汉黄芩苷干预高糖刺激RAW264.7巨噬细胞KLF4/NF-κB信号通路
2022-11-03徐兴欣
谢 姗,徐兴欣
糖尿病肾病(diabetic nephropathy, DN)是糖尿病最常见的微血管并发症,在DN发生和发展中巨噬细胞的参与起着至关重要的作用。研究显示,在DN肾组织中,过表达的Krüppel样因子4(Krüppel-like factor4,KLF4)可以抑制肾脏炎症,表现出抗炎、抗纤维化的保护作用[1-3],也可以在巨噬细胞炎症中进行调节[4-5]。因而,激活KLF4的表达可能是延缓DN进展的有效手段。汉黄芩苷(wogonoside, WG)属于生物活性黄酮,具有抗氧化、抗炎、抗病毒和抗癌的作用[6]。同时,WG能显著降低脂多糖(lipopolysaccharide, LPS)刺激引起的巨噬细胞炎症因子释放[7]。但是WG是否干预高糖诱导巨噬细胞活化,KLF4是否参与炎症及WG干预机制,目前尚未见报道。该研究以巨噬细胞为研究对象,通过观察WG对巨噬细胞中相关信号通路及炎症因子的影响,探讨 KLF4-蛋白核因子κB(nuclear factor κB, NF-κB)信号通路在高糖诱导巨噬细胞激活中的具体机制。
1 材料与方法
1.1 细胞与试剂小鼠巨噬细胞系(RAW264.7)购自中科院上海细胞生物学研究所。WG(Batch NO.:DST210409-026)购自成都乐天美医药科技有限公司。兔抗CD68、白介素1β(interleukin-1β,IL-1β)、肿瘤坏死因子α(tumor necrosis factor alpha,TNF-α)、诱生型一氧化氮合酶(inducible nitric oxide synthase, iNOS)、KLF4单克隆抗体购自美国Abcam公司,NF-κB p65、NF-κB pp65单克隆抗体购自美国CST公司,小鼠抗β-actin单克隆抗体、辣根过氧化物酶标记的羊抗兔IgG、羊抗小鼠IgG抗体购自武汉三鹰公司,小鼠TNF-α、IL-1β ELISA试剂盒购自美国R&D公司, DMEM低糖培养基、胎牛血清购自加拿大Wisent公司,D-葡萄糖、2-脱氧-D-葡萄糖(2-deoxy-D-glucose,2-DG)购自美国Sigma公司,BCA蛋白测定试剂盒、SDS-PAGE凝胶试剂盒购自江苏碧云天公司,TRIzol试剂购自雅酶公司,反转录试剂盒、实时荧光定量PCR试剂盒购自艾科瑞生物公司,ECL化学发光试剂盒购自美国Thermo Scientific公司。PCR引物由上海生工生物工程股份有限公司合成。
1.2 方法
1.2.1细胞培养 将RAW264.7细胞培养于含10%胎牛血清的低糖5.5 mmol/L DMEM培养基,放置于条件为37 ℃、5%CO2的培养箱。将细胞随机分为:LG组(5.5 mmol/L葡萄糖培养基)、D组(30 mmol/L甘露醇培养基)、HG组(30 mmol/L葡萄糖培养基)、LG+WG50组(5.5 mmol/L葡萄糖+50 μmol/L WG)、HG+WG12.5组(30 mmol/L葡萄糖+12.5 μmol/L WG)、HG+WG25组(30 mmol/L葡萄糖+25 μmol/L WG)、HG+WG50组(30 mmol/L葡萄糖+50 μmol/L WG)后同步化培养24 h,高糖刺激在WG预处理2 h后加入培养基,光镜观察各组细胞数量及形态变化,检查各组指标变化,并收集数据。
1.2.2细胞活力测定 将RAW264.7细胞置于96孔板中,于低糖培养条件下同步化24 h后,分别更换0、6.25、12.5、25、50、100 μmol/L WG高糖培养基,24 h后加入CCK-8孵育2 h,酶标仪450 nm波长下进行光密度(optical density,OD)值测定,并使用公式OD(450)实验组/OD(450)0 μmol/L×100%算出细胞活力。
1.2.3细胞荧光检测 将RAW264.7细胞以1×105个/孔的密度接种在玻璃皿底,4%多聚甲醛固定10 min,10%牛血清蛋白阻断30 min,加入iNOS一抗溶液4 ℃孵育过夜。PBS洗涤3次后,加入FITC标记的荧光二抗室温避光孵育1 h,DAPI染核5 min,加荧光抗淬灭剂封片后,在荧光显微镜下观察并采集图象,各组随机选取5~10个高倍镜视野,使用Image J软件计数细胞平均数。
1.2.4Western blot分析 收集各组细胞,加RIPA裂解液冰上裂解,BCA试剂盒测定蛋白浓度并计算上样量。适量蛋白通过SDS-PAGE凝胶进行电泳后转移到NC膜,用5%的脱脂牛奶封闭2 h,再用稀释一抗NF-κB p65(1 ∶1 000)、NF-κB pp65(1 ∶1 000)、KLF4(1 ∶1 000)、iNOS(1 ∶1 000)、TNF-α(1 ∶1 000)、IL-1β(1 ∶1 000)4 ℃孵育12 h,用HRP标记的山羊抗鼠或抗兔二抗(1 ∶8 000)进行孵育,TBST洗3次,滴加ECL显色剂在凝胶成像系统进行化学发光,并用Image J进行半定量分析。
1.2.5qRT-PCR TRIzol试剂裂解细胞后提取总RNA,进行RNA浓度及纯度鉴定,使OD260/280在1.8~2.1之间,并在反转录系统中逆转录为cDNA。依据SYBGREEN PCR试剂盒进行定量分析,终体积10 μl。引物序列见表1。每个样本设置3个复孔,设置GAPDH为内参对照进行校正,用2-△△Ct计算目的基因的相对表达量。

表1 引物序列
1.3 统计学处理所有数据使用SPSS 18.0软件进行统计学分析,方差齐性应用Levene法检验,两组间比较应用独立样本t检验,多组间比较应用单因素方差分析(One-way ANOVA),方差不齐应用K-W检验或Wilcoxon秩和检验,LSD分析数据以确定显著差异性。P<0.05表示差异有统计学意义。
2 结果
2.1 WG对RAW264.7细胞活性的影响WG选取0~100 μmol/L范围中6个浓度,见图1,当WG浓度在0~50 μmol/L时,高糖刺激后的RAW264.7细胞活性无明显影响(P>0.05),WG浓度高于50 μmol/L时,细胞活性明显下降(P<0.01)。
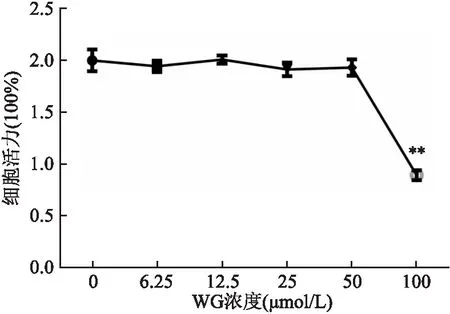
图1 CCK-8法检测WG对RAW264.7细胞活性的影响与0 μmol/L WG比较:**P<0.01
2.2 高糖刺激各组巨噬细胞iNOS的表达高糖刺激的巨噬细胞中iNOS表达量增高,阳性细胞数明显高于低糖培养的巨噬细胞(P<0.01),表明高糖可以促进巨噬细胞炎症。加入50 μmol/L WG预处理的高糖巨噬细胞,iNOS表达的阳性细胞数明显低于HG组,差异有统计学意义(P<0.01),见图2。
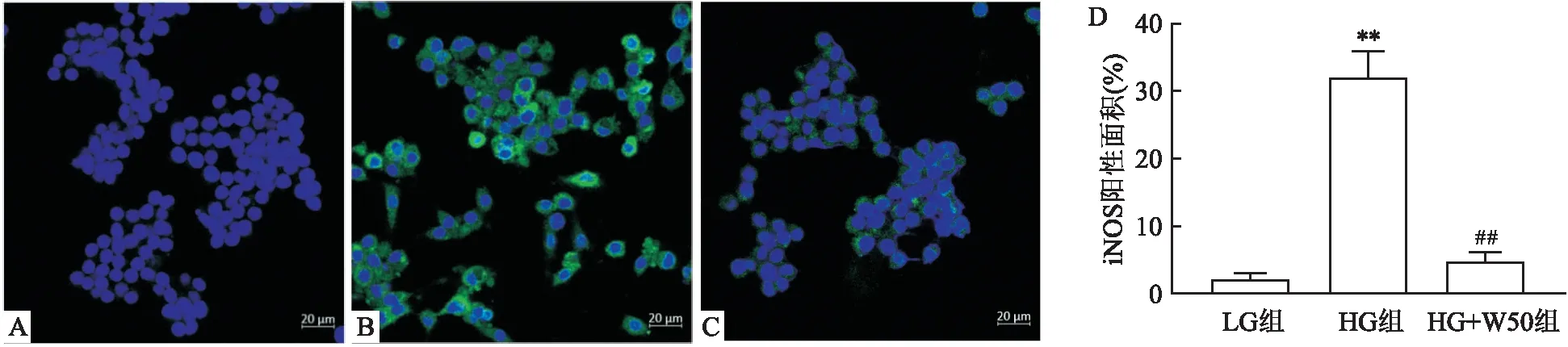
图2 LG组、HG组与HG+W50组iNOS的表达 ×400A:LG组;B:HG组;C:HG+W50组;D:荧光镜下iNOS阳性面积统计;与LG组比较:**P<0.01;与HG组比较:##P<0.01
2.3 WG减轻高糖刺激RAW264.7细胞引起的炎症Western blot、qRT-PCR结果显示,WG预处理2 h后,高糖刺激的RAW264.7细胞iNOS、TNF-α、IL-1β、NF-κB pp65表达均明显增高(P<0.01),WG明显抑制iNOS、TNF-α、IL-1β、NF-κB pp65表达,且50 μmol/L的抑制程度最明显,与HG组对比,差异有统计学意义(P<0.01),见图3~5。ELISA法检测各组细胞上清液,结果表明高糖刺激上清液中TNF-α、IL-1β表达增高,不同浓度的WG作用后TNF-α、IL-1β呈浓度依赖性降低(P<0.01),见图6。

图3 Western blot检测各组细胞中NF-κB p65、NF-κB pp65、KLF4、iNOS、TNF-α及IL-1β蛋白表达A:LG组、HG组、D组、LG+W50组的Western blot实验条带图;B:NF-κB p65、NF-κB pp65、KLF4、iNOS、TNF-α及IL-1β蛋白表达半定量统计;与LG组比较:**P<0.01;与HG组比较:##P<0.01

图4 Western blot检测各组细胞中NF-κB p65、NF-κB pp65、KLF4、iNOS、TNF-α及IL-1β蛋白表达A:LG组、HG组、HG+W12.5组、HG+W25组和HG+W50组的Western blot实验条带图;B:NF-κB p65、NF-κB pp65、KLF4、iNOS、TNF-α及IL-1β蛋白表达半定量统计;与LG组比较:**P<0.01;与HG组比较:#P<0.05,##P<0.01

图5 qRT-PCR检测各组细胞中KLF4、iNOS、TNF-α及IL-1β mRNA表达A: KLF4 mRNA表达;B iNOS mRNA表达;C:TNF-α mRNA表达;D: IL-1β mRNA表达;a:LG组;b:HG组;c:D组;d:LG+W50组;e:HG+W12.5组;f:HG+W25组;g:HG+W50组;与LG组比较:**P<0.01;与HG组比较:#P<0.05,##P<0.01

图6 各组细胞上清液中炎症因子TNF-α及IL-1β水平变化A:TNF-α表达水平;B:IL-1β表达水平;a:LG组;b:HG组;c:D组;d:LG+W50组;e:HG+W12.5组;f:HG+W25组;g:HG+W50组;与LG组比较:**P<0.01;与HG组比较:##P<0.01
2.4 WG干预高糖刺激RAW264.7细胞中KLF4的表达Western blot结果显示,高糖刺激后,巨噬细胞内KLF4蛋白表达量较LG组下降(P<0.01),在WG干预后,KLF4的蛋白表达量呈现增高现象(P<0.01),见图3、4。通过qRT-PCR再次观察了KLF4 mRNA表达量变化,结果同样显示巨噬细胞内KLF4 mRNA表达量较LG组下降(P<0.01),在WG干预后,KLF4 mRNA的表达量呈现剂量依耐性增高现象(P<0.01),见图5A。
3 讨论
DN已经成为终末期肾脏病的主要原因之一,且在中国人群中的患病率逐年上升[8]。众所周知,自身免疫病发展与巨噬细胞的长期激活有关,持续不断分泌的炎症因子,最终对正常组织及细胞造成无法修复的损害,导致肾组织纤维化及肾脏功能下降[9]。本实验以高糖刺激的巨噬细胞为研究对象,模拟糖尿病高糖环境,探究WG抑制DN炎症发展的可能机制。
在长期高血糖状态下,巨噬细胞在肾组织中异常聚集,激活TLR4、NF-κB等通路,分泌大量TNF-α、IL-1β炎症因子,导致肾小球、肾小管及基底膜病理性改变,进而出现蛋白尿增多等临床症状。TNF-α和IL-1β被称作巨噬细胞炎症的“警报细胞因子”,它们会进一步诱导促炎基因iNOS的产生。此次研究中采用高糖刺激小鼠RAW264.7巨噬细胞,观察炎症信号蛋白及相关炎症因子表达。结果表明,高糖刺激后,NF-κB p65、NF-κB pp65、iNOS表达增高,促炎因子TNF-α、IL-1β表达明显上调,巨噬细胞极化。与Zhu et al[9]及汤祥瑞 等[10]研究结果相似,高糖可以直接或间接的刺激巨噬细胞迁移、炎症因子的分泌及iNOS表达增加。
巨噬细胞是先天免疫的重要参与者,而KLF4是巨噬细胞生物学中的关键介质,起着调节巨噬细胞促炎效应。本研究显示,KLF4在高糖诱导的RAW264.7细胞中表达下调,并且伴随炎症因子的释放及iNOS的增加。与多数研究[11-12]结果一致,KLF4在肾脏保护及炎症抑制方面有极大地探索意义。有研究[2-3, 13]报道KLF4抑制STAT3、mTOR和NF-κB p65的激活,以改善动脉粥样硬化及脓毒血症等疾病的进展。此研究验证出高糖刺激巨噬细胞24 h出现抑制KLF4表达的情况,说明在炎症状态中KLF4是处于低表达状态。也有文献[3]报道在炎症激活过程中,NF-κB p65活性增强抑制KLF4的转录作用,KLF4表达下调会激活NF-κB pp65的表达,两者形成正反馈效应加快炎症反应的进行。由此推测KLF4介导是DN炎症发展过程的重要环节。
WG是一种低毒、低成本、高疗效的黄酮类天然化合物[8],具有抗炎、抗氧化、抗癌等作用。多数研究证明,WG有显著的抗炎活性,可以明显减轻结肠炎[6]、脂肪肝[14]及慢性咽炎[15]等疾病进展,但在DN介导的慢性炎症中的作用及机制还有待探索。在本研究中,通过CCK-8实验验证高糖条件下细胞安全浓度范围,再用不同浓度的WG给予细胞预处理,然后再用高糖刺激巨噬细胞活化。结果表明,WG减轻HG组巨噬细胞TNF-α、IL-1β的分泌和iNOS的表达,这种作用是通过抑制NF-κB p65的磷酸化,逆转巨噬细胞抗炎活性,成功减轻了高糖诱导的巨噬细胞炎症。此外,KLF4表达量在WG作用下呈剂量依赖性递增,提示KLF4不仅是炎症中的重要参与者,更在DN中发挥保护性作用,并参与到WG的治疗机制中。
综上所述,本研究表明高糖刺激巨噬细胞后,其KLF4表达减少,而WG可以刺激巨噬细胞KLF4的表达增高,进而抑制高糖刺激巨噬细胞产生促炎因子和炎性物质,其机制是由于KLF4表达增高抑制了NF-κB p65的磷酸化,进而减少了iNOS的表达。提示KLF4/NF-κB相关信号通路可能参与到WG影响巨噬细胞改善DN的过程。由于本研究缺乏对相关靶向蛋白干预和相应体内模型验证,将在以后的研究中加以探讨,以期为DN的治疗提供新的思路。
