基于稻瘟病菌毒性基因的穗颈瘟抗性鉴定菌系筛选与应用
2022-11-15阴长发李湘民杨迎青
兰 波,陈 建,阴长发,李湘民,杨迎青
(江西省农业科学院 植物保护研究所,江西 南昌 330200)
水稻稻瘟病是一种世界性的重要真菌病害,在病害流行的年份严重影响水稻产量和稻米品质,严重地区甚至颗粒无收[1]。选育和利用抗病水稻品种是防治稻瘟病最有效的措施,但稻瘟病菌生理小种不断变化且变异较快,育成的抗病水稻品种通常在种植3~5年后极易丧失抗性[2]。因此,稻瘟病抗性鉴定工作对水稻品种的安全布局来说是一项极其重要的工作,只有在水稻育种的源头明确水稻品种的抗性,才能在实际生产中选择抗性好的品种,减少稻瘟病造成的损失[3]。
稻瘟病抗性鉴定主要有田间自然诱发鉴定与室内人工鉴定这2种方法,田间自然诱发鉴定的稻瘟病菌更符合当地菌群的实际情况,且操作简单实用,但该方法往往由于当年气候环境等因素而影响鉴定结果的准确性;而室内稻瘟病鉴定相对可以人为地控制温度、湿度等稻瘟病发病条件,从而充分保证抗性鉴定结果的准确性,但室内稻瘟病鉴定的一个关键因素是稻瘟病菌的选择,鉴定菌株的筛选必须能够代表某一地区菌群的组成情况,这样才能真实地测定水稻品种在该地区的抗性水平。因此有效地筛选具有代表性的鉴定菌系是稻瘟病抗性鉴定特别是穗颈瘟抗性鉴定成功的关键。我国学者根据当地实际情况选择不同的稻瘟病抗性鉴定方法开展了水稻种植品种或抗病资源品系稻瘟病的抗性鉴定,为当地筛选并获得了丰富的抗病品种(系)。郝中娜等[4]采用人工接种和自然诱发鉴定方法鉴定了2013~2017年我国长江中下游稻区水稻品种区域试验的800个籼稻参试品种的稻瘟病抗性。鄢圣敏等[5]利用自然病圃诱发鉴定了9283份水稻品种的稻瘟病抗性,并发现供试材料的叶瘟与颈瘟表现呈显著正相关。李湘民等[6]采用生态多点分批播种法、离体穗段点滴接种法、活体穗苞注射接种法这3种稻瘟病抗性鉴定方法评价了江西省种植面积较大且抗性差异较明显的9个早稻品种的抗性水平。
传统的稻瘟病抗性鉴定菌株筛选是通过苗期接种某些水稻鉴别品系后的抗致病型来确定的,但到目前为止还没有一套统一的稻瘟病菌鉴别寄主,这为抗性鉴定菌系的选择带来了困难。早在1961年日本学者根据各水稻品种对稻瘟病菌株的反应型确立了第一套稻瘟病菌小种鉴别品种[7]。在20世纪60年代中期,美、日两国合作研究筛选出一套由8个籼、粳稻品种组成的国际鉴别品种,但其抗性基因组成不明且小种鉴别力低因而未得到广泛应用[8]。在20世纪70年代,日本学者山田等[10]根据Flor[9]的基因对基因学说,筛选出一套基于抗性基因分析的9个粳稻“单基因”鉴别品种。我国学者于1980年筛选出特特普、珍龙13、四丰43、东农363、关东51、合江18和丽江新团黑谷(LTH)等7个水稻品种作为中国稻瘟病菌小种鉴别品种,但因其抗性基因组成不明,小种鉴别力低,在国内特别是北方粳稻区的应用受到局限[11-12]。凌忠专等[13-14]以普感品种丽江新团黑谷(LTH)为轮回亲本,选育出6个单基因近等基因系(LTH-NIL)。2000年,IRRI与日本国际农业科学研究中心(JIRCAS)合作,将24个稻瘟病抗性基因导入到LTH背景,育成31个单基因系(LTH-monogenic lines,LTH-MLs)[15-16]。前期研究报道的抗性鉴定菌株筛选都是在苗期接种以鉴别寄主反应型来确定的,其对穗颈瘟抗性是否具有较好的鉴定能力却无法确定。目前还没有直接证据证明水稻对苗(叶)瘟的抗性与穗颈瘟的抗性具有正相关性。由于稻瘟病菌的毒性主要由致病基因与无毒基因控制,本研究通过特异性引物标记测定区域稻瘟病菌群的致病基因与无毒基因情况,根据测定结果聚类划分成不同的遗传宗谱,再从不同的宗谱中按比例筛选菌株,组成代表该地区稻瘟病菌群的抗性鉴定菌系。通过田间自然诱发鉴定验证其鉴定结果,表明该方法具有高效、准确、省时省力的特点,在水稻稻瘟病特别是穗颈瘟的抗性鉴定方面有很大的应用潜力。
1 材料与方法
1.1 稻瘟病标本的采集
在江西婺源(赣东北)、都昌(赣北部)、丰城(赣中)、井冈山(赣西南)、万安(赣南)这5个地区稻瘟病自然病圃种植高感品种丽江新团黑谷,在半黄熟期采集稻瘟病发病标样,用牛皮纸袋装好,记录好标样的采集地点、时间、寄主品种、采集人等信息。
1.2 稻瘟病菌单孢菌的分离、保存
取采集的稻瘟病标样剪成寸许,浸水18~24 h,待充分吸水后,用清水冲洗2~3次,再用灭菌水冲洗一次,放入垫有U形玻璃棒的保湿培养皿中,置于28 ℃恒温培养箱内暗培养2 d,观察到标样上有灰色或灰绿色的孢子后,采用眉毛挑单孢法,将病部置于眉毛挑单孢显微镜下,挑取单个孢子在PDA斜面培养基中培养。纯培养后转接到放有灭菌滤纸片的PDA培养基培养,待滤纸片长满菌丝,将菌丝挑出在无菌的羊皮纸袋中,置于-80 ℃超低温冰箱保存(图1)。本研究从同一地区分离保存了189个稻瘟病菌单孢菌株,编号为WY1~WY189。

图1 稻瘟病菌滤纸片保存
1.3 稻瘟病菌DNA的提取
将分离保存的稻瘟病菌株活化后,挑取适量菌丝接种到 PDA液体培养基中,置于120 r /min的摇床上28 ℃培养3~6 d,待菌丝体有一定量后用灭菌纱布过滤收集、烘干,于-20 ℃保存备用。之后将收集的菌丝体分装于2 mL的Eppendorf管中,加2颗5 mm灭菌碳钢珠后迅速加65 ℃预热的CTAB提取液,应用全自动样品快速研磨仪进行磨样,按照天根植物基因组提取试剂盒TIANGEN(DP350)说明书中的提取方法对病菌基因组的DNA进行提取,于-20 ℃冰箱保存备用。
1.4 稻瘟病菌无毒基因与致病基因测定
选用已公开报道且克隆的8个稻瘟病菌致病基因与8个无毒基因作为检测目的基因,8个稻瘟病菌致病基因分别为MPG1、MPS1、MagB、CPKA、MNH1、MgYCA1、MgS11、MgATG5,8个稻瘟病菌无毒基因分别为Avr-pia、Avr-pit、Avr-pik、Avr-pi-ta、Avr-pizt、Avr-co39、ACE1、PRE1。
根据以上基因序列设计特异性引物标记(表1)[17-19]。PCR总反应体系为20 μL,包括DNA模板1 μL(≤200 ng)、0.2~0.4 μmol/L正、反 向 引 物各1 μL、2.5 mmol /L dNTP 0.5 μL、Taq DNA聚合酶 0.2μL、10×Buffer(含Mg2+)2 μL,ddH2O 14.3 μL。PCR反应程序: 95 ℃预变性5 min; 94 ℃变性30 s,55~57 ℃(按引物设定温度)退火1 min,72 ℃ 延伸60~90 s (根据片段大小而定),共35个循环; 最后72 ℃延伸 10 min。将获得的PCR产物保存于4 ℃冰箱,用1%琼脂糖凝胶电泳进行检测,最后在凝胶成像系统的紫外灯下拍照保存。

表1 稻瘟病菌致病基因与无毒基因分子检测引物序列
1.5 聚类分析与穗颈瘟抗性鉴定菌系的确定
根据PCR扩增条带的结果,有条带则记为“1”,无条带则记为“0”,构建基于SSR的“0-1”数据库;应用NTSYS数据处理系统计算遗传距离,以菌株DNA扩增条带的位置相似水平为基准,用最长距离法对菌株群体遗传多样性进行聚类分析。通过聚类可以将菌株归类成不同的密切相关的组,每一组称为一个遗传宗谱;根据不同的相似系数可以划分成不同数量的遗传宗谱。将每个宗谱菌株占总菌株的百分比作为在抗性鉴定菌系中的百分比,随机挑选菌株组成穗颈瘟抗性鉴定菌系。
1.6 穗颈瘟抗性鉴定
室内离体接种鉴定:待抗性鉴定菌系确定后,将供试的10个不同抗性水平的水稻材料在大田正常栽培,待长至孕穗破口初期,从田间每一品种的小区内剪取20个穗长露出1~2 cm的稻穗,带回实验室,剪取长约7~8 cm长的穗段(穗颈节上部2 cm,下部5~6 cm),然后将4根穗段放入铺有双层过滤纸的培养皿内(每皿加2 mL的无菌水),用移液器在每根穗段的穗颈节位点及节下约4 cm处各滴入5 µL的孢子悬浮液。将培养皿放入26 ℃的光照培养箱中,并用透明的薄膜包被,以保持培养皿内的湿度。光暗交替培养10~12 d。离体接种培养12 d后,检查各培养皿穗颈的发病情况,分级标准如下:0级:无病斑;1级:病斑长度不超过5 mm;3级:病斑长度6~15 mm;5级:病斑长度超过16 mm,但病斑间不相连;7级:病斑相互连接,大部分穗段发病;9级:病斑相互连接,80%以上穗段发病。将平均病级小于1的定为抗病;在1~3之间的定为中抗;在3~5之间的定为中感;在5~7之间的定为感病;大于7的定为高感。抗性频率(%)=(抗病菌株数/总菌株数)×100%。
自然病圃鉴定:将10个供试水稻品种按正常田间操作种植在抗性鉴定菌系来源地区大田,在种植过程中不施杀菌剂,待长至半黄熟期时随机调查每个品种的31个穗,计算平均病级(按国家稻瘟病穗颈瘟分级标准调查)以及病穗率。病穗率(%)=(发病穗数/总穗数)×100%。
2 结果与分析
2.1 稻瘟病菌毒性相关基因分子检测结果
采用16对特异性引物标记检测包括8个稻瘟病菌无毒基因与8个致病基因共16个稻瘟病毒性相关基因在供试的189个稻瘟病菌中的分布情况,并计算每个基因在所有菌株中的检测频率。检测结果(图2、图3)显示,16个稻瘟病菌毒性相关基因在江西稻瘟病菌中的检测频率为0%~96.29%,存在显著差异,其中最高检测频率的基因是MPG1,为96.29%,最低的是MNH1,为0%。结果表明江西稻瘟病菌遗传结构较为复杂,存在多种不同的致病类型。
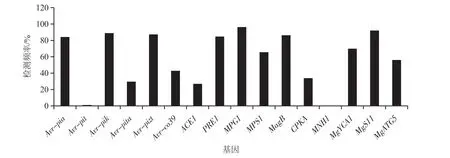
图2 稻瘟病菌毒性相关基因的分子检测频率
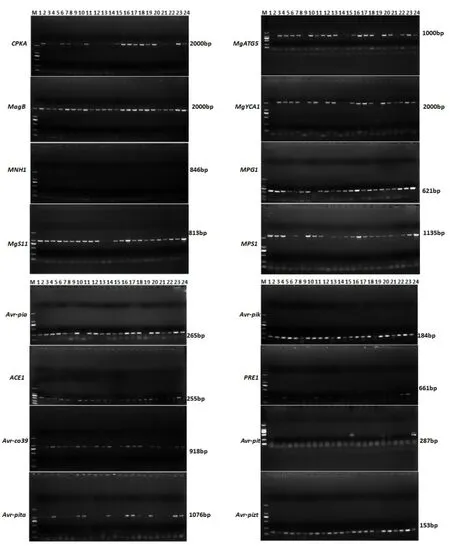
图3 部分稻瘟病菌无毒基因与致病基因的分子检测结果
2.2 稻瘟病菌毒性相关基因分子测定结果聚类分析
根据 PCR 扩增条带的结果构建基于SSR的“0-1”数据库,用最长距离法对菌株群体遗传多样性进行聚类分析。通过聚类结果(图4)可知,不同的相似系数可以将江西稻瘟病菌划分成不同的遗传宗谱单元,本研究确定在相似系数0.75时供试的稻瘟病菌株划分成的遗传宗谱数较为合理,聚类结果显示,在相似系数0.75时供试的189个稻瘟病菌株被聚类成7个遗传宗谱(表2)。其中优势宗谱为L1与L4,菌株占比分别为21.69%与63.49%;其他5个宗谱为小宗谱或稀有宗谱。

图4 稻瘟病菌致病基因与无毒基因分子检测结果的聚类结果

表2 遗传相似系数0.75水平下稻瘟病菌的遗传宗谱聚类结果
2.3 稻瘟病抗性鉴定菌系的确定
根据上述聚类结果,初步确定稻瘟病抗性鉴定菌系总菌株数为20个,挑选菌株的原则:将每个宗谱的稻瘟病菌株占所有菌株的百分比作为菌系中各宗谱菌株所占的百分比,随机挑选菌株;如果宗谱总菌株数与百分比的乘积小于1,则只挑选1个作为鉴定菌株;其他宗谱按照宗谱菌株数与其占比确定筛选鉴定菌株数。本研究确定的20个稻瘟病菌株组成的抗性鉴定菌系包括L1宗谱3个、L2宗谱1个、L3宗谱1个、L4宗谱12个、L5宗谱1个、L6宗谱1个、L7宗谱1个(表3)。
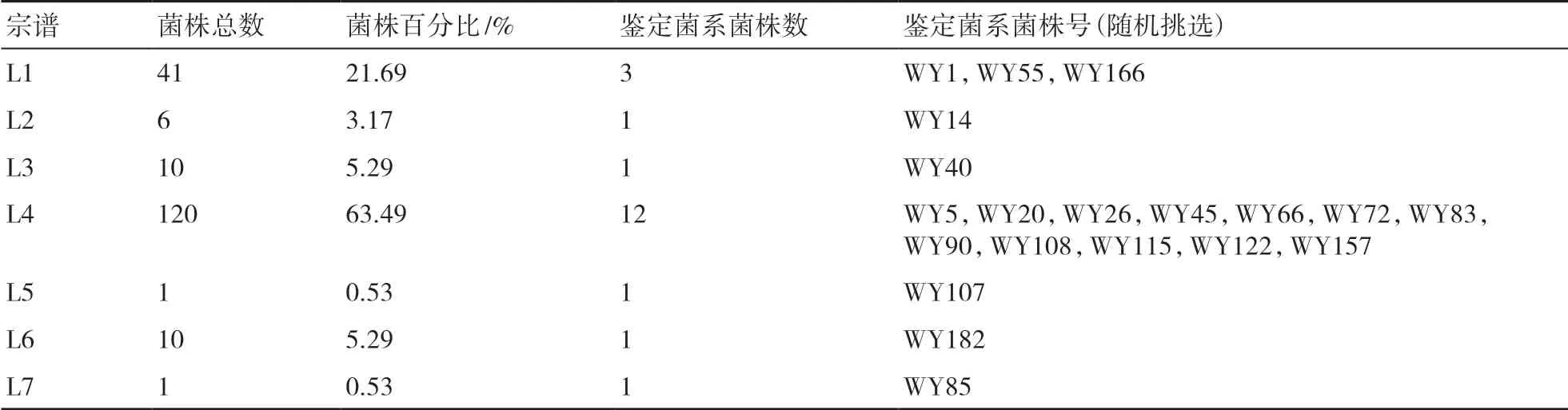
表3 稻瘟病抗性鉴定菌系的筛选结果
2.4 抗性鉴定菌系室内穗颈瘟抗性鉴定结果验证
利用筛选到的20个抗性鉴定菌系,在室内采用离体穗段点滴接种法与田间自然病圃诱发鉴定2种方法对10个水稻品种进行穗颈瘟抗性鉴定。结果(表4、图5)显示:源丰A、昌A、泰香A、莲CS、昌香A、33S靓占、望S靓占、两优靓占、九香粘、唐S靓占这10个供试品种利用本研究筛选的抗性鉴定菌系鉴定穗颈瘟的结果分别为中感、感病、感病、中抗、抗病、高感、中感、感病、高感、抗病,与田间自然诱发鉴定结果的表现完全一致。鉴定结果表明利用本研究筛选的稻瘟病鉴定菌系对水稻品种的穗颈瘟抗性鉴定准确度高。

图5 抗性鉴定菌系室内离体穗颈瘟抗性鉴定结果

表4 抗性鉴定菌系室内鉴定与田间自然诱发鉴定的结果比对验证
3 讨论
利用抗病品种防治稻瘟病是目前最经济有效的方法,但随着抗病品种大面积的持续推广,田间稻瘟病菌生理小种在寄主品种定向选择的压力下会不断变异产生新的致病小种,从而致使抗病品种的抗性丧失。另一方面,单一水稻品种的大面积连续推广也会改变当地的生理小种结构以及优势生理小种种群,因此为克服稻瘟病菌生理小种变异带来的病害流行,需要不断筛选新的抗病品种并加以推广应用。水稻品种的推广应用、稻种遗传资源的评价、抗病基因的发掘和利用等均需要一套长期稳定且具有代表性的稻瘟病菌鉴别体系。
室内人工稻瘟病抗性鉴定相较于自然病圃抗性鉴定具有多方面的优点,目前多采用水稻苗瘟期鉴定。而用于抗性鉴定的菌株筛选最早是通过接种鉴别品系的抗感反应来划分类群,但这种方法由于受接种环境与操作人员素质的影响较大,结果会出现一定的偏差。本研究中的方法是基于稻瘟病菌本身遗传基因水平上的差异,通过特异性分子标记技术测定与稻瘟病菌毒性相关的一些基因的存在或缺失,根据测定结果进行聚类,从中筛选一套与当地稻瘟病菌群组成结构一致的代表性鉴定菌系并应用于水稻品种的稻瘟病抗性鉴定。从实际应用效果来看,从江西省5个具有代表性的不同地理生态型病圃分离筛选20个菌株组成的江西稻瘟病抗性鉴定菌系,在室内离体接种穗颈段测定当地10个抗性不同的水稻种质资源品种的穗颈瘟抗性水平,其结果与田间自然诱发鉴定的结果基本一致,抗性水平的划分也完全属于同一级别。因此,本研究基于稻瘟病菌毒性相关基因筛选的稻瘟病抗性鉴定菌系是可行的,其鉴定准确度高,在稻瘟病抗病育种与抗病品种筛选等方面应用前景广阔。
菌株的来源是稻瘟病抗性鉴定代表菌系构建成功与否的关键。本研究是从江西省5个代表性地区的稻瘟病自然病圃采集发病标本分离菌株,通过分子测定毒性相关基因筛选稻瘟病抗性鉴定菌系。然而由于稻瘟病菌的致病性分化严重以及易变性的特点,在后续研究中需要从更多地区采集标本分离菌株,不断更新和补充已筛选构建的稻瘟病抗性鉴定菌系,从而保证抗性鉴定结果的准确性。
