黄土丘陵区草地群落多样性和优势种养分对氮添加的响应
2022-11-04徐伟洲赖帅彬张艳珍亢福仁徐炳成
徐伟洲, 赖帅彬, 史 雷, 张艳珍, 亢福仁, 徐炳成,3
(1.榆林学院生命科学学院, 陕西 榆林 719000; 2.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨陵 712100; 3.中国科学院水利部水土保持研究所, 陕西 杨陵 712100)
草地是黄土丘陵区陆地生态系统的重要组成部分,具有生物多样性保护、水文调节、水土保持、碳固存以及畜牧业发展等多种生态服务功能[1]。近年来,矿物燃烧和农业氮肥滥用导致大气氮沉降递增[2]。据估算,2050年全球的大气氮沉降将达到195 Tg·a-1,且高氮沉降区正逐渐由东南向西北蔓延[3]。持续氮沉降对草地生态系统产生了多重影响,不同来源的氮输入(施肥或大气沉降)提高土壤氮有效性,是促进草地生产力的有效途径[4]。诸多研究指出,氮提高净初级生产力是以降低物种多样性为代价[5-6]。群落物种多样性是维持草地多种功能和过程的关键,不仅对生产力有积极影响,还能减缓土壤侵蚀、抵抗物种入侵和调节有害生物[7]。然而随着研究深入,发现外源养分添加会加大物种间氮素利用策略的差异,且加剧其对照资源的不对称性竞争进而引起物种多样性丧失[8-10]。草地生态系统初级生产力在很大程度上受到氮(N)和磷(P)的有效性的限制,土壤养分有效性和植物养分含量会随生物多样性的变化而改变[11]。因此,研究氮添加下草地群落物种多样性及其物种养分利用策略的响应差异,有助于准确掌握不同植物生长的氮适应策略,以达到平衡养分富集下植物生产力提高与多样性损失。
不同物种对氮的需求及其敏感性存在明显差异,物种间养分含量反映了不同物种氮需求量、获取方式和利用效率的种间竞争关系,受物种功能特性和土壤氮有效性的共同调控[12]。适当氮添加可显著增加禾本科植物地上生物量,而过量氮添加则会对其生长产生抑制;相比豆科植物,禾本科植物对氮添加的响应更敏感[13-15]。研究表明,氮磷含量较低的物种由于其对氮磷需求量低,反而在氮限制环境中更容易成为优势种,氮磷比高的物种通常在磷限制环境中容易占据优势[16]。植物氮磷比与土壤养分状况密切相关,成为判断植物氮磷限制水平和土壤营养状况的重要指标。Güsewell等[17]认为当植物氮磷比小于10或大于20时,植物分别受到明显氮限制和磷限制,而氮磷比在10~20范围时将和养分限制没有明确关系。宋彦涛等[12]研究发现松嫩草地草本植物氮磷比为13.0,叶片氮含量低于我国其他草原区,叶片磷含量高于或等于其他草原区,综合判断为氮元素限制。氮添加增加了土壤有效氮含量,同时也引起土壤pH值和全磷含量降低,氮富集可促进了土壤磷循环且影响植物对N,P等营养元素的吸收和利用[18-19]。因此,结合植物养分含量与土壤养分含量状况,评估外源氮添加下草地群落物种多样性的响应规律具有重要意义。
优势种对草地群落结构和功能的维持起着至关重要的作用,植被对外源养分输入的响应主要取决于优势种的响应和适应能力[20]。白羊草(Bothriochloaischaemum)和达乌里胡枝子(Lespedezadavurica)在陕北黄土丘陵区退耕草地群落中均具有较广的分布幅度和占比[21],其中白羊草为多年生禾本科植物,具有适应性强和耐践踏等特性,属该区草地群落的建群种和优势种[22]。达乌里胡枝子为豆科多年生半灌木,是优良的水土保持植物,属该区草地群落中的伴生种[23]。因此,本研究选择分别以白羊草和达乌里胡枝子为优势种的草地群落为对象,通过设置不同N添加量处理,研究2种优势物种生物量特征和氮磷含量,群落土壤氮磷含量及群落物种多样性的变化规律,为揭示半干旱区草地对氮沉降加剧的响应以及采用施氮调控草地群落提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市安塞区纸坊沟小流域(109°19′23″E,36°51′31″N),流域总面积8.27 km2,海拔为1 010~1 431 m,气候属于典型的温带大陆性半干旱季风气候。年平均气温8.8℃,年均日照时数为2 395.6 h,年均无霜期160 d,年总辐射量为493 KJ·cm-2,年蒸发量1 500~1 900 mm,多年平均降雨量507.2 mm(1954—2018年),且季节分配不均,7—9月降雨量占60%~80%。该区地形地貌复杂多样,境内沟壑纵横,土壤类型以黄绵土为主,植被类型属于暖温带落叶阔叶林区向草原区过渡带,典型地带性草本植物有白羊草、达乌里胡枝子、长芒草(Stipabungeana)、铁杆蒿(Heteropappusaltaicus)、茭蒿(Artemisiagiraldii)等。
1.2 试验设计
在纸坊沟小流域内,选择分别以白羊草为优势种,达乌里胡枝子为伴生种,以及达乌里胡枝子为优势种,白羊草为伴生种的2个群落作为试验样地,坡度为8°,坡向为东偏西46°。采用随机区组设计,沿坡面设置3个2 m×6 m的样带,每个样带为一个重复,间距为1 m,各样带划分为3个2 m×2 m的样地。根据黄土丘陵区年均氮沉降量(21.76 kg·hm-2·a-1),设置对照、1倍和3倍年均模拟氮沉降水平,即N0(不施N,0 kg·hm-2·a-1)、N25(25 kg·hm-2·a-1)和N75(75 kg·hm-2·a-1)处理,每个草地群落9个样地,总计18个样地。氮肥为硝酸铵钙[5Ca(NO3)2·NH4·NO3·10H2O],于2018年2月将地表所有枯枝落叶移除,在雨季来临前的5月21日,将提前称好的颗粒状氮肥均匀撒入对应试验样地。
1.3 测定指标及方法
1.3.1植物群落调查 于2018年9月中旬,在每个2 m×2 m的样地中设置1个1 m×1 m样方进行调查,以避免边缘效应的影响。记录样方内出现的物种、个体数量,随后各物种随机选择5株个体用卷尺测量其株高。将1 m×1 m样方分成4个0.5 m×0.5 m的小样方,用目测法估算各物种分种盖度和群落总盖度。调查后将样方内植物地上部分用剪刀齐地刈割后,将白羊草、达乌里胡枝子单独分装入信封袋,其余物种一起装入1个信封袋,105℃杀青30 min后,85℃的烘箱中烘干至恒重,获得白羊草和达乌里胡枝子地上生物量,加上其余物种获得群落总地上生物量(0.01 g·m-2)。
1.3.2物种多样性指数 群落物种组成采用各物种重要值(Importance value,IV)表征,群落α多样性指数采用Shannon-Winner多样性指数(H′,)、Pielou均匀度指数(Pielou evenness index,J)和Simpson优势度指数(Simpson diversity index,D)共同表征。
IV=(相对高度+相对盖度+相对生物量)×100/3
(1)
(2)
J=H/InS
(3)
(4)
式中:S为物种总数,相对盖度为某一物种盖度与全部物种盖度之和的比值,相对高度为某一物种高度与全部物种高度之和的比值,相对生物量为某一物种生物量干重与全部物种生物量干重之和的比值,Pi为某一物种的重要值与全部物种重要值之和的比值。
1.3.3土壤采集 草地群落调查测定同期,在上述每个样方内用内径3 cm的土钻取0~10 cm,10~20 cm,20~30 cm土层土壤,各样方随机重复3次,各层3重复样品混合。土壤样品带回实验室后自然风干,通过0.25 mm筛孔筛分用于测定土壤全氮和全磷。
1.3.4氮磷含量分析 将烘干后的不同物种地上部分植物样品分类粉碎过0.15 mm筛,用于测定白羊草、达乌里胡枝子等优势种的全氮和全磷含量。土壤和植物样品的全氮量测定采用全自动凯氏定氮仪(FOSS-8400,丹麦),其全磷含量测定采用H2SO4-HClO4消煮后采用钼蓝比色法(UV-2600紫外可见分光光度计,日本)。
1.4 数据分析
同一指标(株高、盖度、地上生物量、植物全氮、全磷及氮磷比、Shannon多样性、Pielou均匀度、Simpson优势度、土壤全氮、全磷含量及氮磷比)在不同处理间(N0,N25,N75)差异显著性采用单因素方差分析(One-way ANOVA,LSD)进行检验(P<0.05);同一指标在不同群落间(Bi群落,Ld群落)差异采用独立样本t检验(Independent sample t-test)(P<0.05)。上述分析在SPSS 20.0中进行,相关图的绘制在Sigmaplot 14.0中完成。采用Pearson相关性分析方法探究分析优势种特征、优势种养分含量,群落物种多样性以及土壤养分含量间的相关关系。将2个群落中的白羊草特征指标合并,达乌里胡枝子特征指标合并,得到优势种水平的特征指标(株高、盖度、生物量、全氮、全磷、氮磷比),将2个群落的物种多样性指标和土壤养分含量会标合并,得到群落水平的特征指标,形成1个18(样本)×18(指标)的矩阵数据,应用R语言中的“cor”函数及ggplot2包进行相关性分析与绘图。
2 结果与分析
2.1 株高、盖度和生物量
由图1可知,氮添加对Bi群落中白羊草和达乌里胡枝子的株高无显著影响,但N75处理显著提高Ld群落中白羊草的株高(P<0.05)。Bi群落中,氮添加对白羊草的盖度无显著影响,N25和N75处理下达乌里胡枝子的盖度较N0处理分别降低了76%和53%;N25和N75处理下白羊草的地上生物量较N0处理分别增加28%和78%,但达乌里胡枝子的地上生物量,分别降低了88%和48%(P<0.05)。
Ld群落中,氮添加对达乌里胡枝子的盖度无显著影响,白羊草的盖度随着氮添加量的增多而提高,由N0的3%增加到N75的25%,且白羊草的盖度均小于达乌里胡枝子;N75处理显著降低达乌里胡枝子的地上生物量,较N0处理降低了33%,但N25与N0处理间无显著差异;N25和N75处理均显著提高了白羊草的地上生物量,即分别提高了136%和460%(P<0.05)。
相较于N0处理,N25处理均显著提高了Bi群落和Ld群落总地上生物量,N75处理显著提高了Ld群落总地上生物量(P<0.05,图2),而Bi群落中N75处理与N0和N25处理间均无显著差异。相同氮处理下,Bi群落和Ld群落总地上生物量相似,N0处理地上总生物量分别为181 g·m-2和141 g·m-2,N25处理下分别为247 g·m-2和252 g·m-2,N75处理下分别为247 g·m-2和218 g·m-2。Bi群落中,氮添加后群落总地上生物量增加主要为白羊草的地上生物量增加,而达乌里胡枝子和其他物种均无显著差异;Ld群落中,群落总生物量增加主要为白羊草和其他物种的地上生物量增加,而达乌里胡枝子呈下降趋势。

图2 2种典型草地群落不同物种生物量贡献与总地上生物量对氮添加的响应Fig.2 Biomass contributions of different species and the responses of total aboveground biomass to N addition in two typical grassland communities
2.2 物种多样性
单因素方差分析结果表明,氮添加对Bi群落和Ld群落的3个物种多样性指标(Shannon多样性指数、Pielou均匀度指数和Simpson优势度指数)均产生显著影响(P<0.05)(图3)。Bi群落中,氮添加显著降低了Shannon多样性、Pielou均匀度和Simpson优势度指数,N75处理下均显著最低(1.51,0.73和0.66),N0处理下均显著最高(2.1,0.88和0.86)(P<0.05)。Ld群落中,氮添加显著增加了Shannon多样性、Pielou均匀度和Simpson优势度指数,N75处理下均显著最高(2.44,0.91和0.89),N0处理下显著最低(2.02,0.85和0.83)(P<0.05),且N25与N75处理间均无显著差异。氮添加后,N25和N75处理下Bi群落的3种多样性指数均低于Ld群落。
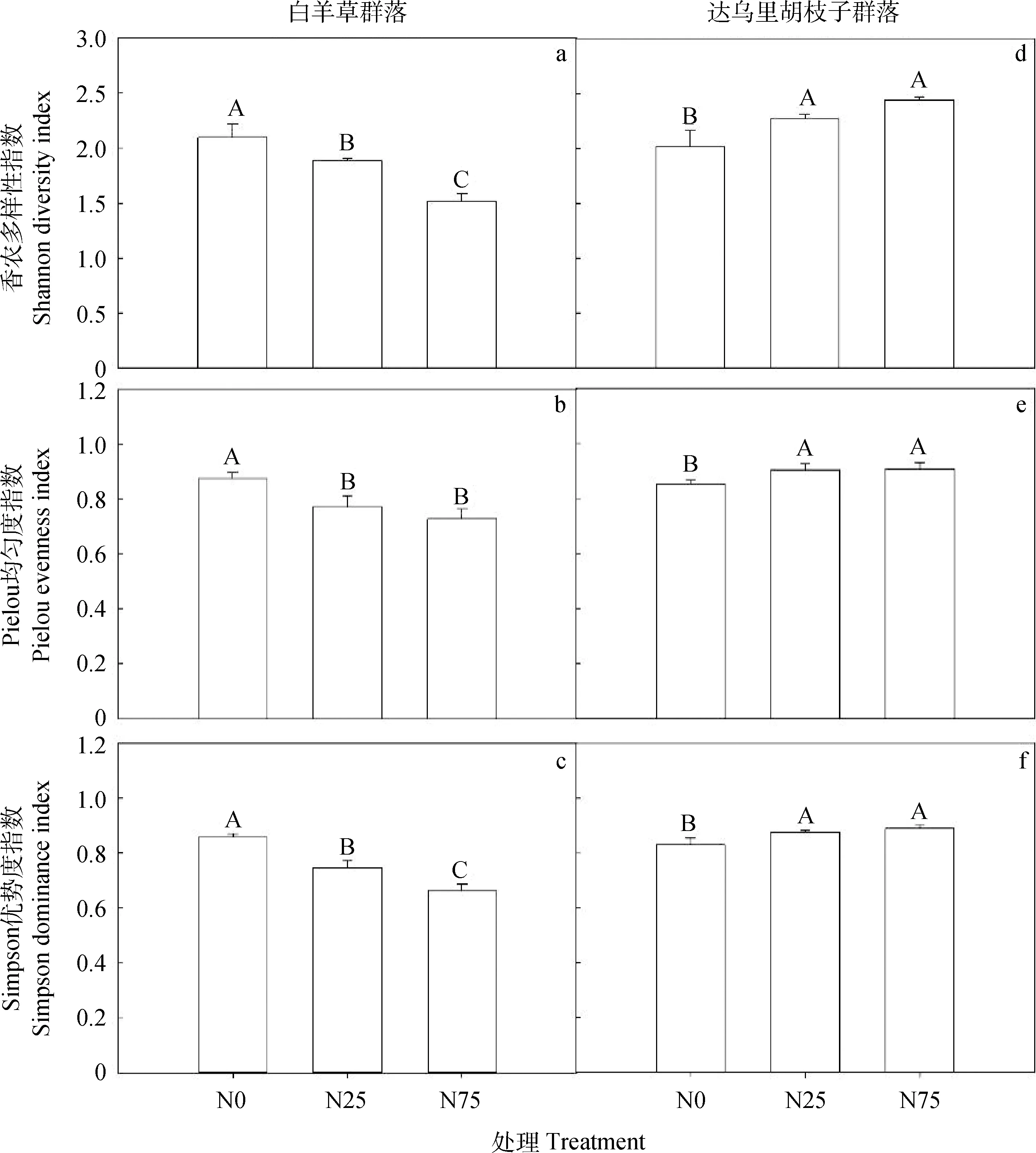
图3 2种典型草地群落的物种多样性指数对氮添加的响应Fig.3 Species diversity indices in response to N addition in two typical grassland communities
2.3 优势种植物氮磷含量
由图4可知,达乌里胡枝子的全氮含量和氮磷比均高于白羊草。N0处理下,Bi群落中白羊草的全氮含量显著低于Ld群落(P<0.05);N25或N75处理下,白羊草的全氮含量在Bi和Ld群落间均无显著差异;不同氮添加处理下,达乌里胡枝子的全氮含量在Ld群落中均显著高于Bi群落;达乌里胡枝子的全磷含量在Ld群落和Bi群落间均无显著差异,而N25与N75处理下Ld群落中白羊草的全磷含量均显著低于Bi群落(P<0.05)。
Bi群落中,氮添加显著提高了白羊草的全氮含量,由N0的7.84 g·kg-1分别增加到N25的9.49 g·kg-1和N75的10.81 g·kg-1;N75处理下白羊草的全磷含量较N0处理显著降低了32%,N25与N0间无显著差异(P<0.05);白羊草的氮磷比分别由N0的5.48增加到N25的7.29和N75的11.05;N25和N75处理下达乌里胡枝子的全氮、全磷及氮磷比均与N0处理间无显著差异。
Ld群落中,氮添加显著降低达乌里胡枝子的全氮含量及氮磷比,全氮含量由N0的24.37 g·kg-1分别降低为N25和N75的19.64 g·kg-1和20.28 g·kg-1;氮磷比由N0的24.54 g·kg-1分别降低为N25和N75的17.40 g·kg-1和18.08 g·kg-1;N25和N75处理下白羊草的全磷含量分别较N0处理降低了23%和44%,而白羊草氮磷比在N25和N75处理下分别提高了33%和88%。
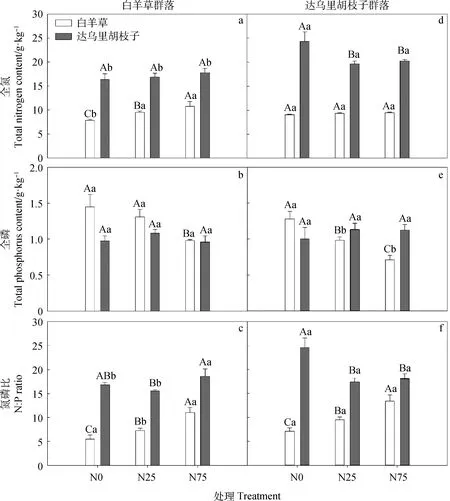
图4 2种典型草地群落中白羊草和达乌里胡枝子的全氮、全磷含量和氮磷比对氮添加的响应Fig.4 Total nitrogen and phosphorus contents and N∶P ratio of B.ischaemum and L.davurica in responses to N addition in two typical grassland communities
2.4 土壤氮磷含量
Bi群落中,0~10 cm及10~20 cm土层的全氮含量在不同氮添加下无显著差异,20~30 cm土层的全氮含量以N75处理显著低于N25和N0处理(P<0.05,图5);N0和N75处理下,20~30 cm土层的全磷含量显著低于0~10 cm土层(P<0.05);N0处理下,土壤氮磷比随土层深度增加显著增加;N25处理下,10~20 cm土层的氮磷比显著低于N0和N75处理;N75处理下,20~30 cm土层的氮磷比显著低于N25和N0处理(P<0.05)。
Ld群落中,同一氮处理下,全氮含量和氮磷比均随土层深度的增加而显著降低(P<0.05);N25处理下,20~30 cm土层的全磷含量显著低于0~10 cm;0~10 cm土层,不同氮添加处理下的全氮和全磷含量均无显著差异,而氮磷比以N25处理显著低于N0和N75处理(P<0.05);10~20 cm土层,N25处理的全氮含量显著低于N75和N0处理,N75处理的氮磷比显著高于N25处理,而其全磷含量在不同氮处理间无显著差异;20~30 cm土层,全氮含量和氮磷比均为N25处理显著最低且N75处理显著最高(P<0.05),N0和N75处理的全磷含量无显著差异。
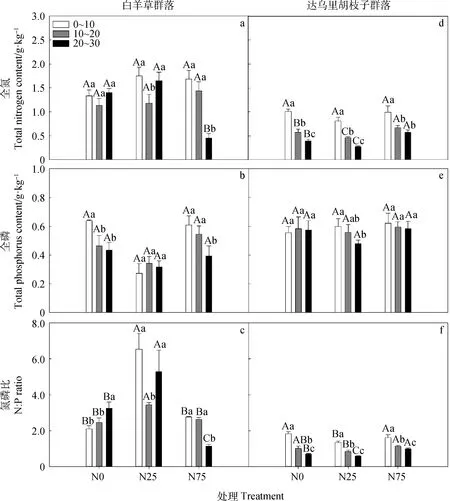
图5 2种典型草地群落土壤全氮、全磷及氮磷比对氮添加的响应Fig.5 Soil total nitrogen,total phosphorus and N:P ratio in responses to N addition in two typical grassland communities注:不同大写字母表同一土层不同氮处理间差异显著(P<0.05);不同小写字母表示同一氮处理下不同土层间差异显著(P<0.05)Note:Different capital letters indicate significant differences among N treatments in the same soil layer at the 0.05 level,and different lowercase letters indicate significant differences among different soil layers under the same N treatment at the 0.05 level
2.5 物种多样性与优势物种特征的关系
Pearson相关性分析结果表明,Shannon多样性指数、Pielou均匀度指数和Simpson优势度指数与白羊草的盖度和生物量存在显著负相关关系(-0.82
白羊草的养分含量与土壤养分无显著相关关系,而达乌里胡枝子的全氮含量和氮磷比与土壤总氮含量和土壤氮磷比呈显著负相关关系(R2<0,P<0.05);达乌里胡枝子的全磷含量与土壤全磷含量呈显著正相关关系(R2>0,P<0.05)。土壤全氮含量与全磷呈极显著负相关关系(R2=-0.74,P<0.001),与氮磷比呈极显著正相关关系(R2=0.90,P<0.001),土壤全磷含量与氮磷比呈极显著负相关关系(R2=-0.94,P<0.001)。
3 讨论
3.1 2种优势种生物量对氮素添加的响应
白羊草和达乌里胡枝子分别作为优势种和伴生物种时,其个体生长对氮素添加的响应存在显著差异。在白羊草群落中,氮添加显著提高了白羊草地上生物量及群落总生物量;在白羊草作为伴生种的达乌里胡枝子群落中,氮添加同样显著提高了白羊草地上生物量,表明氮添加促进禾本科植物的生物量积累的普遍规律不受群落中其他物种影响。有研究认为,氮素添加显著增加了禾草类生物量及其生物量占比,本研究结论与其一致[24]。这是由于白羊草属多年生无性系物种,庞大的须根系可增大其对土壤养分的吸收面积,且其株高均大于达乌里胡枝子,即通过快速生长和光合产物增加而获得相对较大的植株个体[5,16]。氮添加对豆科植物达乌里胡枝子的株高和盖度无显著影响,但N75处理下的地上生物量显著降低,且其对群落总生物量的贡献量显著降低,而白羊草的地上生物量占比显著增加,表明达乌里胡枝子作为优势种或非优势种时,N75处理(75 kg·hm-2·a-1)已超出达乌里胡枝子的可利用氮阈值,且过量氮添加抑制了其生物量积累。氮对豆科植物的抑制作用已被研究证实,豆科植物因其自身具有固氮能力可在缺氮环境中成为优势种,但当土壤有效氮含量增加时,导致其他物种对氮素的竞争加剧[25-26];且达乌里胡枝子个体较低矮,杂类草等更容易抢占冠层上部光资源,阻碍了达乌里胡枝子的光能利用效率[5]。相关性分析结果显示,白羊草的盖度和地上生物量与达乌里胡枝子呈极显著负相关,表明2种植物在群落中存在着明显的竞争拮抗作用,而白羊草的竞争优势高于达乌里胡枝子。
3.2 群落物种多样性对氮添加的响应
白羊草群落的物种多样性随氮添加量增加而显著降低,而达乌里胡枝子群落则略有增加,这进一步说明不同优势种群落对氮添加的响应存在差异,体现了物种间性状互补和生态位分离。白羊草群落的物种多样性与生产力存在显著负相关关系,证实了氮添加对群落多样性的负向影响与对生产力的正向影响是同时存在的,筛选出最适量的氮添加阈值进而平衡对草地的积极和消极影响已成为研究热点[5,27]。一项Meta分析结果表明,典型草原氮添加阈值为17.3 g·m-2·a-1,远高于本研究设置的氮添加量[28];另一项Meta分析[27]结果显示,钙质草地氮沉降的临界负荷为15~25 kg·hm-2·a-1,与本研究中N25处理水平接近。相比对照,白羊草群落中3种多样性指数(Shannon多样性指数、Pielou均匀度指数和Simpson优势度指数)随着氮添加量的增加而显著降低;达乌里胡枝子群落中N25和N75处理间的多样性指数无显著差异,表明N25处理可能为该区草地群落较适合的氮添加量,此添加量下可权衡2种草地群落生产力增加与多样性下降。本研究发现,白羊草群落的盖度与地上生物量随氮添加增加而显著增加,白羊草地上生物量占比也随之显著增加,其群落总地上生物量的贡献量远高于其他物种之和(图2),且在达乌里胡枝子群落中N75处理下生物量的占比同样显著增加。凋落物竞争假说可以解释禾草群落物种多样性下降机制,研究表明地上生物量增加会导致更多的凋落物积累,阻碍幼苗生长发育或降低植物的光合有效辐射,可能是造成物种多样性降低的重要原因[29]。达乌里胡枝子群落的物种多样性在氮添加后显著增加,3种多样性指数(Shannon多样性指数、Pielou均匀度指数和Simpson优势度指数)均与达乌里胡枝子株高、盖度、生物量呈显著正相关关系。Seabloom等[25]研究发现未施肥草地多样性的增加主要来源于物种均匀度的增加,并不是由于物种间的竞争和排斥所驱动。本研究中,达乌里胡枝子群落中达乌里胡枝子的优势地位在氮添加后降低,进而空出更多的生态位而有利于其他非优势种植物生长[30],其他物种的均匀度增加可能是物种多样性增加的主要原因。白羊草群落和达乌里胡枝子群落的物种多样性对氮素添加呈完全相反的响应规律,“凋落物假说”和“生态位假说”可以解释这2种群落不同的响应机制,表明优势种功能性状决定着草地群落功能的变化,且优势种差异与群落响应间的层级联系需更多深入的研究。
3.3 优势种植物氮磷含量对氮添加的响应
优势种植物氮磷含量及氮磷比的变化反映了自身的生理代谢状况,是植物资源利用策略的体现。不同氮添加处理下2种草地群落中达乌里胡枝子的全氮含量与氮磷比均显著高于白羊草,表明豆科植物拥有更高的氮吸收利用能力,这归因于豆科植物根瘤菌具有固氮作用,因此在缺氮环境中更具优势[31]。白羊草的全氮含量随着氮添加量的增加而显著增加,但其全磷含量逐渐降低,氮磷比由5.48和7.13分别提高为11.05和13.39,该变化趋势在白羊草为优势种和伴生种的群落中均相似,表明氮添加促进了白羊草对氮素利用能力且加剧了其对磷的需求,白羊草逐渐由氮限制转变为磷限制。研究认为,植物的N∶P<14.0或10.0时受到明显的氮限制,N∶P>14.0或20.0时受到明显的磷限制[32-33]。白羊草全氮与土壤全氮无显著相关关系,植物全磷与土壤全磷呈负相关关系,N25处理下土壤全磷含量显著最低,可能是因为氮诱导的植物和细菌生物量积累增加了其对速效磷的需求,从而导致磷循环的大幅上调而进一步加剧磷限制[33]。白羊草的氮磷含量在2个群落间无显著差异,具有相对稳定的养分利用状况,全氮含量随氮添加显著增加,表明氮添加提高了白羊草的氮吸收利用能力,从而促进其地上生物量的积累。氮添加对白羊草群落中的达乌里胡枝子全氮、全磷及氮磷比无显著影响,而在达乌里胡枝子群落中显著降低其全氮含量及氮磷比,且N75下达乌里胡枝子生物量显著降低,表明氮添加对豆科达乌里胡枝子产生抑制作用。豆科植物对短期氮添加的响应表现为减少根瘤的形成、固氮效率及固氮量均明显降低[34]。达乌里胡枝子的氮磷比在2种群落中均高于14.0,说明其长期受到磷限制[16];达乌里胡枝子群落中,氮添加后达乌里胡枝子的氮磷比有所下降,说明氮添加缓解了部分磷限制但不足以改变磷限制的现状,可通过对豆科草地群落进行氮磷添加或配施以促进群落稳定。
3.4 优势种地上生物量与物种多样性关系
氮添加促进了白羊草和达乌里胡枝子群落的生产力,但多样性的响应呈现相反的变化趋势,不同优势种群落生产力与多样性间关系存在着不同的响应机制。生物量为资源数量的代表,植被N∶P为资源供应的代表,有学者研究发现在N∶P比例平衡的条件下植物的丰富度最高[29]。本研究未发现植物氮磷比与多样性间存在相关关系,优势物种生产力对群落多样性的影响更加强烈。群落多样性响应一般分为物种的更替或种间丰度变化,优势种生产力在调节群落多样性方面起着重要作用[35]。氮添加后白羊草生产力增加且达乌里胡枝子下降,物种多样性与白羊草生产力呈负相关,与达乌里胡枝子生产力呈正相关,白羊草生产力增加将不利于多样性的维持,而达乌里胡枝子生产力占比下降有利于多样性维持。白羊草属多年生无性系禾草,氮添加未改变其株高与盖度,但提高了地上生物量,表明白羊草的氮素利用效率显著增加[36]。更高的养分利用效率将会延长养分在植物中的停留时间且增加了衰老植物对养分的重吸收能力,可促进在第二年生长中快速占据绝大部分生产资料[37-38]。白羊草较高的株高阻碍了低矮植物对光的吸收,因此在一定程度上抑制了群落多样性,可能造成了物种多样性的损失[39]。达乌里胡枝子群落中土壤全氮、全磷含量及氮磷比均低于白羊草群落,资源互补效应是群落中生产力与多样性之间存在正相关关系的主要机制[40]。豆科植物达乌里胡枝子自身具有固氮能力(即使氮添加抑制其生长),额外氮添加削弱了土壤的氮限制条件,因此有利于非固氮植物(如白羊草、铁杆蒿和茭蒿等)与豆科植物互补利用添加后的土壤有效氮。且达乌里胡枝子的株高相对较矮,在光竞争中处于劣势,因此氮诱导其他物种生物量积累的情况下更易受到竞争抑制,其他学者研究同样发现了氮添加对豆科物种的抑制作用[41]。因此观察其他物种生产力占比显著增加,且物种多样性同样增加,多样性较高的群落中植物对营养资源利用更为充分,使得植被根系层土壤养分浓度降低[39],这进一步支持了本研究结果。
4 结论
本研究结果表明,以白羊草和达乌里胡枝子为优势种的2种草地群落对氮添加的响应存在显著差异。氮添加显著提高了白羊草的氮磷比,但显著降低了达乌里胡枝子的全氮含量及氮磷比。未发现白羊草和达乌里胡枝子的氮磷含量与物种多样性间的相关性,而优势种生物量是影响氮添加下群落物种多样性变化的主要因素。白羊草为优势种的群落,氮添加下地上生物量显著增加对群落多样性产生显著负效应,禾草优势种产生较强的竞争排斥作用不利于物种多样性。达乌里胡枝子群落在氮添加下物种多样性的提高得益于其他物种生物量占比的提高,物种间对氮素的互补利用是豆科群落多样性增加的主要原因。研究结果阐明了优势种对氮添加的响应与群落水平多样性的响应间的递进关系,优势种的生物量与生态位优势而非养分含量影响着群落物种多样性水平。
