机理研究
2022-11-18刘嘉方云兴福
刘嘉方,李 蕾,云兴福
(1.内蒙古农业大学园艺与植物保护学院,呼和浩特 010019;2.内蒙古科技大学包头师范学院,包头 014030;3.满洲里俄语职业学院,满洲里 021400)
番茄晚疫病由致病疫霉Phytophthora infestans(Mont.)de Bary侵染所致,是番茄生产上的毁灭性病害[1]。致病疫霉属卵菌纲 Oomycetes,主要侵染番茄的叶和茎,在潮湿、凉爽的环境条件下侵染速度极快,会加速病害发生,造成严重损失[2]。传统生产上多采用化学农药对番茄晚疫病进行防治,但农药的大量使用会对环境造成污染,目前需要寻找一种绿色环保、生态有效的防治方法。
化感作用(allelopathy)是植物通过释放化学物质进而影响自身或其他微生物的生长发育[3]。利用植物化感作用可以有效防治蔬菜病害,如大蒜的根系提取物对番茄晚疫病菌[4]、辣椒疫霉菌Phytophthora capsici、番茄青枯病菌Ralstonia solanacearum和番茄灰霉病菌Botrytis cinerea[5]等多种病原菌均有显著抑制效果;韭菜挥发物[6]、西芹提取物[7]和蚕豆根系分泌物[8]能抑制枯萎病菌Fusariumspp.的菌丝生长和孢子萌发,降低病原菌的致病力。烟草Nicotiana tabacumL.是一种一年生或有限制性的多年生的草本植物,全株被细毛,基生稍木质化,在夏天和秋天开花[9]。作为一种高敏感的化感植物,烟草的根系分泌物[10]、叶片浸提物[11]以及残体腐解物[12]等不同部位均具有化感效应。烟草中机酸类、烟碱类、酮类物质及钾元素等含量较高,可促进作物生长,并有一定的抗病效果[13]。已有研究发现,低浓度的烟草浸提液对水稻[14]和莴苣[15]等作物的种子萌发和幼苗生长具有促进作用;烟草化感物质能提高植物的光合能力,对真菌、细菌和放线菌有明显的抑制作用[16],对蔬菜病虫害也有较好的防治效果[17]。因此,烟草化感作用在番茄晚疫病防治方面具有较大潜力。
本研究通过菌丝生长和致病力测定探讨烟草浸提液对番茄晚疫病菌的化感作用,并连续处理多代,直至获得 1株符合疫霉菌弱毒评价标准的 PIAS-R菌株(番茄晚疫病菌弱毒菌株英文全称Phytophthora infestansattenuated strain,PIAS)。并进一步通过测定PIAS-R菌株的孢子萌发率、菌丝生物量和细胞壁降解酶活性,观察菌丝体形态特征及其转录组分析比较,解析烟草浸提液对番茄晚疫病菌的化感抑制效应机理,以期为利用烟草防治蔬菜病害或研制植物性抑菌剂提供理论依据。
1 材料与方法
1.1 材料
供试材料:干烟草由辽宁省朝阳烟草基地提供,品种为“大叶葵”;番茄晚疫病病原菌由中国农业科学院植物保护研究所提供;番茄种子购自内蒙古农业科学院,品种为“粉红918”。
菌种培养:试验前期将保存的番茄晚疫病病原菌接种在PDA培养基上,于25 ℃恒温培养箱中倒置培养,7 d后菌丝长满整个培养皿,菌落形态完整,可进一步扩繁备用。
马铃薯葡萄糖琼脂固体培养基(potato dextrose agar,PDA):马铃薯200 g、葡萄糖20 g、琼脂16 g、蒸馏水1 L;马铃薯葡萄糖液体培养基(potato dextrose,PD):马铃薯200 g、葡萄糖20 g、蒸馏水1 L;改良MS培养基(Murashige & Skoog,MS):KNO32 g,KCl 0.5 g,FeSO40.01 g,K2HPO41 g,MgSO40.5 g,VB10.1 mg,L-天冬酰胺 0.5 g,羧甲基纤维素钠盐 10 g或柑橘果胶 10 g,蒸馏水1 L。
1.2 方法
1.2.1 烟草浸提液的制备 用天平称取烟草根、叶6 g等量3份置于100 mL锥形瓶中,分别以1:10比例加入蒸馏水,25 ℃条件下摇床振荡浸提(240 r/min),1 d后取出置于超净工作台上,然后过滤,第1次用定性滤纸过滤除滤渣,第2次经细菌过滤器(0.22 μm)过滤除菌2次,即得烟草根、叶浸提液母液,浓度为100 mg/mL;将烟草浸提液母液用无菌水稀释2倍作为处理浓度(50 mg/mL),贮于冰箱(4 ℃)中备用。PI-R代表根浸提液处理,PI-L代表叶浸提液处理。
1.2.2 不同烟草浸提液每代处理后筛选番茄晚疫病菌弱毒菌株 烟草对番茄晚疫病菌的化感作用:在无菌条件下,用直径为0.6 cm的打孔器打取番茄晚疫病病原菌株,用接种针将菌饼移入不同PDA培养基(121℃,30 min)中。以培养皿中添加2 mL各处理浸提液和18 mL培养基的平板为处理,以添加20 mL培养基的平板为对照(CK)。菌丝面向下,每皿取1块放于中央,25 ℃恒温培养,菌丝生长2 d后采用十字交叉法测量菌落直径(cm)[18]。每隔1 d测1次,共测5次。持续培养6 d后长出的菌落为第1代。用打孔器分别打取第1代各处理和对照菌饼放入上述对应不同培养基中,长出的菌落为第2代,依此类推,直至处理培养8代,测定每一代的菌落直径,5次重复。菌落直径(cm)=测量菌落直径平均值-0.6 cm,化感效果(%)=(对照菌落直径-处理菌落直径)/对照菌落直径×100。
烟草对番茄晚疫病菌的抑制效果:取25 ℃条件下在PDA平板上培养6 d的每一代各处理番茄晚疫病菌,在平板中央加入适量预冷的无菌水,用涂布棒轻轻刮取菌丝表面,2层纱布过滤即得到孢子囊悬浮液,在显微镜下调整浓度为5.0×104cfu/mL,将配制好的悬浮液在4 ℃低温箱静置2~3 h,释放的游动孢子用于接种。以接种番茄晚疫病病原菌的孢子囊悬浮液为对照(CK)。将露白的番茄种子播种在直径为8 cm的塑料育苗钵中自然生长,每钵1株,待番茄植株长至6~8叶期时,用手持喷壶在植株叶片正反面分别均匀喷施不同处理的悬浮液,以叶面上有明显小液滴为度。接种处理在下午 6-7点进行并在喷施完扣上小棚保温保湿,在接种21 d后依据病情分级标准[19]进行番茄晚疫病调查并计算病情指数,每处理24株,3次重复。病情指数=Σ(各级病叶数×相对级数值)/(调查总叶数×最高级数)×100。
第一,福建小学学校的数量和施教人数逐年增加。“二十九年改制之初,设中心学校875校、国民学校3700校、私立小学586校、共4161校① 原文计算有误,应为5161校。,施教人数小学部514490人。三十年中心学校增为1164校、国民学校增为3782校、私立小学516校、共5462校、施教人数小学部648551人。三十一年中心学校增为1349校、国民学校增为4041校、私立小学468校、共5732校②原文计算有误,应为5858校。、施教人数小学部634562人。”[27]13三年间,小学学校的数量增长了1571所,施教人数仅小学部就增长了120072人。
1.2.3 PIAS-R生理指标的测定 将1.2.2筛选到的番茄晚疫病菌弱毒菌株PIAS-R进行生理指标的测定,以番茄晚疫病病原菌株为对照(CK),5次重复。
孢子萌发率的测定:参照黄英菊等[20]的方法制备浓度为1×107cfu/mL的PIAS-R和CK菌株游动孢子悬浮液。将制备好的游动孢子悬浮液在黑暗10 ℃条件下放置3 h,刺激游动孢子萌发。每次镜检50个游动孢子,计算孢子萌发率(注:芽管长度达孢子直径一半时认为萌发),孢子萌发率(%)=孢子萌发数/孢子总数×100。
菌丝生物量的测定:在无菌条件下,用接种针将等龄PIAS-R和CK菌饼移入30 mL PD培养基中,25 ℃黑暗培养6 d后收集菌丝,蒸馏水洗涤、晾干,记录菌丝生物量(g)[21]。
粗酶液的制备:无菌条件下,分别取等龄PIAS-R和CK菌落,移至事先灭菌的50 mL产酶培养基(改良MS和PD培养基)中,每个三角瓶接种直径0.6 cm的5块菌饼,并在26 ℃下恒温振荡培养6 d,将收集的培养液用3层纱布过滤,除去孢子和菌丝体,滤液在4 ℃下12000×g离心15 min,再次过滤后弃去沉淀,取上清液为粗酶液,将其保存在4 ℃下待测。测定各项细胞壁降解酶指标。β-葡萄糖苷酶活性测定参照高晓敏[22]的方法;纤维素酶活性测定参照 Eveleigh等[23]方法;果胶酶活性测定参考姜立春等[24]方法;蛋白酶活性测定参照赵爽等[25]方法。
1.2.4 PIAS-R菌丝体形态观察 用相机对1.2.2筛选到的番茄晚疫病菌弱毒菌株PIAS-R菌落进行直接拍照,以番茄晚疫病病原菌株菌落为对照(CK)。同时从PDA平板上挑取菌丝用光学显微镜(Olympus BX53)以200倍放大倍数观察菌株的形态结构,选择典型特征视野对菌丝进行拍照和描述。
1.2.5 PIAS-R转录组测序 将1.2.2筛选到的番茄晚疫病菌弱毒菌株PIAS-R用无菌镊子小心刮取菌丝体300 mg,放入提前标记的冻存管中,液氮冷冻,-80 ℃超低温冰箱保存,备用。以番茄晚疫病病原菌株菌丝体为对照(CK),3次重复。利用DNase提取样品的总RNA;用1%琼脂糖凝胶电泳和Agilent 2100 Bioanalyzer检测总RNA的完整性、纯度和浓度;文库的构建参照TruSeq Stranded mRNA LT Sample Prep Kit的说明进行;使用Illumina HiSeqTM 2500平台进行无参转录组测序,由上海欧易生物医学科技有限公司完成。
1.3 数据统计与分析
采用WPS(2019)软件处理原始数据,SPSS(20.0)统计软件对试验数据进行方差分析,T检验法(T-test)、最小显著差异法(LSD)和Duncan法(SSR)多范围检验各处理间的差异显著性。转录测序中用eXpress[26]软件分析基因FPKM[27],用DESeq[28]软件进行基因的差异表达分析。筛选出P<0.05且|log2fold change| ≥1的差异基因进行GO和KEGG[29]富集分析,以判定差异基因主要影响的生物学功能或者通路。
2 结果与分析
2.1 烟草浸提液每代处理后番茄晚疫病菌弱毒菌株的筛选

表1 不同烟草浸提液对番茄晚疫病菌的化感作用Table 1 Allelopathy of different tobacco extracts on Phytophthora infestans
2.1.2 烟草浸提液对番茄晚疫病菌的抑制效果 在番茄植株6~8叶期,用每一代各处理菌株的悬浮液对番茄植株进行人工接种。从接种后21 d开始调查不同菌株的病情指数(图1)。随接种代数的增加,每一代各处理的病情指数均始终低于CK且有显著差异。在接种后21 d,每一代PI-R处理的病情指数较CK显著降低37.31%、41.43%、47.88%、49.30%、60.97%、70.56%、76.63%和82.45%。接种后期PI-R处理的病情指数显著低于PI-L处理。在连续处理8代后,烟草浸提液对番茄晚疫病菌有明显的抑制效果,其中PI-R处理菌株在第7、8代的病情指数连续<10,表明已经由番茄晚疫病菌的强致病性菌株逐步变为弱致病性菌株,将这株番茄晚疫病菌弱毒菌株命名为PIAS-R。此外,随继代培养代数的增加,CK菌株的病情指数总体表现为下降的趋势,说明继代培养对番茄晚疫病菌的致病力有一定的抑制作用。
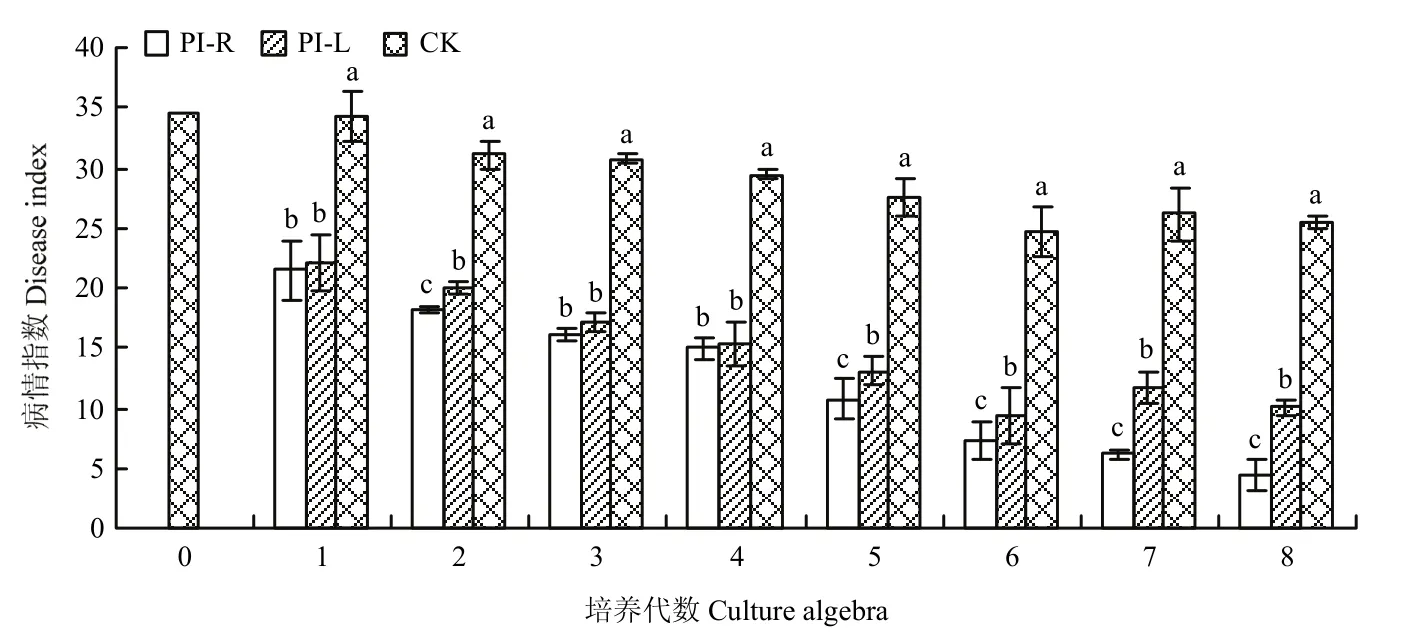
图1 不同烟草浸提液处理后番茄晚疫病菌致病力的变化Fig.1 Changes of pathogenicity of Phytophthora infestans after treatment with different tobacco extracts
2.2 PIAS-R孢子萌发、生物量和酶活性的变化
对PIAS-R和CK菌株的孢子萌发率、菌丝生物量和细胞壁降解酶活性进行测定(表2)。PIAS-R菌株的孢子萌发率和生物量为31.4%和0.224 g,较CK菌株显著降低55.01%和35.45%。PIAS-R菌株分泌的4种细胞壁降解酶活性均低于CK菌株且有显著差异。β-葡萄糖苷酶、纤维素酶、果胶酶和蛋白酶活性较对照降低33.79%、51.11%、60.11%和37.63%;其中纤维素酶和果胶酶活性最低,为10.15 μ/(g·min)和110.52 U/mL。说明PIAS-R菌株分泌更低活性的纤维素酶和果胶酶,且分泌的果胶酶活性下降程度大于纤维素酶。

表2 PIAS-R的孢子萌发率、菌丝生物量和酶活性Table 2 Spore germination rate, mycelial biomass and enzymes activities of PIAS-R
2.3 PIAS-R菌丝体形态结构的观察
2.3.1 PIAS-R菌丝体形态比较 对25 ℃恒温培养6 d后的PIAS-R和CK菌株进行拍照观察(图2)。通过观察菌株的生长状态,发现不同菌株的形态特征有较大差异(表3)。PDA平板透明光滑,菌丝都以接种块为中心,生长成圆形或近圆形的绒毛状或絮状菌丝体。与CK菌株相比,PIAS-R菌株的菌落颜色和表型不同且生长速率减慢。

表3 PIAS-R的形态特征Table 3 Morphological characteristics of PIAS-R

图2 PIAS-R在PDA培养基上生长6 d后的菌落状态Fig.2 Colony status of PIAS-R after 6 d growth on PDA medium
2.3.2 PIAS-R菌丝体显微结构的观察 从PDA平板上挑取PIAS-R和CK菌丝体,用光学显微镜观察不同菌株的显微结构(图3)。在放大200倍的显微镜视野下观察PIAS-R菌丝体出现不同种类的畸形变化,包括菌丝分枝增多(图3A),原生质分布不均匀(图3B),菌丝局部膨胀(图3C)、变短(图3D)、扭曲和透明(图3E,F),菌丝相互缠绕(图3G)且顶端膨大(图3H);而正常生长的CK菌丝体光滑顺直、粗细均一、内含物饱满且原生质分布均匀(图3I-L)。

图3 PIAS-R的微观变化Fig.3 Microscopic changes of PIAS-R
2.4 PIAS-R转录组分析
2.4.1 总RNA质量分析及差异表达基因的筛选 对6 个样本进行RNA-Seq测序后获得总 RNA 质量结果(表 4)。根据测序数据的质量统计,可进行后续相关数据分析。将 RNA读取映射到基因组中,分析PIAS-R和CK菌株的差异表达,绘制差异表达基因(differentially expressed genes,DEGs)统计图(图4),PIAS-R相对于CK有2651个基因(P<0.05,|log2fold change| ≥1)在表达水平上存在显著差异,其中1086个DEGs上调,1565个DEGs下调。
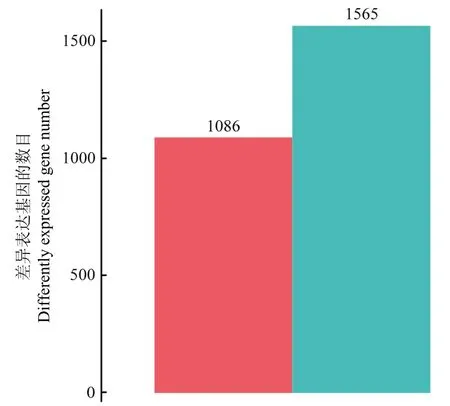
图4 RNA差异表达基因对比图Fig.4 RNA statistics and comparison plot of differently expressed genes (DEGs)

表4 总RNA的质量统计Table 4 Quality results of total RNA
2.4.2 GO富集分析 对PIAS-R与CK之间的DEGs进行GO 功能富集分析,选取富集最显著的前30个分类条目,绘制GO富集柱状图(图5)。在 GO 数据库中,DEGs根据功能分为生物过程(biological process)、细胞组分(cellular component)和分子功能(molecular function)3 大类。根据已经报道的与菌株生长发育和致病力相关的GO功能分类,生物过程中的内吞标志位点集合(eisosome assembly)、纤维素分解代谢过程(cellulose catabolic process)和脂质转运(lipid transport)均在上/下调DEGs富集最多的条目;细胞组分中内吞标志位点(eisosome)、脂滴(lipid droplet)和细胞壁(cell wall)均在上/下调DEGs富集最多的条目;分子功能中的纤维素结合(cellulose binding)、纤维素酶活性(cellulase activity)和单酚单加氧酶活性(monophenol monooxygenase activity)均在上/下调DEGs富集最多的条目。

图5 差异表达基因GO分类注释top 30Fig.5 Top 30 GO classification of DEGs
2.4.3 KEGG富集分析 对PIAS-R与CK之间的DEGs进行KEGG 显著性通路富集分析,如图6所示。结果表明,DEGs主要集中在其他氨基酸代谢(metabolism of other amino acids)、脂质代谢(lipid metabolism)、能量代谢(energy metabolism)、碳水化合物代谢(carbohydrate metabolism)和氨基酸代谢(amino acid metabolism)通路中。

图6 差异表达基因KEGG富集通路Fig.6 Enriched KEGG pathways of DEGs
3 讨论
烟草中的有机酸类[30]和生物碱类[31]物质对常见的植物病原菌都有一定的抑制作用。病原菌弱毒菌株一般可通过自然分离法[32]、化学刺激法[33]和植物提取物化感法[34]等筛选获得。在筛选番茄晚疫病菌的弱毒菌株时,根据疫霉菌致病力分类标准[35]把病情指数<10的菌株认定为弱致病性菌株。本研究中,烟草根、叶浸提液对番茄晚疫病菌的菌丝生长表现为化感抑制作用,且随处理代数的增加抑制作用累积增强;培养后期PI-R处理的菌落直径始终低于CK且有显著差异。经过连续8代处理,烟草浸提液对番茄晚疫病菌的抑制效果逐渐提高;接种后每一代各处理菌株的病情指数均始终低于CK且有显著差异;其中,PI-R处理菌株在第7、8代的病情指数连续<10,据此确定第8代PI-R处理的番茄晚疫病菌为番茄晚疫病菌弱毒菌株PIAS-R。另外,由于连续处理代数较多,本试验仅仅利用人工接种对每一代不同处理的番茄晚疫病菌进行致病力测定,而没有对接种发病后各代番茄植株上的病原菌进行重新分离,并再次接种测定其致病力的稳定性,这在后续的试验过程中会进一步探讨研究。
李杨等[36]观察表明不同油茶炭疽病菌Colletotrichum gloeosporioides的致病力可能与其孢子萌发率存在一定关系。朱荷琴等[37]对棉花黄萎病菌Verticillium dahliae不同致病菌株的生物学特性进行了比较,发现强致病菌株的生物量都高于弱致病菌株。Khaledi等[38]发现,镰刀菌F.graminearum强致病力菌株产生的细胞壁降解酶活性水平明显高于弱致病力菌株。王前前[39]研究发现核盘菌Sclerotinia sclerotiorum低毒菌株 SZ-150菌丝较细小,分枝较多,较为密集。黄娟等[40]研究表明核盘菌的致病力减弱和生长抑制与其菌丝结构和形态变异有关,其弱致病型 Ep-1PN具有菌落扩展、菌丝顶端分枝异常、原生质不均匀和内部结构不完整等特点。
番茄晚疫病菌以孢子或菌丝体形式在植株病残体或土壤中存活,通过气流或雨水侵染到番茄植株上[41]。本研究中,PIAS-R菌株的孢子萌发率、菌丝生物量和细胞壁降解酶活性均低于CK菌株且有显著差异,菌丝体形态结构出现畸形变化(菌落变薄、颜色变浅,菌丝分枝增多、顶端膨大、变短、扭曲和透明,原生质分布不均匀),推测烟草是通过抑制菌株的孢子萌发、菌丝生长和自身分泌的酶活性,破坏菌丝体形态结构,影响对寄主的侵染能力,从而来发挥抑菌作用的。
内吞标志位点是一种大型蛋白复合体,可作为支架诱导膜皱纹或内陷[42]。病原真菌的菌丝体生长受阻或形态发生畸变可能是细胞中的纤维素发生变化[43]。脂滴参与菌丝生长和致病力等表型[44]。本研究中,根据GO功能富集分析,内吞标志位点集合、内吞标志位点、纤维素分解代谢过程、纤维素结合、纤维素酶活性和脂滴均在上调DEGs主要富集的分类条目中。推测烟草通过调控菌体内内吞标志位点、纤维素和脂滴相关编码基因上调表达,进而影响菌株的表型特征、菌丝生长和致病力等。
缺失脂质转运蛋白的突变体在生长、产孢和致病力等方面表现出了诸多缺陷[45]。致病真菌细胞壁对其毒力和致病力至关重要,可以保护菌株免受宿主防御机制的伤害[46]。单酚加氧酶是酪氨酸酶[47],低活性的酪氨酸酶能降低菌株的褐变度[48]。酪氨酸酶是生物体中参与黑色素合成的关键酶[49],缺乏黑色素生成的突变菌株丧失致病能力[50]。本研究中,根据GO功能富集分析,脂质转运、细胞壁和单酚单加氧酶活性均在下调DEGs主要富集的分类条目中。推测烟草通过调控菌体内脂质转运、细胞壁和酪氨酸酶相关编码基因下调表达,进而影响菌株的孢子萌发、致病力和菌落颜色等。烟草具体调控哪些编码基因差异表达来影响菌株的生长和致病力还需进一步研究证实。
综上所述,烟草浸提液对番茄晚疫病菌具有良好的化感抑制作用,其抑菌机理与干扰菌株生理生化代谢,破坏菌丝体形态结构,影响菌体内的基因表达有关。后续需进一步明确烟草内部抑菌活性物质及差异基因的具体调控机制,最终开发出防治番茄晚疫病的有效植物源。
