海南岛滨海沙生植物叶片功能性状分异及其与土壤因子的关系
2022-10-17王旭刘敬坤罗水星赵坤坤李新建
王旭, 刘敬坤, 罗水星, 赵坤坤, 李新建
海南岛滨海沙生植物叶片功能性状分异及其与土壤因子的关系
王旭1, 刘敬坤1, 罗水星2, 赵坤坤2, 李新建3*
(1. 中国林业科学研究院热带林业研究所,广州 510520;2. 中国林业科学研究院热带林业研究所试验站,海南 乐东 572542;3. 国家林业和草原局中南调查规划设计院,长沙 410014)
为探讨滨海沙生植物对环境的适应策略,对海南岛滨海沙生植物单叶蔓荆、苦郎树、马缨丹、飞机草、假马鞭、厚藤的叶片主要功能性状分异特征及其土壤因子的关系进行了研究。结果表明,热带滨海沙生植物叶片功能性状存在明显的种间分异,叶片功能性状不仅受植物种类的影响,还受土壤因子影响;比叶面积呈现草本>灌木>藤本的规律;叶片的N/P为7.78~ 10.85,热带滨海沙生植物生长受土壤氮限制;叶片功能性状中Na含量的变异系数最大(18.46%~76.36%),说明不同植物对Na+的吸收存在较大差异,这将影响其在滨海沙地的自然分布;比叶面积、叶干物质含量与叶片的K、Na含量呈负相关,叶N/P与土壤Na+含量呈负相关,叶片Na+含量与土壤有机质、全磷、全钾呈负相关,土壤盐分限制植物对氮磷的吸收,滨海沙生植物通过比叶面积变化来适应盐胁迫。因此,在滨海沙生植被恢复中,施加土壤肥力是其快速恢复的重要措施,叶片Na+含量可作为滨海沙生植物耐盐性筛选的主要指标。
沙生植物;功能性状;土壤化学性质;适应性
植物功能性状是植物与环境长期相互作用,以最大程度减小环境的不利影响,逐渐形成了许多生理和形态方面的适应对策而表现出来的性状[1]。叶功能性状与植物的生长对策有着紧密的联系[2]。但不同植物的叶功能性状变异特征不同,陈文等[3]对粤东89种常见植物叶功能性状研究表明,科级水平的比叶面积(SLA)、干物质含量(LMDC)差异显著,藤本与灌木叶面积显著大于草本植物,即使同植物种不同品种间也存在较大的差异。此外环境的变化也会导致叶片功能性状的分异,在逆境条件下,植物为适应环境胁迫,如干旱、盐碱等胁迫, 常通过叶功能性状的调整,来提高对生境的适应[4]。已有研究表明,肥沃土壤上生长的植物有较大的比叶面积和较高的相对生长速率[5];从湿润到干旱的环境梯度上,植物叶片会由大变小,N和P含量增加、C/N下降、同化速率升高[6–7]。李芳兰等[8]报道,干旱胁迫限制了白刺花(i)新叶的发生与单叶面积扩展。黎洁[9]对广西北仑河口海岸带红树林的研究表明,土壤P含量与叶片的C、C/P、N/P存在显著相关性。温度对叶片养分也会产生一定的影响, Reich等[10]的研究表明,随年均温的升高,叶片N、P含量呈下降趋势。空气质量对植物生长也会产生影响,盐雾胁迫会导致植物体内丙二醛大量积累[11],对植物生长发育和某些器官产生影响, 如根、茎、整株生物量增加,株高、花序数和视觉质量下降,且草本植物更易受盐雾的侵害[12]。
了解植物群落与环境间的功能关系一直是生态学领域的一大挑战[13–14]。叶功能性状与土壤养分供应密切相关[15–16]。滨海沙地是海洋、陆地和大气的交界面,生境严峻,土壤贫瘠,保肥力差,易透水,易干旱,阳光强烈,常年风大,盐分高,植被演替困难[17]。滨海沙生植物,由于海水潮汐的影响,形成了明显的分布特征[17–18],处于不同潮位线的植物由于土壤水分和质地的异质性,如叶片革质或肉质、表面多绒毛[19],叶子或短枝退化成枝刺,叶片具盐腺或盐泡[20],形成发达的根系[21]等,是“狭阈生态特征明显”的类群,成为滨海沙地特有种,表现出不同的叶片功能性状。虽然对植物叶片功能性状研究较多,但对高温、干旱、高盐的热带滨海潮汐带沙地研究缺乏。有研究表明,SLA与植物相对生长速率、光合速率等有关[6,22],是植物生理过程的最佳指示者[23];LDMC是反映植物生态行为差异的重要指标之一,常与潜在相对生长速率有关;叶片pH主要由植物种类决定,具有重要的指标生态系统地球化学循环过程和特征的作用[24];叶N/P反映了土壤N、P供给能力[25];叶片K、Na含量可评估植物的耐盐能力[26]。因此本研究在海南岛北部半湿润区、南部半干半湿润区、西部半干旱区和东部湿润区4个不同气候区采集滨海沙生植物叶片,测定叶片的SLA、LDMC、pH、N/P、Na和K含量,探讨滨海沙地常见植物的叶片功能分异特征,及土壤因子对其分异特征的影响,为研究滨海沙生植物对环境的适应策略提供科学依据。
1 研究区概况
海南岛处于热带北缘,位于北纬18°08′~20°10′,东经108°37′~111°03′,海岸线长1 890 m,岛面积3.39×104km2,以砂质岸线为主,占海南自然岸线的42.5%[19]。海南岛属热带季风气候,全年无冬, 年均温为22 ℃~27 ℃,最低温在10 ℃以上,年均日照时数1 750~2 650 h,年均降水量为1 639 mm,其中5—10月的雨量超过全年总雨量的70%以上,约为1 500 mm,全岛水热资源充足。考虑到水热条件对植物功能性状的影响,尤其是降水,本研究在北部半湿润区临高、南部半干半湿润区三亚和西部半干旱区昌江和东部湿润区文昌进行采样(表1, 2)。
2 材料和方法
2.1 材料
根据实地踏查,海南滨海潮汐带沙地植物主要由灌木、多年生草本和藤本3类植物组成,少有乔木树种,因此选择常见的单叶蔓荆()、苦郎树()、马缨丹()、飞机草()、假马鞭()、厚藤()为研究材料(表3)。
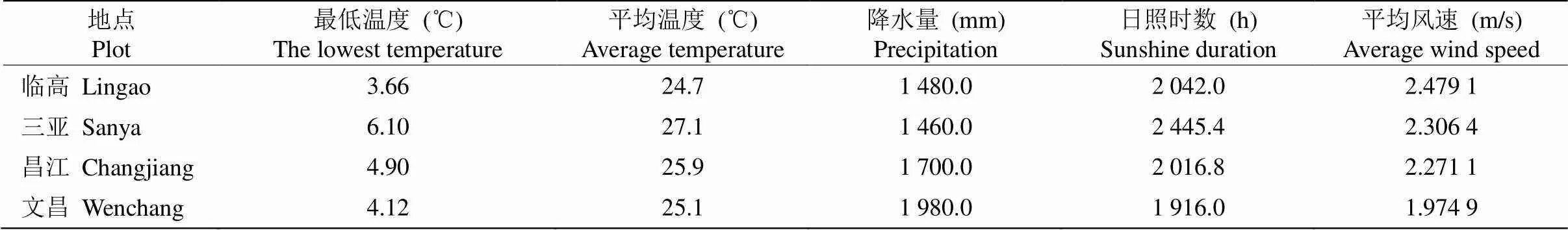
表1 采样地基本气象资料[27]
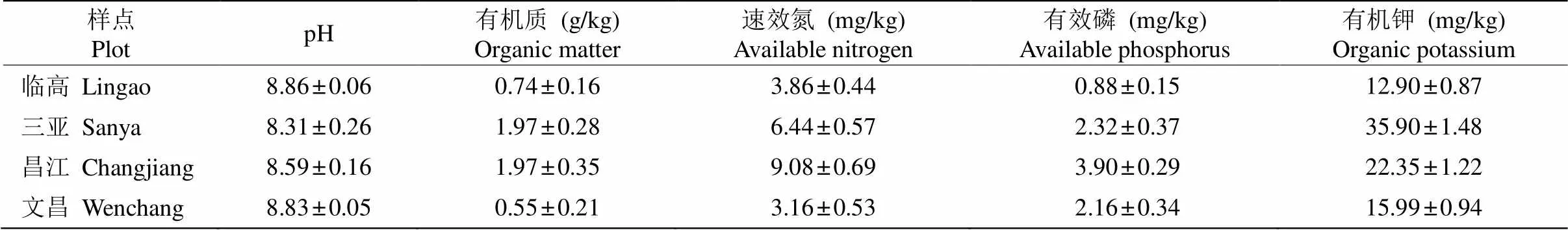
表2 研究点土壤主要化学性质

表3 研究材料
LG: 临高; WC: 文昌; CJ: 昌江; SY: 三亚。以下图表同。
LG: Lingao; WC: Wenchang; CJ: Changjiang; SY: Sanya. The same is following Tables and Figures.
2.2 样品采集和处理
叶样品采集:灌木,在20 m×20 m样方内每种随机选择生长良好、个体大小基本一致的5株植株,在每株冠层的东、南、西、北4个方向,采集全展开、无病虫害的成熟叶片,每种不少于20片。藤本和草本植物,在20 m×20 m样方内随机采集全展开、无病虫害的成熟叶片,每种不少于20片。用电子天平及时称量叶片鲜质量,然后用CID CI-203手持式激光叶面积仪(美国)测叶面积;再将叶片放入纸袋内带回实验室,105 ℃杀青20 min后,在80 ℃烘箱内烘48 h至恒定,称量干质量。土壤样品采用机械取样法,按照四分法取样,将样方内0~30 cm土层的土壤样品混合,每个样点3个重复,测定土壤理化性质。另用环刀取样测定土壤水分和容重, 3个重复。分析临高(LG)采样点不同植物间叶片功能性状的差异,同时分析厚藤和单叶蔓荆叶片功能性状在不同采样点间的差异。用变异系数()分析不同植物和不同生境对叶片功能性状的分异情况,参照Cornelissen等[28]的方法计算叶片功能性状。= SD/MN×100%, SLA=/, LDMC=/m, 式中, SLA为比叶面积(cm2/g),为叶面积(cm2),LDMC为叶片干物质量,为叶干质量(g),m为鲜叶质量(g),为变异系数,SD为标准偏差,MN为平均值。
采用R 3.5.1进行做图和统计分析,差异性分析采用最小显著性差异法,采用R语言cor函数Pearson相关系数对叶片功能性状及其与土壤化学性质进行相关分析。
2.3 土壤理化性质和叶片化学成分测定
土壤理化性质测定参照国家或地方标准(LY/T 1268—1999、NY/T 2017—2011、LY/T 1237—1999、DB12/T 846—2018)进行分析。土壤和叶片pH采用电位法(土∶水=1∶2.5);土壤有机质采用重铬酸钾氧化-外加热法,土壤全氮和叶片氮含量采用凯氏消煮扩散法,土壤全磷和叶片磷含量采用氢氧化钠碱熔-钼锑抗比色法,土壤全钾和叶片钾含量采用氢氧化钠碱熔-火焰光度法,Na+测定采用火焰光度法, 土壤水解氮采用碱解扩散法、土壤有效磷采用HCl- H2SO4浸提法、土壤有效钾采用乙酸铵浸提-火焰光度法、土壤电导率采用电导法测定,土壤水分采用烘干法。
3 结果和分析
3.1 不同植物叶片功能性状的分异特征
比叶面积和叶干物质含量综合反映了植物利用资源的能力。从图1可见,6种滨海沙生植物的比叶面积、叶片干物质含量均存在显著差异,比叶面积为83.26~205.62 cm2/g,其中单叶蔓荆最小, 飞机草最大,相差达2.5倍。干物质含量为0.297~ 0.134,其中单叶蔓荆最大,厚藤最小。叶片的化学计量特征不仅能反映植物对资源的利用情况,还可反映植物的养分适应策略。6种沙生植物叶片pH均表现为酸性,为5.43~6.58,其中单叶蔓荆最小, 马缨丹最大,单叶蔓荆与飞机草,厚藤、假马鞭和苦郎树间的pH无显著差异;叶片N/P为7.78~9.51, 其中飞机草最大,苦郎树最小,但不同植物间无显著差异;叶片的全K和Na+含量均为厚藤最高,苦郎树次之,飞机草最低,且厚藤与苦郎树间无显著差异,说明厚藤和苦郎树对Na、K有较强的富集作用。6种沙生植物的比叶面积、叶干物质含量、pH、N/P、全K和Na+的变异系数分别为32.10%、27.47%、7.17%、7.04%、33.28%和76.36%,以Na+含量的变异系数最大,说明热带沙生植物对Na+的吸收能力差异较大。

图1 植物叶片功能性状的差异。1: 单叶蔓荆; 2: 飞机草; 3: 厚藤; 4: 假马鞭; 5: 苦郞树; 6: 马缨丹;SLA: 比叶面积; LDMC: 叶干物质量;TK: 全钾;柱上不同字母表示差异显著(P<0.05, TukeyHSD检验)。下图同。
3.2 不同地点植物叶功能性状分异特征
为比较不同气候区域共有种的叶片功能性状间的差异,选择大部分样点共有的厚藤进行研究。从图2可见,叶片的形态性状表现不一致,临高与三亚,昌江与文昌的比叶面积无显著差异,而除三亚与文昌的叶干物质含量无显著差异外,其他样点间均存在显著差异。而叶片化学性状也同样表现不一致,pH的差异性与叶干物质含量表现一致,而N/P除临高与三亚间无显著差异,其他样点间均有显著差异,叶全K含量除昌江与三亚间无显著差异,其他样点间均存在显著差异,三亚与昌江、临高和文昌的Na+含量差异显著,其余样点间无显著差异,说明外部环境因素可能对叶片的功能性状产生影响,如降水、土壤理化性质等。厚藤的比叶面积、叶干物质含量、pH、N/P、全K和Na+含量在4个取样点间的变异系数分别为9.58%%、9.03%、1.96%、14.26%、15.72%和18.46%,以Na+含量变异系数最大。

图2 不同样点厚藤的叶片功能性状
3.3 叶片功能性状间相关性
从图3可见,临高6种沙生植物叶片功能性状中,pH与K含量的相关性最大,K含量与Na+含量相关性最大,叶干物质量、比叶面积与K、Na+含量均呈负相关,比叶面积与叶氮磷比呈正相关。不同采样点单叶蔓荆的叶干物质量与N/P呈正相关,与K、Na+含量和比叶面积呈负相关, N/P与K含量、pH与比叶面积和Na+含量、K含量与Na+含量均呈负相关;厚藤的比叶面积与pH呈正相关,与Na+含量、干物质量、N/P呈负相关,pH与K含量呈正相关,与Na+含量、干物质量、N/P呈负相关,K含量与N/P、叶干物质量呈负相关, Na+含量与干物质量呈负相关,叶干物质量与N/P呈正相关。这也说明植物对K、Na+的吸收限制了植物的生长,对N、P的利用率影响植物的生长。
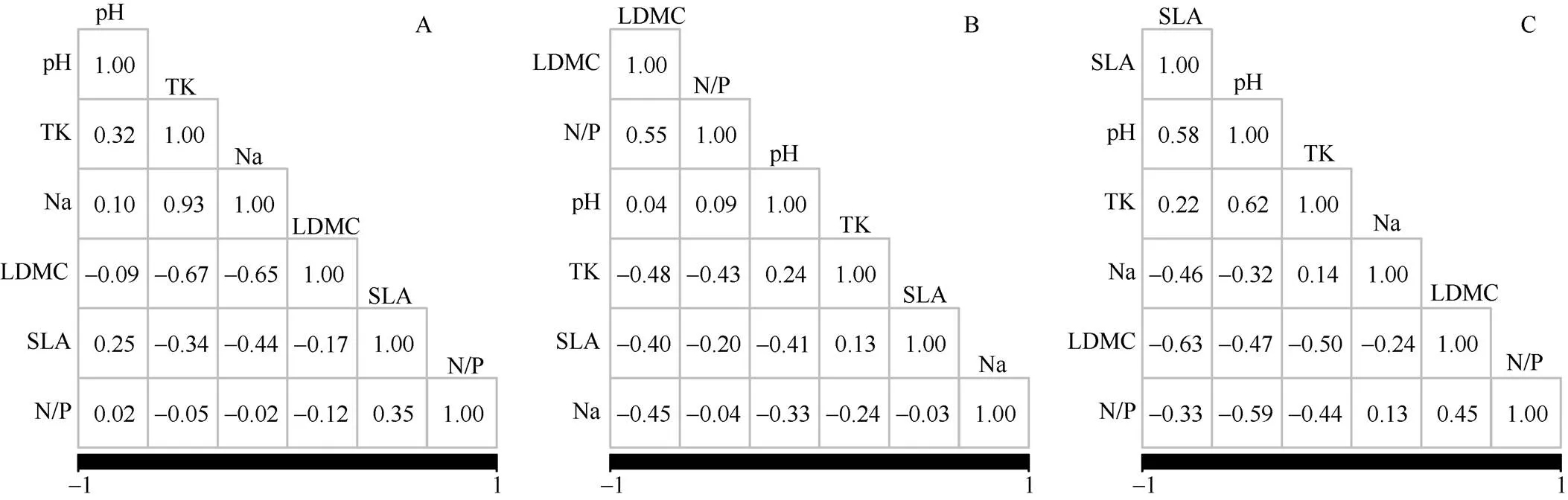
图3 叶功能性状间的相关性。A: 临高采样点6种植物; B: 不同采样点的单叶蔓荆; C: 不同采样点的厚藤。
3.4 叶片功能性状与土壤化学性质的相关性
土壤是植物生长的物质基础。从表4可见,比叶面积与土壤的有机质、总P、总N和有效K呈正相关,与pH呈负相关;叶干物质含量与土壤水分和总P呈负相关,叶pH与土壤pH呈负相关,与土壤有机质、总P和有效K呈正相关;叶N/P与土壤Na含量呈负相关,与土壤水分呈正相关;叶K含量与土壤Na含量、有效K、总P呈正相关, 与土壤pH呈负相关;叶Na含量与土壤pH呈正相关,而与土壤有机质、总P、总K呈负相关。土壤电导率与叶功能性状间的相关性较弱。可见,比叶面积受土壤有机质、总P、总N、有效K和pH影响较大,叶干物质含量受土壤水分和总P影响较大,叶N/P受土壤Na+和土壤水分影响较大,叶K含量受土壤Na含量、有效K、总P和pH影响较大;叶Na含量受土壤pH、有机质、总P、总K影响较大,叶功能性状中比叶面积、氮磷比、钾含量、pH等受土壤Na、K含量和pH影响较大。

表4 叶片功能性状与土壤化学性质相关性
SOC: 土壤有机质; TN: 总氮; TP: 总磷; HN: 水解氮; EP: 有效磷; EK: 有效钾; SEC: 电导率; SH: 含水量。
SOC: Soil organic matter; TN: Total N; TP: Total P; HN: Hydrolyzed N; EP: Available P; EK: Available K; EC: Electrical conductivity; SH: Moisture content.
4 结论和讨论
叶功能性状是植物为适应环境而表现出来的叶片水平功能特征。比叶面积(SLA)表征了植物对光能的捕获能力,较高的比叶面积被认为是植物为了快速生长所采取的一种生存策略[23,29]。土壤贫瘠导致植物SLA偏小,生长缓慢[30]。本研究中,6种沙生植物的叶功能性状存在较大差异,飞机草的比叶面积最大,单叶蔓荆最小,且呈现草本植物>木本植物>藤本植物的规律。这与前人[15,31]对沙丘植物的研究结果较一致。比叶面积与光合速率等有关[22],是植物对光能捕捉能力的重要表征指标。本研究中植物主要分布在潮上带,飞机草为草本,以须根系为主,沙地保水保肥能力差,为了在适宜条件下快生长,通过扩大叶面积来捕获更多光能的权衡策略弥补根系养分吸收能力的不足,为完成生活史贮存能量,而木本植物具有深的根系,藤本植物通过长的匍匐茎增加对养分吸收面积,从而提高资源的获取能力[13]。一般来说叶干物质含量高,植物贮存的营养物质量多。水分是影响滨海沙地演替的主要因素之一[32]。6种植物中厚藤的叶干物质含量最小,这可能与厚藤为匍匐生长藤本植物,茎的节点处多有根系,较易获得水分和养分,从而缓解沙地水分和养分的限制作用。植物的N/P常用来表征植物养分亏缺状况,N/P<14表明植物生长受N限制;N/P>16表明植物生长受P限制[25,33–34]。6种沙生植物N/P为7.78~9.51,且种间差异不显著, 不同采样点的叶N/P<14,说明热带滨海沙生植物生长受氮限制。叶全K和Na+含量均表现为厚藤、苦郎树最高, 这可能与其生境有关,单叶蔓荆、厚藤、苦郎树主要分布在潮间带,而飞机草、假马鞭、马缨丹主要分布在潮上带。潮间带因海水反复冲洗, 表现高盐分低养分的特征,植物在受到干旱胁迫或者盐分胁迫时,由低亲和K+吸收系统转换为高亲和K+吸收系统,以促进K+的吸收[35]。
本研究中,不同样点厚藤的叶片功能性状并未表现出一致的规律性,昌江的比叶面积最小,临高的最大,而单叶蔓荆以文昌最大,昌江最小;临高与文昌的厚藤叶干物质含量差异显著,而单叶蔓荆则无显著差异。叶片N/P和全K含量等指标也表现出同样的规律。这可能是植物功能性状并不是单独响应环境变化,而是在长期适应环境的过程中,通过调节资源的分配实现彼此之间的协同与权衡,从而提高植物的适应性[36]。本研究中比叶面积与土壤有机质、有效K呈显著的正相关,4个样点中昌江的土壤有效K含量最低,土壤有机质仅次于文昌, 处于次低位,从而导致文昌植物的比叶面积受土壤有效K含量和有机质限制;再者滨海植物为外生演替植物,由于风大、干旱、盐生等影响,每种植物会有1个突出的功能性状以适应逆境[32];厚藤与单叶蔓型虽然都具有匍匐的茎,但其生活型不同,前者为草本,全株光滑,后者为灌木,且小枝和叶密生细柔毛[37],对滨海沙地产生不同的适应方式。比叶面积与叶干物质含量间呈负相关关系[38],本研究结果与其一致,只是相同地点不同植物中呈弱的负相关,不同地点同种植物呈较强的负相关,这也进一步说明植物种类对这一规律的表现更明显。
在本研究中,比叶面积、叶干物质含量与叶片K、Na含量呈负相关,叶N/P与土壤Na+含量呈负相关,叶片Na+含量与土壤有机质、总P、总K呈负相关。由于Na+是表征盐胁迫的主要因子,植物吸收较多的Na+,会改变细胞膜的结构和功能,细胞膜上原有Ca2+被Na+取代,导致细胞内离子种类和浓度发生变化,核酸和蛋白质的合成和分解的平衡受到破坏,从而严重影响植物的生长发育[39–40]。K+是植物生长必不可少的常量元素,主要通过植物根系吸收,对缓解植物胁迫起着重要作用,盐胁迫能够降低根部吸收K+的能力[41]。滨海沙生植物在受到盐胁迫时,通过调节比叶面积来缓解胁迫, 同时盐胁迫直接影响植物对干物质的积累,以及对N、P的吸收,进而影响植物的生长发育。此外,从本研究结果来看,可以通过增加土壤肥力减少植物对Na+的吸收[42],来缓解盐分对植物生长发育的胁迫。
[1] MENG T T, NI J, WANG G H. Plant functional traits, environments and ecosystem functioning [J]. Chin J Plant Ecol, 2007, 31(1): 150–165. doi: 10.17521/cjpe.2007.0019.
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能 [J]. 植物生态学报, 2007, 31(1): 150–165. doi: 10.17521/cjpe.2007.0019.
[2] VENDRAMINI F, DÍAZ S, GURVICH D E, et al. Leaf traits as indicators of resource-use strategy in floras with succulent species [J]. New Phytol, 2002, 154(1): 147–157. doi: 10.1046/j.1469-8137.2002. 00357.x.
[3] CHEN W, WANG J H, MA R J, et al. Variance in leaf functional traits of 89 species from the Guangdong of China [J]. Chin J Ecol, 2016, 35 (8): 2101–2109. doi: 10.13292/j.1000-4890.201608.033.
陈文, 王桔红, 马瑞君, 等. 粤东89种常见植物叶功能性状变异特征 [J]. 生态学杂志, 2016, 35(8): 2101–2109. doi: 10.13292/j.1000- 4890.201608.033.
[4] ZHAO X W, WANG Y F, MA W M. The relationship of transpiration rate with leaf characters ofin different slopes of an alpine meadow [J]. Acta Ecol Sin, 2019, 39(7): 2494–2500. doi: 10. 5846/stxb201801170131.
赵夏纬, 王一峰, 马文梅. 高寒草地不同坡向披针叶黄华蒸腾速率与叶性状的关系 [J]. 生态学报, 2019, 39(7): 2494–2500. doi: 10. 5846/stxb201801170131.
[5] RODERICK M L, BERRY S L, NOBLE I R. A framework for understanding the relationship between environment and vegetation based on the surface area to volume ratio of leaves [J]. Funct Ecol, 2000, 14(4): 423–437. doi: 10.1046/j.1365-2435.2000.00438.x.
[6] REICH P B, ELLSWORTH D S, WALTERS M B, et al. Generality of leaf trait relationships: A test across six biomes [J]. Ecology, 1999, 80(6): 1955–1969. doi: 10.1890/0012-9658(1999)080[1955:GOLTRA] 2.0.CO;2.
[7] WRIGHT I J, REICH P B, WESTOBY M, et al. The worldwide leaf economics spectrum [J]. Nature, 2004, 428(6985): 821–827. doi: 10. 1038/nature02403.
[8] LI F L, BAO W K, WU N. Morphological and physiological responses of currentseedlings to drought stress [J]. Acta Ecol Sin, 2009, 29(10): 5406–5416. doi: 10.3321/j.issn:1000-0933.2009.10.027.
李芳兰, 包维楷, 吴宁. 白刺花幼苗对不同强度干旱胁迫的形态与生理响应 [J]. 生态学报, 2009, 29(10): 5406–5416. doi: 10.3321/j. issn:1000-0933.2009.10.027.
[9] LI J. Plant population dynamics and ecological stoichiometry of man- grove in Beilun estuary in Guangxi [D]. Nanning: Guangxi Teachers Education University, 2017.
黎洁. 广西北仑河口红树林植物种群动态与生态化学计量特征研究 [D]. 南宁: 广西师范学院, 2017.
[10] REICH P B, UHL C, WALTERS M B, et al. Leaf demography and phenology in Amazonian rain forest: A census of 40 000 leaves of 23 tree species [J]. Ecol Monogr, 2004, 74(1): 3–23. dio: 10.1890/02- 4047.
[11] CHEN G J, LIU W G, XU Y C, et al. Evaluation of salt spray tolerance of 9 coastal plants [J]. J For Environ, 2018, 38(3): 341–347. doi: 10. 13324/j.cnki.jfcf.2018.03.013.
陈国军, 刘维刚, 徐迎春, 等. 9种滨海植物盐雾的耐性评价 [J]. 森林与环境学报, 2018, 38(3): 341–347. doi: 10.13324/j.cnki.jfcf.2018. 03.013.
[12] SCHEIBER S M, SANDROCK D, ALVAREZ E, et al. Effect of salt spray concentration on growth and appearance of ‘Gracillimus’ maiden grass and ‘Hamelin’ fountain grass [J]. Hort Technol, 2008, 18(1): 34– 38. doi: 10.21273/HORTTECH.18.1.34.
[13] ZHANG S H, ZHANG Y, XIONG K N, et al. Changes of leaf functional traits in karst rocky desertification ecological environment and the driving factors [J]. Glob Ecol Conserv, 2020, 24: e01381. doi: 10.1016/j.gecco.2020.e01381.
[14] HE P C, YE Q. Plant functional traits: From individual plant to global scale [J]. J Trop Subtrop Bot, 2019, 27(5): 523–533. doi: 10.11926/jtsb. 4108.
贺鹏程, 叶清. 基于植物功能性状的生态学研究进展: 从个体水平到全球尺度 [J]. 热带亚热带植物学报, 2019, 27(5): 523–533. doi: 10.11926/jtsb.4108.
[15] LI S J, WANG H, GOU W, et al. Leaf functional traits of dominant desert plants in the Hexi Corridor, northwestern China: Trade-off relationships and adversity strategies [J]. Glob Ecol Conserv, 2021, 28: e01666. doi: 10.1016/j.gecco.2021.e01666.
[16] SHIPLEY B, DE BELLO F, CORNELISSEN J H C, et al. Reinforcing loose foundation stones in trait-based plant ecology [J]. Oecologia, 2016, 180(4): 923–931. doi: 10.1007/s00442-016-3549-x.
[17] WANG W Q, CHEN Y F, LI Q Q, et al. Coastal Sand Plant Resources and Vegetation Restoration of South China [M]. Xiamen: Xiamen University Press, 2016.
王文卿, 陈洋芳, 李芊芊, 等. 南方滨海沙生植物资源及沙地植被修复 [M]. 厦门: 厦门大学出版社, 2016.
[18] HUANG Y Q, WANG J J, ZHANG R T, et al. The resources and protection of psammophilous vegetation on coasts in Xiamen Guan- yinshan, Fujian Province [J]. Subtrop Plant Sci, 2013, 42(1): 73–76. doi: 10.3969/j.issn.1009-7791.2013.01.016.
黄雅琴, 王建军, 张娆挺, 等. 厦门观音山海岸沙生植被资源现状及保护对策 [J]. 亚热带植物科学, 2013, 42(1): 73–76. doi: 10.3969/ j.issn.1009-7791.2013.01.016.
[19] LI L, JIA Z Q, ZHU Y J, et al. Research advances on drought resistance mechanism of plant species in arid area of China [J]. J Dese Res, 2010, 30(5): 1053–1059.
李磊, 贾志清, 朱雅娟, 等. 我国干旱区植物抗旱机理研究进展 [J]. 中国沙漠, 2010, 30(5): 1053–1059.
[20] ZHOU L L, LIU P, WANG J. Nutritive organs anatomical structure of two species ofin Xinjiang [J]. Acta Bot Boreali-Occid Sin, 2007, 27(6): 1127–1133. doi: 10.3321/j.issn:1000-4025.2007.06.010.
周玲玲, 刘萍, 王军. 新疆2种盐生补血草营养器官的解剖学研究 [J]. 西北植物学报, 2007, 27(6): 1127–1133. doi: 10.3321/j.issn:1000- 4025.2007.06.010.
[21] DU J H, LIU A L, DONG Y X, et al. Architectural characteristics of roots in typical coastal psammophytes of south China [J]. Chin J Plant Ecol, 2014, 38(8): 888–895. doi: 10.3724/SP.J.1258.2014.00083.
杜建会, 刘安隆, 董玉祥, 等. 华南海岸典型沙生植物根系构型特征 [J]. 植物生态学报, 2014, 38(8): 888–895. doi: 10.3724/SP.J.1258. 2014.00083.
[22] POORTER H, DE JONG R. A comparison of specific leaf area, chemical composition and leaf construction costs of field plants from 15 habitats differing in productivity [J]. New Phytol, 1999, 143(1): 163–176. doi: 10.1046/j.1469-8137.1999.00428.x.
[23] WILSON P J, THOMPSON K, HODGSON J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies [J]. New Phytol, 1999, 143(1): 155–162. doi: 10.1046/j.1469-8137.1999. 00427.x.
[24] CORNELISSEN J H C, SIBMA F, VAN LOGTESTIJN R S P, et al. Leaf pH as a plant trait: Species-driven rather than soil-driven variation [J]. Fun Ecol, 2011, 25(3): 449–455. doi: 10.1111/j.1365-2435.2010. 01765.x.
[25] KOERSELMAN W, MEULEMAN A F M. The vegetation N∶P ratio: A new tool to detect the nature of nutrient limitation [J]. J Appl Ecol, 1996, 33(6): 1441–1450. doi: 10.2307/2404783.
[26] AN D, CHEN J G, GAO Y Q, et al.drives adaptation ofto salinity by reducing floral sodium content [J]. PLoS Genet, 2017, 13(10): e1007086. doi: 10.1371/journal.pgen.100 7086.
[27] LI W T, ZHANG M J, ZHANG J B, et al. GIS-based small-grid simulation of climate resources in Hainan Island [J]. Chin J Trop Agric, 2020, 40(9): 83–93. doi: 10.12008/j.issn.1009-2196.2020.09.015.
李文韬, 张明洁, 张京红, 等. 基于GIS的海南岛气候资源小网格推算研究 [J]. 热带农业科学, 2020, 40(9): 83–93. doi: 10.12008/j. issn.1009-2196.2020.09.015.
[28] CORNELISSEN J H C, LAVOREL S, GARNIER E, et al. A handbook of protocols for standardized and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 2003, 51(4): 335–380. doi: 10.1071/ BT02124.
[29] POORTER H, NIINEMETS Ü, POORTER L, et al. Causes and conse- quences of variation in leaf mass per area (LMA): A meta-analysis [J]. New Phytol, 2009, 182(3): 565–588. doi: 10.1111/j.1469-8137.2009. 02830.x.
[30] THUILLER W, LAVOREL S, MIDGLEY G, et al. Relating plant traits and species distributions along bioclimatic gradients for 88taxa [J]. Ecology, 2004, 85(6): 1688–1699. doi: 10.1890/03-0148.
[31] LI Y L, CUI J Y, SU Y Z. Specific leaf area and leaf dry matter content of some plants in different dune habitats [J]. Acta Ecol Sin, 2005, 25(2): 304–311. doi: 10.3321/j.issn:1000-0933.2005.02.019.
李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较 [J]. 生态学报, 2005, 25(2): 304–311. doi: 10.3321/j. issn:1000-0933.2005.02.019.
[32] REN H, JIAN S G, ZHANG Q M, et al. Plants and vegetation on South China Sea Islands [J]. Ecol Environ Sci, 2017, 26(10): 1639–1648. doi: 10.16258/j.cnki.1674-5906.2017.10.001.
任海, 简曙光, 张倩媚, 等. 中国南海诸岛的植物和植被现状 [J]. 生态环境学报, 2017, 26(10): 1639–1648. doi: 10.16258/j.cnki.1674- 5906.2017.10.001.
[33] CERNUSAK L A, WINTER K, TURNER B L. Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physio- logical and environmental controls [J]. New Phytol, 2010, 185(3): 770– 779. doi: 10.1111/j.1469-8137.2009.03106.x.
[34] JIA J, BAI J H, WANG W, et al. Changes of biogenic elements inandfrom salt marshes in Yellow River delta, China [J]. Chin Geogr Sci, 2018, 28(3): 411–419. doi: 10. 1007/s11769-018-0959-1.
[35] LU L M, YANG T Z. Potassium transporters and their molecular regulation in higher plants [J]. Acta Bot Boreali-Occid Sin, 2006, 26 (11): 2402–2410. doi: 10.3321/j.issn:1000-4025.2006.11.034.
鲁黎明, 杨铁钊. 高等植物K+吸收转运蛋白及其分子调节 [J]. 西北植物学报, 2006, 26(11): 2402–2410. doi: 10.3321/j.issn:1000-4025. 2006.11.034.
[36] CORNWELL W K, SCHWILK D W, ACKERLY D D. A trait-based test for habitat filtering: Convex hull volume [J]. Ecology, 2006, 87(6): 1465–1471. doi: 10.1890/0012-9658(2006)87[1465:ATTFHF]2.0.CO;2.
[37] Guangdong Institute of Botany. Flora of Hainan, Vol. 4 [M]. Beijing: Science Press, 1977.
广东省植物研究所. 海南植物志, 第4卷 [M]. 北京: 科学出版社, 1977.
[38] WU M L, ZHU J, ZHU Q, et al. Analysis of leaf functional traits and functional diversity of woody plants in evergreen and deciduous broad-leaved mixed forest of Xingdoushan [J]. Acta Bot Boreali-Occid Sin, 2019, 39(9): 1678–1691. doi: 10.7606/j.issn.1000-4025.2019.09. 1678.
吴漫玲, 朱江, 朱强, 等. 星斗山常绿落叶阔叶混交林木本植物叶功能性状及其多样性特征分析 [J]. 西北植物学报, 2019, 39(9): 1678–1691. doi: 10.7606/j.issn.1000-4025.2019.09.1678.
[39] WANG L Q, SHAO M A. The salt influx, transport and accumulation in higher plant [J]. Agric Res Arid Areas, 2005, 23(5): 244–249. doi: 10.3321/j.issn:1000-7601.2005.05.048.
王林权, 邵明安. 高等植物对Na+的吸收、运输和累积 [J]. 干旱地区农业研究, 2005, 23(5): 244–249. doi: 10.3321/j.issn:1000-7601. 2005.05.048.
[40] CHEN P C, CHEN X F, MA B J, et al. Na+homeostasis and salt tolerance of plants [J]. J Zhejiang Norm Univ (Nat Sci), 2016, 39(2): 207–214. doi: 10.16218/j.issn.1001-5051.2016.02.014.
陈鹏程, 陈析丰, 马伯军, 等. 植物耐盐性与Na+动态平衡研究进展 [J]. 浙江师范大学学报(自然科学版), 2016, 39(2): 207–214. doi: 10. 16218/j.issn.1001-5051.2016.02.014.
[41] REN Z B, WANG Z G, NIE Q J, et al. Effect of salt stress on growth and ion contents incuttings [J]. J NE For Univ, 2011, 39(5): 24–26. doi: 10.3969/j.issn.1000-5382.2011.05.008.
任志彬, 王志刚, 聂庆娟, 等. 盐胁迫对锦带花幼苗生长及不同部位Na+、K+、Ca2+、Mg2+离子质量分数的影响 [J]. 东北林业大学学报, 2011, 39(5): 24–26. doi: 10.3969/j.issn.1000-5382.2011.05.008.
[42] MU J. The effects of nutrient on salt tolerance and growth of sweet sorghum in salinity region [D]. Beijing: University of Chinese Academy of Sciences, 2013.
穆静. 养分对盐碱区甜高粱耐盐性和生长的影响 [D].北京: 中国科学院大学, 2013.
Difference of Leaf Functional Traits of Coastal Psammophytes in Hainan Island and Its Relationship with Soil Chemical Properties
WANG Xu1, LIU Jingkun2, LUO Shuixing2, ZHAO Kunkun2, LI Xinjian3*
(1. Research Institute of Tropical Forestry, Chinese Academy of Forestry,Guangzhou 510520, China; 2. Experimental Station of Research Institute of Tropical Forestry, Chinese Academy of Forestry,Ledong 572542, Hainan, China; 3. Central South Inventory and Planning Institute of State Forestry and Grassland Administration,Changsha 410014, China)
Leaf functional traits are closely related to plant growth strategies and resource utilization ability. In order to explore the adaptation strategies of psammophytes to environment in Hainan Island, the leaf functional traits of,,,,andand their relationships with soil chemical properties were analyzed. The results showed that the leaf functional traits of tropical psammophytes were different among species, which were affected not only by species but also by soil factors. The specific leaf area of species were in the order of herb>shrub>vine. The N/P ratio of leaves ranged from 7.78 to 10.85, suggesting that the growth of tropical psammophytes was restricted by soil N. The variation coefficient of Na content in leaves was the highest, ranging from 18.46% to 76.36%, indicating that species had a great difference in the absorption of Na+, which would affect its natural distribution in coastal sandy land. The specific leaf area, leaf dry matter content had negative correlations with K and Na contents in leaves, as well as N/P ratio of leaves with Na content in soil, Na content in leaves with soil organic carbon, total P and total K, implying that sandy plants adapted to salt stress through specific leaf area change. Therefore, applying soil fertilizer would be an important measure for rapid restoration of coastal sandy vegetation, and the Na+content in leaves could be used as the main index for salt-tolerance screening of sandy plants in tropical coast.
Psammophyte; Functional trait; Soil chemical property; Adaptation
10.11926/jtsb.4519
2021-09-03
2021-12-10
中国林业科学研究院基本科研业务专项项目(CAFYBB2018MB015)资助
This work was supported by the Special Project for Fundamental Research of Chinese Academy of Forestry (Grant No. CAFYBB2018MB015).
王旭(1977生),男,博士,副研究员,研究方向为森林生态学和恢复生态学。E-mail: cafwangxu111@caf.ac.cn
. E-mail: 282869481@qq.com
